
Download
ORIGINAL ARTICLE
Erythropoietin inhibits neutrophil extracellular traps formation to ameliorate lung injury in a pneumonia model
Sheng Yea#, Wei Lib#, Jinghui Yangc, Xiang Xuec, Jiao Chenc, Wei Zhaoc, Lei Jiangd*, Ling Jiac*
aDepartment of Respiratory and Critical Care Medicine, Nanjing BenQ Medical Center, the Affiliated BenQ Hospital of Nanjing Medical University, Nanjing, Jiangsu Province, China
bDepartment of Critical Care Medicine, Affiliated Hospital of Integrated Traditional Chinese and Western Medicine, Nanjing University of Chinese Medicine, Nanjing, Jiangsu Province, China
cDepartment of Intensive Care Unit, SIR RUN RUN Hospital of Nanjing Medical University, Nanjing, Jiangsu Province, China
dDepartment of Emergency, the First Affiliated Hospital of Naval Medical University, Shanghai, China
#These authors contributed equally to the work and should be considered first co-authors.
*Corresponding author: Lei Jiang, Department of Emergency, the First Affiliated Hospital of Naval Medical University, No. 168, Changhai Road, Shanghai 200433, China. Email address: [email protected]
Ling Jia, Department of Intensive Care Unit, SIR RUN RUN Hospital of Nanjing Medical University, No. 109 Longmian Avenue, Nanjing, Jiangsu Province 211100, China. Email address: [email protected]
Abstract
Background: Severe pneumonia is a kind of disease that develops from lung inflammation, and new drugs are still required to treat the same. Erythropoietin (EPO) is widely used to treat anemia in patients. However, there are fewer studies on the role of EPO in neutrophil extracellular trappings (NETs) and pneumonia, and the mechanism is unclear.
Objective: To investigate the possible effects of EPO on the formation of NETs and progression of pneumonia.
Methods: Mice pneumonia model was induced by tracheal lipopolysaccharide (LPS) administration. Hematoxylin and eosin (H&E) staining and automatic blood cell analysis were performed in this model to confirm the effects of EPO on lung injury. Flow cytometry, enzyme-linked immunosorbent serological assay, and immunostaining assay were conducted to detect the effects of EPO on the inflammation as well as formation of NETs in mice. Immunoblot was further conducted to confirm the mechanism.
Results: EPO alleviated LPS-induced lung injury. EPO reduced the release of inflammatory factors induced by LPS. In addition, EPO inhibited the formation of NETs. Mechanically, EPO inhibited tumor necrosis factor (TNF) receptor associated factor 2 (TRAF2)/nuclear factor kappa-B (NF-κB) activity in mouse models, and therefore suppressed the progression of pneumonia.
Conclusion: EPO inhibited formation of NETs to ameliorate lung injury in a pneumonia model, and could serve as a drug of pneumonia.
Key words: erythropoietin (epo), lung injury, neutrophil extracellular trapping (nets), pneumonia, TRAF2/NF-κB
Received 24 August 2023; Accepted 11 September 2023; Available online: 1 November 2023
Copyright: Ye S, et al
License: This open access article is licensed under Creative Commons Attribution 4.0 International (CC BY 4.0). http://creativecommons.org/licenses/by/4.0/
Introduction
Severe pneumonia develops from inflammation in the lungs, causing organ dysfunction and even life-threatening diseases. It has a case fatality rate of up to 30–50%, which causes serious complications and high economic burden.1,2 Pneumonia is often accompanied by lung damage that progresses to pulmonary edema and neutrophil accumulation.3,4 Neutrophils are the first line of defense against inflammation.3 They have three main functions: phagocytosis, degranulation, and the release of neutrophil extracellular trappings (NETs).5 NETs are reticular extracellular structures composed of DNA, histones, and myeloperoxide (MPO) neutrophil elastocytic enzymes.6 NETs are like a double-edged sword, as they trap and kill pathogens during local infections to the benefit of host cells.6 In systemic infection, acute lung injury is caused by its toxicity to lung epithelial cells, which increases case fatality rate through sepsis.7,8
Erythropoietin (EPO) is widely used to treat anemia in patients.9 In the past two decades, EPO has been used to treat various diseases.10 Studies have shown that EPO alleviates the cardiac dysfunction of mice with experimental sepsis and improves the survival rate of mice with sepsis by activating β-coreceptor.11 EPO can regulate the function of immune cells, including T cells, B cells, macrophages, etc.11 Studies have shown that EPO can promote the proliferation and differentiation of T cells and enhance their ability to kill tumor cells and pathogens.11 Meanwhile, EPO can also inhibit the activity of B cells and reduce the occurrence of autoimmune reaction.11 In addition, EPO can also promote the phagocytosis of macrophages.12 The anti-inflammatory effects of EPO are multifactorial, which are achieved by inhibiting the production of inflammatory mediators, regulating the functioning of immune cells, protecting the nervous system, and inhibiting oxidative stress.12 These effects not only have important significance for treating inflammatory diseases but also provide new ideas and directions for the application of EPO to other fields. However, few studies are found on the role of EPO in NETs and pneumonia, and the mechanism is unclear.
The aim of this study was to elucidate the effects of EPO on NET and pneumonia. We found that EPO alleviates lipopolysaccharide (LPS)-induced pneumonia by regulating tumor necrosis factor (TNF) receptor associated factor 2–nuclear factor kappa-B (TRAF2/NF-κB) activity and inhibiting the formation of neutrophil extracellular trapping NETs. Therefore, EPO can be used as a potential anti-pneumonia agent.
Materials and methods
Animal treatment
Male C57BL/6 mice (aged about 8 weeks) were accessed from the SLAC (Shanghai, China). The mice were kept in sterile atmosphere. All experimental procedures in this study were conducted under ethical approval obtained from the Institutional Animal Care and Use Committee of Nanjing BenQ Medical Center, The Affiliated BenQ Hospital of Nanjing Medical University (Approval No. SYXK-2020-0022). The mice were sacrificed by cervical dislocation, and the lack of heartbeat was confirmed to validate death. The mice were monitored once a day before the experiment began and twice a day until the experiment ended. The mice were anesthetized by injection of 4% chloral hydrate and instilled with 3-mg/kg LPS (Escherichia coli 0111:B4; Sigma, St. Louis, MO, US) dissolved in 50-μL phosphate-buffered saline (PBS) or only sterile PBS (control group). EPO (1248200; Sigma) was administered by oral gavage 1 h after LPS treatment as follows: bronchoalveolar lavage fluid (BALF) and lung tissues were collected to monitor pathological changes and production of cytokines. The total number of cells and neutrophils were observed in BALF.
Histological analysis
Lung tissues were fixed with 5% paraformaldehyde (PFA), embedded with paraffin, and cut into slices. The sections were counterstained with hematoxylin and eosin (H&E). The dried slices were removed from warm tank and immediately de-waxed into xylene for 5–10 min depending on whether the wax was not dissolved completely. Low temperature extends the time whereas high temperature appropriates to shorten the time or accelerate de-waxing in temperature box. The sample was then transferred into gradient alcohol. It was then moved into hematoxylin and impregnated for 5 min; then transferred into eosin solution and impregnated for 2–5 min, and finally transferred into xylene I transparent for 3–5 min and sealed.
Lung injury score was marked considering hemorrhage, alveolar wall thickness, inflammatory infiltration, alveolar congestion, and hyaline membrane formation. The score of 0 means no damage; 1 represents mild damage; 2 represents moderate damage; 3 for severe damage, and 4 for very severe histological alterations.
Lung wet-to-dry (W/D) weight ratio and mean platelet volume measurement
The lung samples were dissected after the mice were sacrificed, and weighed immediately. Then, lung samples were dried until the weight remained unchanged. The W/D weight ratio and mean platelet volume were calculated.
Enzyme-linked-immunosorbent serologic assay (ELISA)
The levels of tumor necrosis factor-α (TNF-α) (PT512), interleukin 6 (IL-6) (PI326), IL-1β (PI301), and IL-17A (PI545), bought from Beyotime (Beijing, China) in BALF were assessed using ELISA kit according to manufacturer’s instructions. Briefly, samples were aspirated into wells. Biotin-conjugated primary antibodies were added into wells prior to adding avidin-conjugated horseradish peroxidase (HRP). Then enzyme substrate was added for color development. The intensity of each well was measured with a microplate reader.
Immunofluorescence
Lung tissues were fixed with 5% PFA and embedded with optimal cutting temperature (OCT) compound. Afterwards, sections were sliced and incubated with MPO (rabbit, ab208670, 1:200; Abcam, Cambridge, UK) and histone (mice, ab52484, 1:200; Abcam) primary antibody at 4°C overnight. After rinsing in PBS, slices were incubated with respective secondary antibody and counterstained with 4',6-diamidino-2-phenylindole (DAPI). Cover slips were examined using a microscope and analyzed by the ImageJ 9.0 software.
Western blot analysis
Tissue lysates were collected after addition of a buffer containing 1% Triton X-100, 150-mM NaCl, and 50-mM Tris (pH 7.5). After collecting supernatant, protein concentration was obtained by a bicinchoninic acid (BCA) protein assay kit (Beyotime). Then proteins were separated with 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE), transferred onto polyvinylidene difluoride membranes (MilliporeSigma, MA, US), which were blocked at room temperature for 2 h in Tris-buffered saline containing 0.2% Tween 20 and 5% nonfat milk. Then proteins were complied by primary antibodies targeting TRAF2 (ab126758, 1:1000; Abcam), p65 (ab32536, 1:1000; Abcam), p-p65 (ab76302, 1:1000, Abcam), and β-actin (ab8226, 1:3000, Abcam). Membranes were maintained in HRP-conjugated secondary antibodies for 2 h after rinsing in tris buffered saline with tween (TBST) for 15 min. The signals were discovered with enhanced chemiluminescence (ECL) detection kit.
Statistics
Data were analyzed using the GraphPad 8.0 software (GraphPad Software Inc., La Jolla, CA, US). Error bars represented mean ± SD. The unpaired Student’s t-test was used to determine statistical significance between two groups. One-way ANOVA, followed by Tukey’s post hoc test, was used for multiple comparisons. P < 0.05 was considered statistically significant.
Results
EPO alleviated LPS-induced lung injury
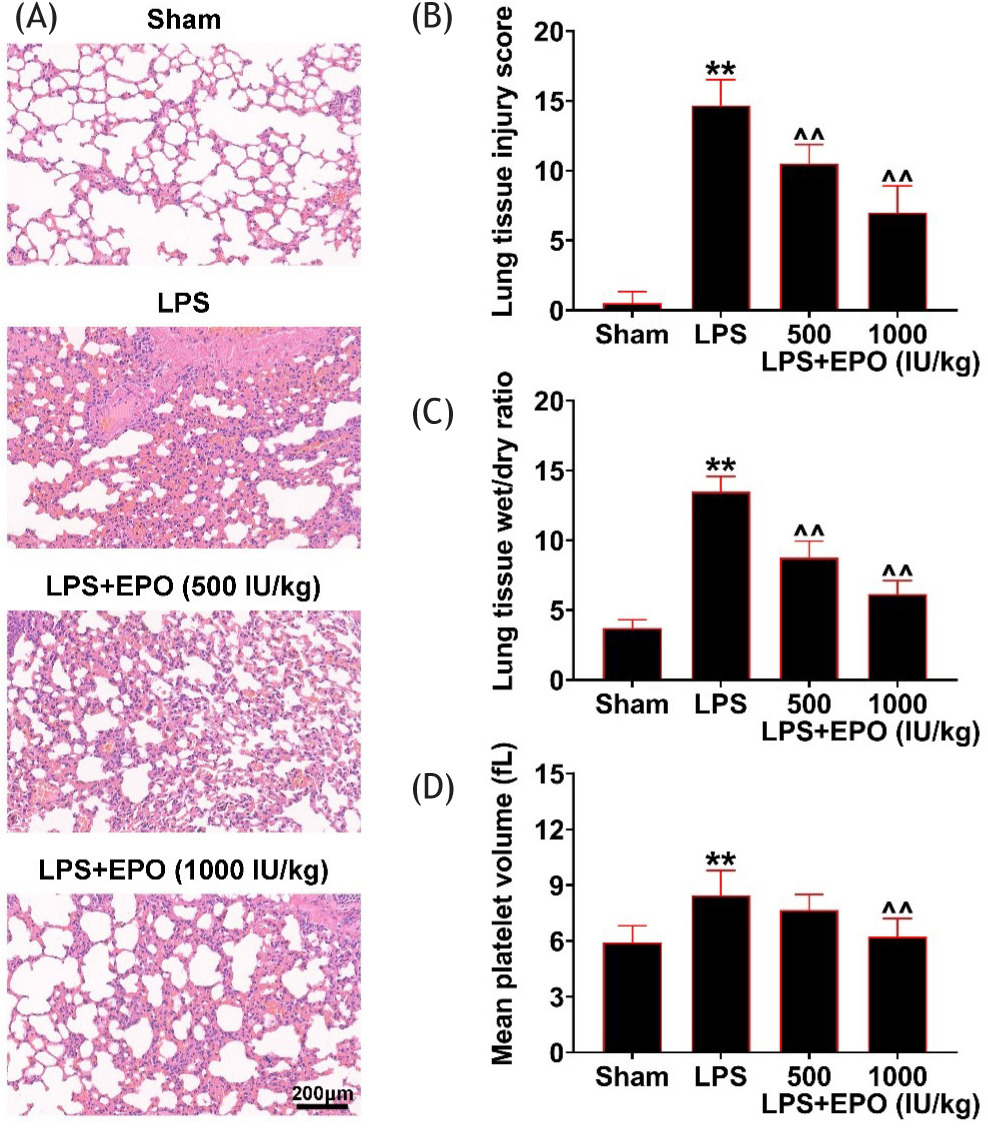
After induction of acute lung injury, the histological examination of lung tissues was conducted. The LPS-induced mice displayed tissue injury characterized by irregular alveolar integrity and alveolar size, partial collapse in alveoli, enlarged spaces between the alveoli, and inflammatory cell infiltration (Figure 1A). These phenotypes were recovered after administration of EPO as well as the relieved injury score (Figure 1B). Moreover, the W/D ratio and mean platelet volume, which reflect the degree of lung injury, enhanced by LPS, was improved by EPO treatment (Figure 1C). These results reflected the alleviated effect of EPO in lung injury.
Figure 1 EPO alleviated LPS-induced lung injury. (a) The histological analysis of lung tissues in control, LPS, EPO (500 IU/kg)-treated LPS, and EPO (1000 IU/kg)-treated LPS mice. The H&E staining was performed. Scale bar: 200 um. (b) Lung tissue injury score in control, LPS, EPO (500 IU/kg)-treated LPS, and EPO (1000 IU/kg)-treated LPS mice. (c) Lung tissue wet/dry ratio and (d) mean platelet volume in mice of control, LPS, EPO (500 IU/kg)-treated LPS, and EPO (1000 IU/kg)-treated LPS groups. **P < 0.01 vs. control; ^^P < 0.01 vs. LPS
EPO reduced the release of inflammatory factors induced by LPS
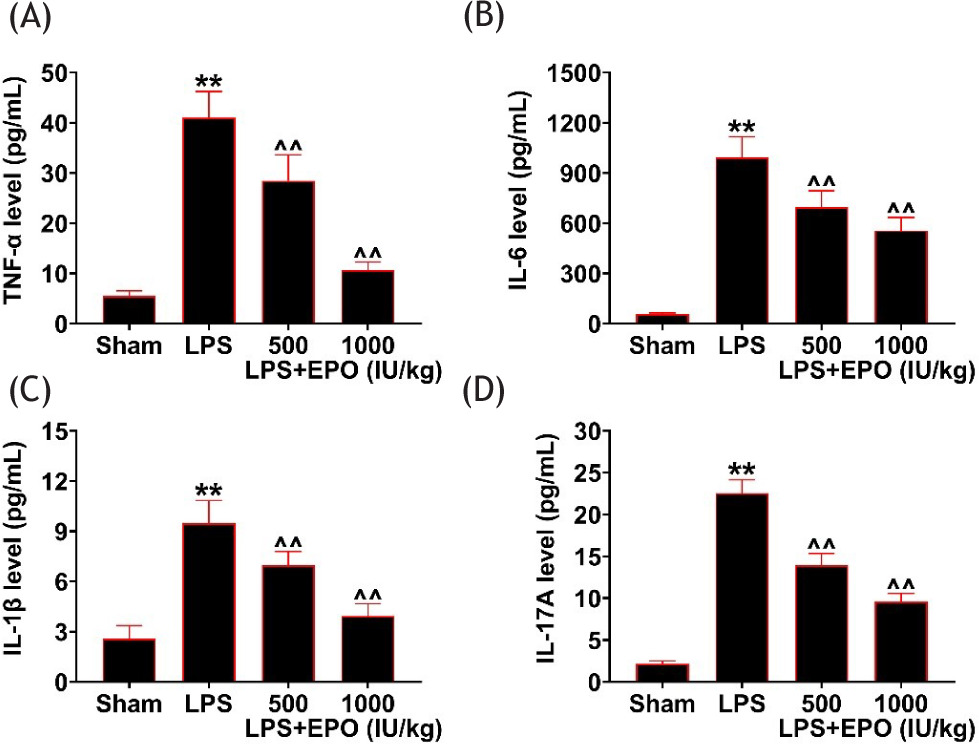
We then discovered the effect of EPO on the inflammation of lung tissues in LPS model. The inflammatory cytokines were also monitored in LPS model. The levels of inflammatory factors, such as TNF-α, IL-6, IL-1β, and IL-17A, were enhanced in LPS mice (Figures 2A–2D). However, EPO significantly rescued the enhanced levels of TNF-α, IL-6, IL-1β, and IL-17A in the BALF of LPS mice (Figures 2A–2D). Therefore, EPO reduced the release of inflammatory factors induced by LPS.
Figure 2 EPO reduced the release of inflammatory factors induced by LPS. The levels of (a) TNF-α, (b) IL-6, (c) IL-1β, and (d) IL-17A in control, LPS, EPO (500 IU/kg)-treated LPS, and EPO (1000 IU/kg)-treated LPS mice were observed through ELISA. ##P < 0.01 vs. Sham; ^^P < 0.01 vs. LPS
EPO inhibited the formation of NETs
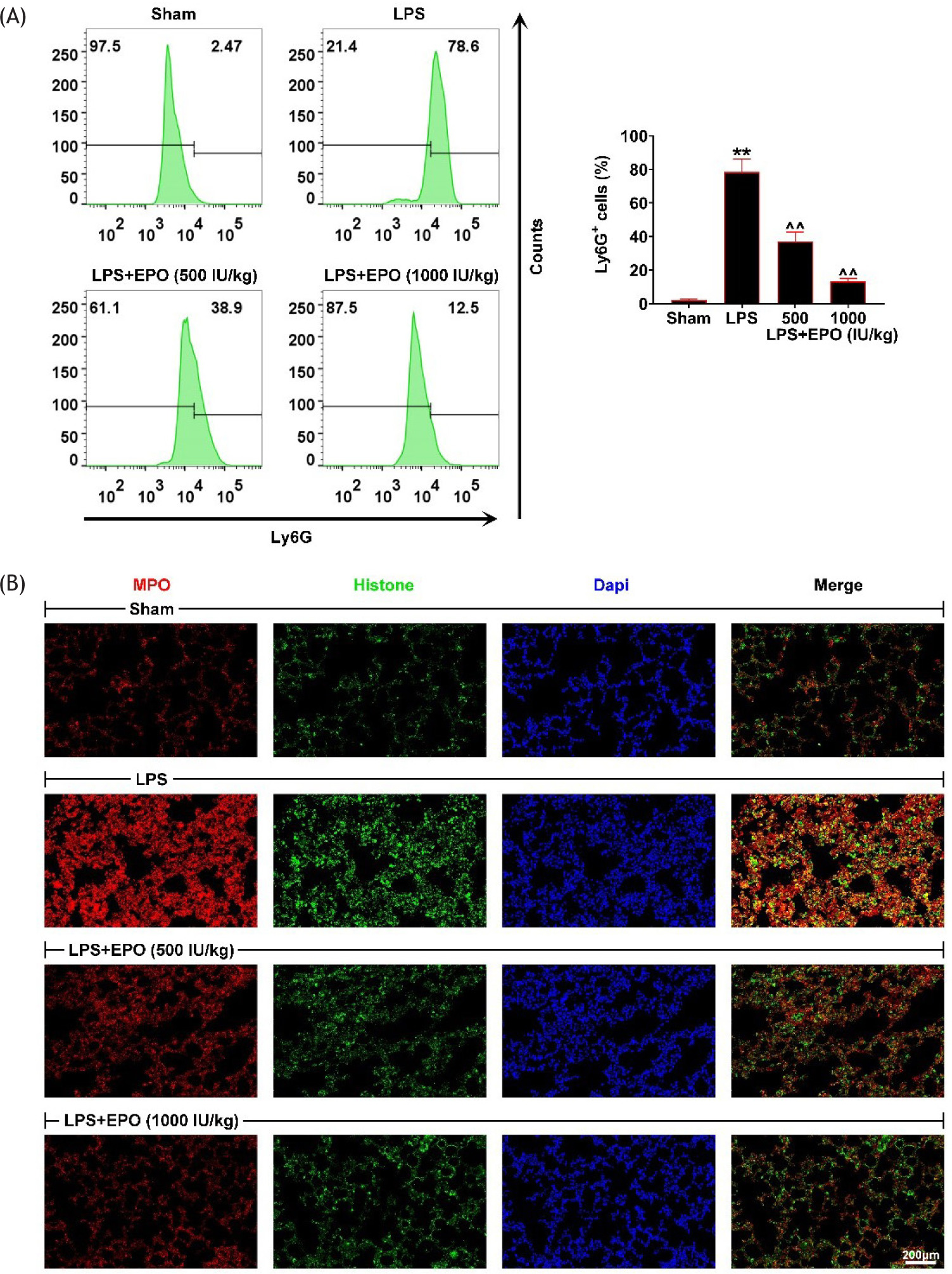
Subsequently, we investigated the effects of EPO on the formation of NETs of lung tissues in LPS mice. Through flow cytometry (FCM) assays, we discovered that LPS induction increased the number of neutrophils (Ly6G+ cells) in lung tissues (Figure 3A). However, EPO treatment reversed the increased number of neutrophils caused by LPS induction (Figure 3A). In addition, we observed the formation of NET through immunostaining assays. Histone and MPO are the main components of NETs, and their co-localization promotes the formation of NETs. We found that LPS induction promoted the formation of NETs, with increased -co-localization degree of histone and MPO (Figure 3B). However, EPO rescued the promotion of co-localization degree of histone and MPO caused by LPS treatment in lung tissues (Figure 3B). Therefore, EPO inhibited the formation of NETs in lung tissues of LPS-induced lung injury mice.
Figure 3 EPO inhibited the formation of NETs. (a) The number of neutrophils in control, LPS, EPO (500 IU/kg)-treated LPS, and EPO (1000 IU/kg)-treated LPS mice was measured by flow cytometry. Ly6G-positive cells were measured to reflect the formation of NETs. (b) Immunostaining assays showed the expression of MPO (red) and histone (green) in lung tissues from control, LPS, EPO (500 IU/kg)-treated LPS, and EPO (1000 IU/kg)-treated LPS mice. Scale bar: 200 um. **P < 0.01 vs. Sham; ^^P < 0.01 vs. LPS
EPO inhibited TRAF2/NF-κB activity in LPS-induced lung injury models
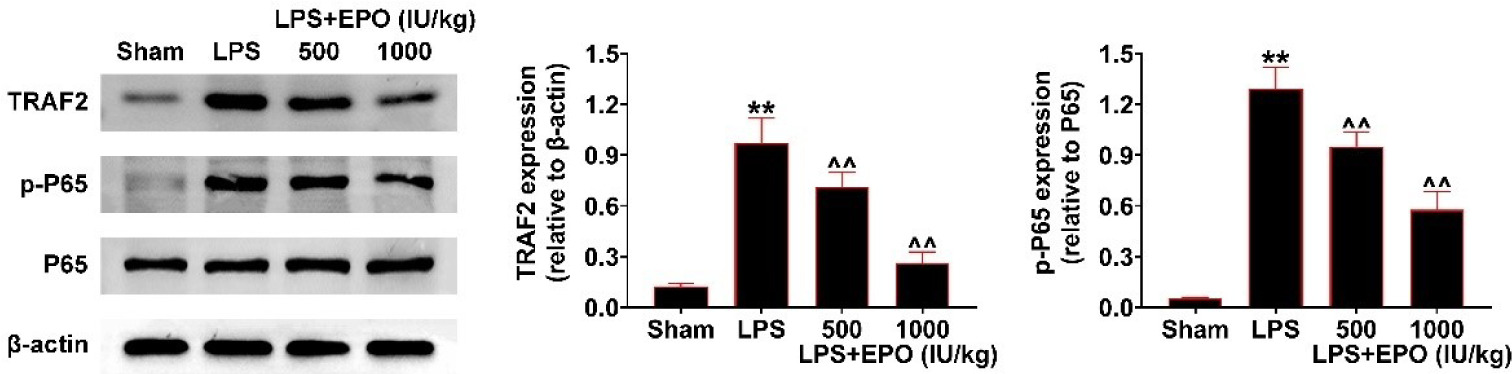
To unveil the mechanism underlying EPO suppressing lung injury, we observed the role of EPO in TRAF2/NF-κB pathway. We observed the upregulation of TRAF2 expression and p65 phosphorylation caused by LPS stimulation (Figure 4). EPO treatment further suppressed the expression of TRAF2 and phosphorylation levels of p65 in LPS-induced lung injury mice (Figure 4). Therefore, the data confirmed that EPO inhibited the TRAF2/NF-κB activity in LPS-induced lung injury models.
Figure 4 EPO inhibited TRAF2/NF-κB activity in mouse models. Immunoblot showed the expression of TRAF2, p65, and the phosphorylation levels of p65 in lung tissues from control, LPS, EPO (500 IU/kg)-treated LPS, and EPO (1000 IU/kg)-treated LPS mice. The relative expression was quantified. **P < 0.01 vs. Sham; ^^P < 0.01 vs. LPS.
Discussion
Severe pneumonia is the primary cause of death in children.6 Lung injury refers to the injury caused by multiple factors. Lung injury can occur at all ages and may be associated with various factors, such as accidents, inhalation of stomach contents, infection, lung surgery, oxygen poisoning, etc. Neurodegenerative diseases and cardiovascular and cerebrovascular diseases also cause lung damage.2 With the understanding of coagulation disorders in severe infections, it was found that immune thrombosis formed by the interaction between external neutrophil trapping network and coagulation factors played an important role in preventing the spread of pathogenic microorganisms during early stages as well as in the identification and removal of pathogenic microorganisms.5,13,14 Moreover, some other extrapulmonary factors affecting the clinical course of pneumonia were revealed.3
In addition, exercise training is one of the rehabilitation strategies that have anti-inflammatory effects and maybe willcould serve as a good strategy for to decrease lung injury. While recovering from pneumonia, patients should actively participate in appropriate physical activities according to theirdepending on physical strength. However, excessive activation of the coagulation system causes microcirculation disorders and tissue ischemia and hypoxia.5 Dysfunctional neutrophils are recruited into lung tissues, often promoting abnormal inflammatory responses.5 If infection persists, it could further promote neutrophil migration and aggregation.5,6 Intervention of neutrophil aggregation is an effective strategy to improve severe pneumonia. We found that EPO relieves severe pneumonia and NETs through this process.
Neutrophil extracellular trappings are a network of DNA fibers, histones, and antibacterial proteins secreted by activated neutrophils responsible for trapping and killing extracellular pathogens and having a protective role in antibacterial defense.5,7 The process by which neutrophils secrete NETs, known as NETosis, is a mode of inflammatory cell death of neutrophils.5,7 This cell death is distinct from apoptosis and necrosis, and usually requires neutrophils activation and nicotinamide adenine dinucleotide phosphate (NADPH) oxidase production of reactive oxygen species (ROS).6,15 Through FCM, ELISA, and immunostaining assay, we revealed that EPO inhibits the formation of NETs. We anticipated that EPO suppressed the progression of pneumonia by inhibiting NETs.
Erythropoietin is an endogenous glycoprotein hormone that stimulates red blood cell production.9,12 Hypoxia stimulates the production of EPO, and recombinant human EPO has been used for long in the clinical treatment of a variety of diseases.9,10,12 Previous studies have confirmed that EPO inhibits the production of inflammatory mediators, reduces the degree and duration of inflammatory response, and thus protects tissues from inflammation.9,12,16 However, our data confirmed that EPO affected inflammation in pneumonia through a different mechanism. The inhibition of oligomerization domain-like receptor family pyrin domain-containing 3 (NLRP3) inflammasome by EPO via the erythropoietin receptor protein–Janus kinase 2–signal transducer and activator of transcription 3 (EPOR/JAK2/STAT3) pathway alleviates acute lung injury in mice.17 EPO promotes mitochondrial respiration in developing postnatal hippocampus by increasing mitochondrial content and enhancing cellular respiratory potential, thereby enhancing cognitive function in mice.18 EPO improves lupus nephritis in Murphy Roths large–lymphoproliferative (MRL/LPR) mice by increasing spleen Th2 and Treg lymphocytes, decreasing Th1 and Th17 lymphocytes, inhibiting inflammation of the spleen and kidney.19 EPO reduces apoptosis and regulates macrophage polarization through EPOR/JAK2/STAT3 axis to prevent diffused alveolar hemorrhage in mice.20 EPO also improves ischemia/reperfusion-induced acute kidney injury by reducing the cyclooxygenase-2 (COX-2) and NF-κB pathways and inhibiting inflammasome in mice.20 Next, we elucidated whether EPO affected NETs and pneumonia through these mechanisms.
NF-κB is a binding protein that regulates gene expression.21 This pathway plays an important role in pulmonary inflammatory diseases by regulating the gene expression of many important cytokines and adhesion molecules in immune cells and airway epithelial cells, and by participating in lung inflammation and immune response.21 Studies have shown that EPO reduces the expression of pro-inflammatory factors and reduces inflammatory response in traumatic ischemia of the brain and spinal cord and in cerebral multiple sclerosis models.22 The nuclear NF-κB family is a major regulator of inflammatory genes. After spinal cord injury, NF-κB is obviously up-regulated, and EPO exerts its anti-inflammatory effect by regulating NF-κB.22 Interestingly, we also revealed that EPO could mediate NF-κB pathway, and therefore affected the progression of pneumonia, although the precise mechanism needs further study.
Conclusion
In summary, we found that EPO inhibits NETs formation to relieve lung injury in a pneumonia model. We believed that EPO could act as a promising drug to treat pneumonia.
Funding
This work was supported by Research Project on Elderly Health of Jiangsu Provincial Health Commission (Grant No. LKM2022009) and Jiangsu Provincial Health Commission Research Project (Grant No. M2022033).
Competing interests
The authors stated that there was no conflict of interest to declare.
Data availability
The authors declare that all data supporting the findings of this study are available within the paper and any raw data can be obtained from the corresponding author upon request.
Author Contributions
Sheng Ye, Lei Jiang, and Ling Jia designed and carried out the study. Sheng Ye, Wei Li, Jinghui Yang, Xiang Xue, Jiao Chen, and Wei Zhao supervised data collection, analysis, and interpretation. Sheng Ye, Lei Jiang, and Ling Jia prepared the manuscript for publication and reviewed its draft. All authors had read and approved the final manuscript.
REFERENCES
1. Kroner PT, Picco MF, Cangemi JR, Hayney MS, Farraye FA, Caldera F. The burden of vaccine-preventable diseases in patients with inflammatory bowel disease. J Clin Gastroenterol. 2022;56. 10.1097/MCG.0000000000001635
2. Rasouli S, Heshmatnia J, Mosaffa N, Marjani M, Mortaz E. Dysregulation of immunity in pulmonary fibrosis is associated with increased myeloid-specific triggering receptor-1 and transforming growth factor-beta 1 expression. Iran J Allergy Asthma Immunol. 2023;22(1):12–24. 10.18502/ijaai.v22i1.12002
3. Miethe S, Potaczek DP, Bazan-Socha S, Bachl M, Schaefer L, Wygrecka M, et al. The emerging role of extracellular vesicles as communicators between adipose tissue and pathologic lungs with a special focus on asthma. Am J Physiol Cell Physiol. 2023;324:C1119–25. 10.1152/ajpcell.00057.2023
4. Luan Y, Liu H, Luan Y, Yang Y, Yang J, Ren KD. New insight in HDACs: Potential therapeutic targets for the treatment of atherosclerosis. Front Pharmacol. 2022;13:863677. 10.3389/fphar.2022.863677
5. Sanders NL, Martin IMC, Sharma A, Jones MR, Quinton LJ, Bosmann M, et al. Neutrophil extracellular traps as an exacerbating factor in bacterial pneumonia. Inf Immun. 2022;90(3):e0049121. 10.1128/iai.00491-21
6. Pulavendran S, Rudd JM, Maram P, Thomas PG, Akhilesh R, Malayer JR, et al. Combination therapy targeting platelet activation and virus replication protects mice against lethal influenza pneumonia. Am J Resp Cell Mol Biol. 2019;61(6): 689–701. 10.1165/rcmb.2018-0196OC
7. Sung PS, Peng YC, Yang SP, Chiu CH, Hsieh SL. CLEC5A is critical in pseudomonas aeruginosa-induced NET formation and acute lung injury. JCI Insight. 2022;7(18). 10.1172/jci.insight.156613
8. Yao ME, Huang Y, Dong QQ, Lu Y, Chen W. The renshen chishao decoction could ameliorate the acute lung injury but could not reduce the neutrophil extracellular traps formation. Evidence Complement Alternat Med (eCAM). 2022;2022:7784148. 10.1155/2022/7784148
9. Duygu G, Yalcin-Ulker GM, Gunbatan M, Soluk-Tekkesin M, Ozcakir-Tomruk C. Evaluation of preventive role of systemically applied erythropoietin after tooth extraction in a bisphosphonate-induced MRONJ model. Medicina. 2023;59(6). 10.3390/medicina59061059
10. Chen H, Yu J, Wei Q, Zhang Y, Ouyang X, Wang S. Intravenous iron and erythropoietin therapy for postoperative anemia among orthopedic surgery patients. J Orthop Surg Res. 2023;18(1):510. 10.1186/s13018-023-03926-y
11. Skrifvars MB, Luethi N, Bailey M, French C, Nichol A, Trapani T, et al. The effect of recombinant erythropoietin on long-term outcome after moderate-to-severe traumatic brain injury. Intensive Care Med. 2023;49(7):831–9. 10.1007/s00134-023-07141-5
12. Horwitz JK, Bin S, Fairchild RL, Keslar KS, Yi Z, Zhang W, et al. Linking erythropoietin to treg-dependent allograft survival through myeloid cells. JCI Insight. 2022;7(10). 10.1172/jci.insight.158856
13. Li J, Yan W, Ren F, Sang H. Tectorigenin inhibits inflammation in keratinocytes by inhibition of NLRP3 inflammasome regulated by the TLR4/NF-kappaB pathway. Allergol Immunopathol (Madr). 2023;51(2):82–9. 10.15586/aei.v51i2.780
14. Liu L, Huan L, Zhang Y, Wei W, Chen Z, Xu D, et al. Ubiquitin-specific protease 8 inhibits lipopolysaccharide-triggered pyroptosis of human bronchial epithelial cells by regulating PI3K/AKT and NF-kappaB pathways. Allergol Immunopathol (Madr). 2022;50(2):96–103. 10.15586/aei.v50i2.568
15. Regua A, Papp C, Grageda A, Porter B, Caza T, Bichindaritz I, et al. ABI1-based expression signature predicts breast cancer metastasis and survival. Mol Oncol. 2021;16:2632-2657 10.1002/1878-0261.13175
16. Li C, Fan R, Li DB, Zhou W, Huang YE, Wang B. Intervention effects of erythropoietin derived peptide (HBSP) on renal injury induced by acute skeletal muscle strain in rats. Zhongguo ying yong sheng li xue za zhi. (Chin J Appl Physiol). 2022;38(6):617–21.
17. Cao F, Tian X, Li Z, Lv Y, Han J, Zhuang R, et al. Suppression of NLRP3 inflammasome by erythropoietin via the EPOR/JAK2/STAT3 pathway contributes to attenuation of acute lung injury in mice. Front Pharmacol. 2020;11:306. 10.3389/fphar.2020.00306
18. Jacobs RA, Aboouf MA, Koester-Hegmann C, Muttathukunnel P, Laouafa S, Arias-Reyes C, et al. Erythropoietin promotes hippocampal mitochondrial function and enhances cognition in mice. Commun Biol. 2021;4(1):938. 10.1038/s42003-021-02465-8
19. Chen H, Luo B, Yang X, Xiong J, Liu Z, Jiang M, et al. Therapeutic effects of nonerythropoietic erythropoietin analog ARA290 in experimental autoimmune encephalomyelitis rat. J Neuroimmunol. 2014;268(1–2):64–70. 10.1016/j.jneuroim.2014.01.006
20. Wang S, Zhang C, Li J, Niyazi S, Zheng L, Xu M, et al. Erythropoietin protects against rhabdomyolysis-induced acute kidney injury by modulating macrophage polarization. Cell Death Dis. 2017;8(4):e2725. 10.1038/cddis.2017.104
21. Wu X, Lv Y, Li Z, Yang Z. Serelaxin inhibits LPS-induced inflammatory response in cardiac fibroblasts by activating PPAR-gamma and suppressing NF-kappaB signaling pathway. J Cardiovas Pharmacol. 2023;82: 201-211.
22. Simon F, Floros N, Ibing W, Schelzig H, Knapsis A. Neurotherapeutic potential of erythropoietin after ischemic injury of the central nervous system. Neural Regen Res. 2019;14(8):1309–12. 10.4103/1673-5374.253507