
Download
ORIGINAL ARTICLE
Kaempferitrin alleviates LPS-induced septic acute lung injury in mice through downregulating NF-κB pathway
Xiaoli Zhua*, Yongyue Pana, Xin Xub, Jing Xua
aCritical Care Medicine, Zhejiang Youth Hospital, Hangzhou, Zhejiang Province, China
bCritical Care Medicine, Zhejiang Chinese Medicine University, Hangzhou, Zhejiang Province, China
*Corresponding author: Xiaoli Zhu, Critical Care Medicine, Zhejiang Youth Hospital, No. 54, Qingchun East Road, Kaixuan Street, Shangcheng District, Hangzhou City, Zhejiang Province 310000, China. Email address: [email protected]
Abstract
Background: Acute lung injury (ALI) causes severe and uncontrolled pulmonary inflammation and has high morbidity in dying patients.
Objective: This study aimed to evaluate the potential function of Kaempferitrin (Kae) and uncover its mechanisms in ALI.
Material and Methods: We evaluated the role of Kae in ALI through the lipopolysaccharide (LPS)-induced histopathological changes, lung wet/dry (W/D) ratio, total bronchoalveolar lavage fluid (BALF) cells count, pulmonary inflammation, and the levels of interleukin (IL)-6, tumor necrosis factor-α (TNF-α), and IL-1β. The effect of Kae on NF-κB signaling pathway was discovered through the protein expression levels of transcription factors p65, p-p65, IκBα, and p-IκBα by Western blot analysis.
Results: The results showed that Kae could improve lung injury by reducing apoptosis, histopathological changes, and lung W/D ratio; more importantly, Kae enhanced the survival of ALI mice. Moreover, Kae relieved inflammation, as it reduced total BALF cells count, and deceased the levels of TNF-α, IL-6, and IL-1β in serum. In addition, Western blot analysis data suggested that Kae could decrease the protein expression levels of transcription factors p65, p-p65, IκB-α, and p-IκB-α, which were promoted by LPS.
Conclusion: The results of this study suggested that Kae could relieve LPS-induced ALI in mice and reduce inflammation and apoptosis through NF-κB pathway.
Key words: acute lung injury, kaempferitrin, lipopolysaccharide, NF-κB pathway, sepsis
Received 9 February 2023; Accepted 27 February 2023; Available online 1 November 2023
Copyright: Zhu X, et al.
License: This open access article is licensed under Creative Commons Attribution 4.0 International (CC BY 4.0). http://creativecommons.org/licenses/by/4.0/
Introduction
Sepsis is a latently life-threatening systemic inflammatory syndrome characterized by inflammatory disorders, coagulation dysfunction, and multiple organ failure.1 World Health Organization (WHO) has emphasized the importance of sepsis because of its enormous impact on global mortality.2 In the context of antibiotic use, the mortality rate of severe sepsis is as high as 20–30%.3 There is an urgent need to identify new targets to improve the survival outcomes of sepsis.4 Moreover, acute lung injury (ALI) causes the most usual organ damage in sepsis.5 ALI also causes severe inflammation.6 Recent epidemiologic studies have suggested that ALI is an important public health issue.7 Despite the development of clinical treatment for better outcome, the mortality rate because of ALI remains high.8 Thus, it is essential to explore new therapeutic methods for managing ALI.
Lipopolysaccharide (LPS) is an endotoxin and a key constituent of the cell wall of Gram-negative bacteria, and ithashaving a complex function in pneumonia.9 Systemic or intranasal administration of LPS has been widely used to induce lung inflammation in animal models of ALI.2 Endotoxin can binds to toll-like receptor (TLR)-4, resulting in the production of chemokine and pro-inflammatory cascade reactions, that might beare vital for the pathogenesis of ALI.10 Previous studies suggested that sepsis was constantly adjoinedadjointed by the up-regulation of nuclear factor kappa B (NF-κB) activation.11 The inhibition of NF-κB activation could reduce inflammatory reaction, and meliorateimprove the survival of septic experimental mice.12 NF-κB transcription factor an important regulatory factor of immune response, whereas p65 is a member of NF-κB family.13 In fact, ALI induced by LPS in mice has been a well-established method for investigating inflammatory and histological changes that take place in the lungs.14
Kaempferitrin (Kae) is a potent flavonoid compound extracted from Cinnamomum osmophloeum.15 Kae has an abundant of biological activities, such as anti-inflammatory, antioxidant, anticonvulsant, antidepressant, and anti-tumor activities.16 It is reported that Kae suppresses NF-κB and protein kinase B (PKB/Akt)/mammalian rapamycin target protein (mTOR) pathway activation.15 In addition, treatment with Kae in a mouse model of collagen-induced arthritis decreases interleukin (IL)-1β, IL-6, and tumor necrosis factor-α (TNF-α) levels in serum.15 Kae restrains the exudate of white blood cells and bradykinin-induced pleurisy.17 However, the functioning and mechanism of Kae in sepsis lung injury needs further exploration.
In the present study, the findings suggested that Kae inhibited LPS-induced septic ALI in mice by suppressing NF-kB pathway.
Methods
Animals
Specific pathogen-free (SPF) C57BL/6 male mice, aged 6–8 weeks, were bought from the Laboratory Animal Center of Zhejiang Chinese Medicine University (Hangzhou, China). Body weight of the mice was 20–24 g, and lived in cages. The mice were supplemented with free water and food. All experiments involving the animals were approved by the Animal Ethical and Welfare Committee of ZCMU (Approval No. IACUC-20230130-05).
Murine model of LPS-induced ALI
The mice were randomly divided into the following four groups (every group having six animals): control group, LPS group, Kae+LPS group (Kae 10 mg/kg), and Kae+LPS group (Kae 20 mg/kg). Intraperitoneal injection of Kae was given to all animals for 2 days. After 30 min, 30-mg/kg pentobarbital sodium (Beijing Chemical Co., China) was injected to anesthetize the animals. Then, the mice were intraperitoneally injected with LPS (10 µg/mouse) (L6261; Sigma-Aldrich) to build the model of LPS-induced ALI. Mice were executed after 24 h of LPS stimulation.18)
Collection of bronchoalveolar lavage fluid
Bronchoalveolar lavage fluid (BALF) was collected through i/v cannulation, and the left lung was rinsed thrice with 2-mL phosphate-buffered saline (PBS).(19) Then BALF was centrifuged at 300×g for 10 min. We collected and resuspended the cell pellet of BALF with PBS.
Enzyme-linked immunosorbent serologic assay (ELISA)
The expression levels of TNF-α (PT518; Beyotime Institute of Biotechnology, China), IL-1β (PI305; Beyotime Institute of Biotechnology), and IL-6 (PI330; Beyotime Institute of Biotechnology) were detected by ELISA kits by following manufacturer’s instructions. Briefly, samples or standards of different concentrations were added into the corresponding wells of 96-well plates (100 μL/well), and incubated at room temperature for 120 min. After the plates were washed for five times, 100-μL antibody was added into wells of the experiment and incubated for 60 min at room temperature. Then, 100-μL horseradish peroxidase (HRP)-conjugated streptavidin was added and incubated for 20 min in dark at room temperature. Tetramethylbenzidine (TMB) developing solution, 100 μL, was added and incubated for 20 min in dark at room temperature. Finally, 50-μL/well termination solution was added and absorption was measured at 450 nm immediately.
Lung wet/dry (W/D) weight measurement
The W/D ratio was applied for assessing the gravity of pulmonary edema. Lung tissues were collected and metage was made immediately for wet weight (W) after execution of mice. After that, dry lung tissues were obtained from the wet tissues placed in an oven at 60°C for 48 h. Finally, the W/D ratio was obtained by fractioning the weight of dry tissues with the weight of wet tissues.
Hematoxylin and eosin (H&E) staining
Neutral buffered formalin 10% was applied for fixing the right lower lung of each mouse for 24 h. Then, tissues were cut into 3-μm-thick sections by rotary microtome after they were dehydrated and embedded in paraffin. H&E stain was applied for staining 3-μm-thick lung tissues to analyze pathological alterations.
TUNEL kit staining was used to detect level of lung tissue apoptosis in each group
The paraffin-fixed sections of mouse lung were dewaxed and stained with terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) reagent to detect the apoptosis of cells (green fluorescence) according to the instructions, and 4',6-diamidino-2-phenylindole (DAPI) was used for nuclear staining (blue fluorescence).
Western blot analysis
Total protein was extracted by radioimmunoprecipitation assay (RIPA) lysis and extraction buffer (89901; Thermo Scientific, Rockford, IL). Protein was quantitated by bicinchoninic acid (BCA) protein assay (23227; Thermo Scientific). Then, sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) was used to separate total proteins. Next, the proteins were transferred on polyvinylidene fluoride (PVDF) membranes. PVDF membranes with protein were blocked in 5% non-fat milk powder dissolved in tris buffered saline with tween (TBST) for 1 h at room temperature. The membranes were probed with primary antibodies at 4°C overnight, including BAX (ab32503, 1:500; Abcam, Cambridge, UK), cleaved caspase-3 (ab32042, 1:500; Abcam), p-65 (ab204604, 1:500; Abcam), p-p65 (ab31624, 1:500; Abcam), IκBα (ab32518, 1:500; Abcam), p-IκBα (ab133462, 1:500; Abcam), and β-actin (ab8226, 1:1,000; Abcam). After this, on the second day, anti-rabbit antibody (ab190492, 1:1,000; Abcam) was applied for incubating the membranes for 1 h at room temperature. The immunoreactive bands were quantitated by the Image J software.
Statistical analyses
The data were analyzed by the SAS 9.0 software. Statistical comparison of two groups was compared by unpaired two-tailed Student’s t-test. Statistical comparison of more than two groups was compared by one-way ANOVA, followed by Tukey’s post hoc test. P < 0.05 was considered as statistically significant.
Results
Kaempferitrin improved LPS-induced lung injury in mice
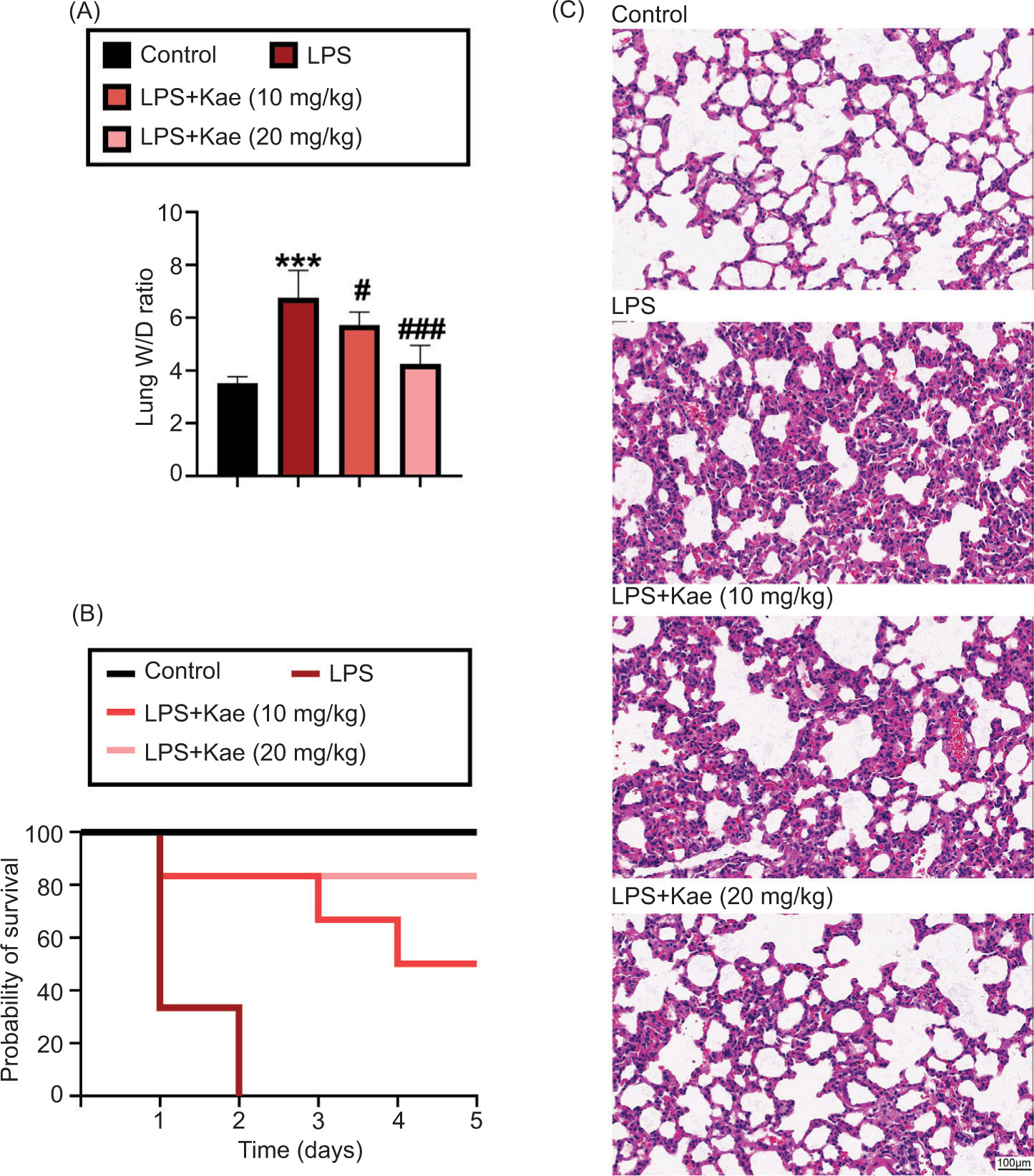
Acute lung injury model was established successfully by the lung pathological alteration assessed via H&E staining. The pathological changes were presented as destroyed pulmonary architecture, notable inflammatory cell infiltration, thickened alveolar septa, and alveolar hemorrhage in LPS-induced ALI mice. Pulmonary edema of mice was evaluated by lung W/D ratio. As shown in Figure 1A, the W/D ratio in the LPS group was significantly higher than in the control group. However, the W/D ratio was significantly lower in the Kae+LPS group (Kae 10 mg/kg) and the Kae+LPS group (Kae 20 mg/kg) than in the LPS group. As shown in Figure 1B, mice in the LPS group died within 2 days, compared with the control group. One of the mice died on the first day in the Kae+LPS group (Kae 10 mg/kg), another died on the third day, and the third one died on the fourth day. All mice in the Kae+LPS group (Kae 20 mg/kg) were alive, indicating that the survival rate of mice in these Kae+LPS groups was evidently longer than in the LPS group. Compared to the LPS group, pathological analysis indicated that Kae treatment significantly reduced lung weight and improved the condition of scatted lung tissues from acute injury in the Kae+LPS group (Kae 10 mg/kg) and the Kae+LPS group (Kae 20 mg/kg) (Figure 2C). In a word, results suggested that Kae could mitigate pathological process of ALI in vivo.
Figure 1 Successful establishment of LPS-induced ALI in vivo. (A) The lung wet/dry (W/D) weight ratio. (B) Probability of survival. (C) Histological changes in the lungs are assessed by H&E staining. *P < 0.005, **P < 0.001, ***P < 0.0001 (n = 3), #P < 0.005, ##P < 0.001, ###P < 0.0001 (n = 3). *LPS group, compared with the control group; #Kae+LPS group, compared with the LPS group.
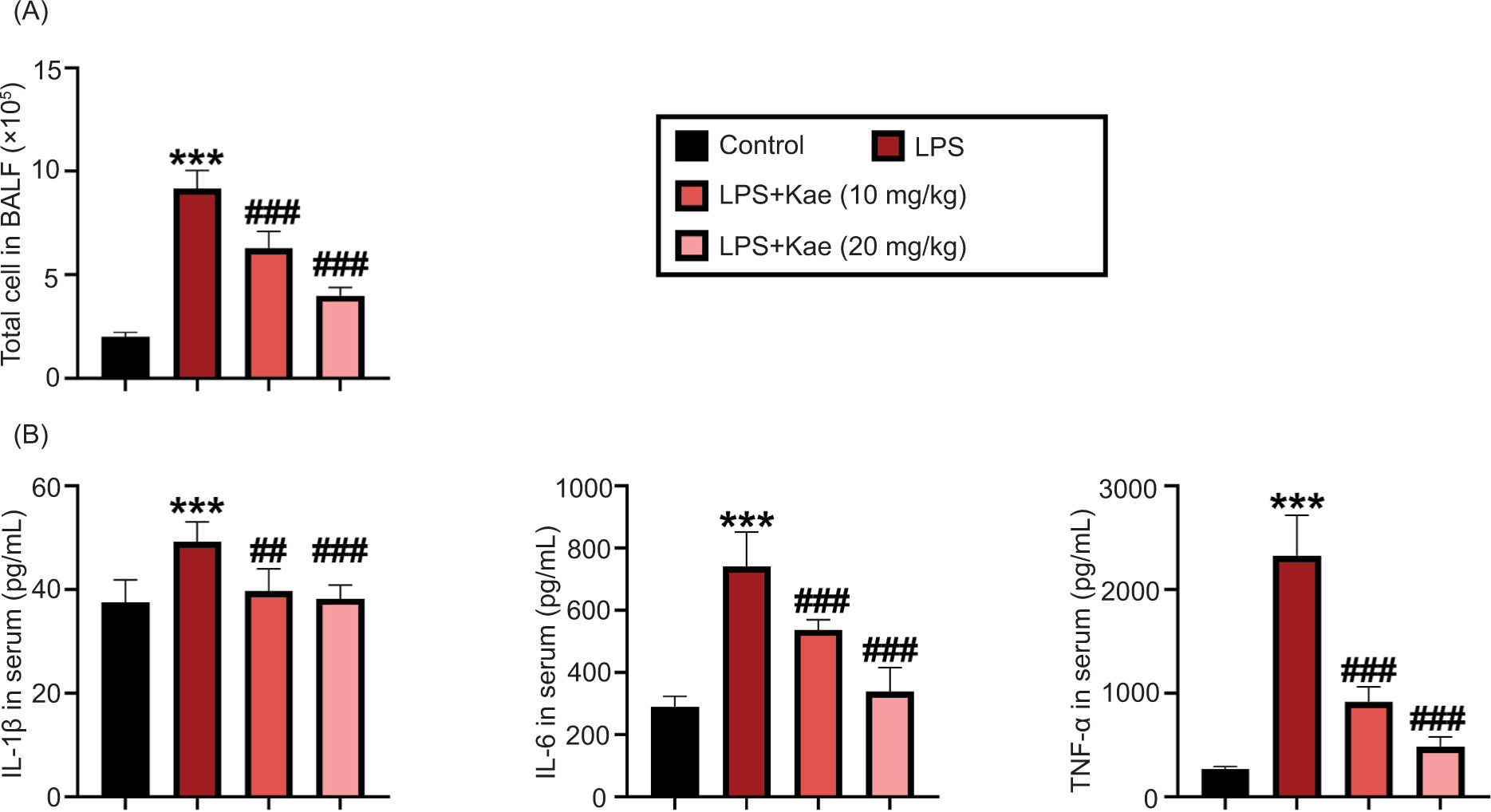
Figure 2 Kaempferitrin improves LPS-induced mouse pneumonia factor. (A) Total cells count in BALF in different groups. (B) Pro-inflammatory cytokines, TNF-α, IL-1β, and IL-6, levels in serum determined by ELISA. *P < 0.05 vs control group. *P < 0.005, **P < 0.001, ***P < 0.0001 (n = 3), #P < 0.005, ##P < 0.001, ###P < 0.0001 (n = 3). *LPS group, compared with the control group; #Kae+LPS group, compared with the LPS group.
Kaempferitrin lessens LPS-induced pneumonia in mice
Next, the effect of Kae on LPS-induced inflammatory response was explored. First, the total cells count was discovered in BALF. As shown in Figure 2A, the total cells count was significantly increased by LPS in BALF, compared to the control group. However, the total cells count was evidently suppressed by the treatment of Kae in both Kae+LPS (Kae 10 mg/kg) and Kae+LPS (Kae 20 mg/kg) groups, compared with the LPS-treated group. Additionally, TNF-α, IL-1β, and IL-6 levels in BALF cells count was determined by ELISA. LPS administration led to a significant increase in TNF-α, IL-1β, and IL-6 levels in BALF (Figure 2B) whereas Kae administration significantly restrained TNF-α, IL-1β, and IL-6 levels induced by LPS. These results indicated that Kae had a protective effect against pneumonia in ALI by repressing the production of inflammatory factors.
Kaempferitrin lessens LPS-induced apoptosis in mouse lung tissue
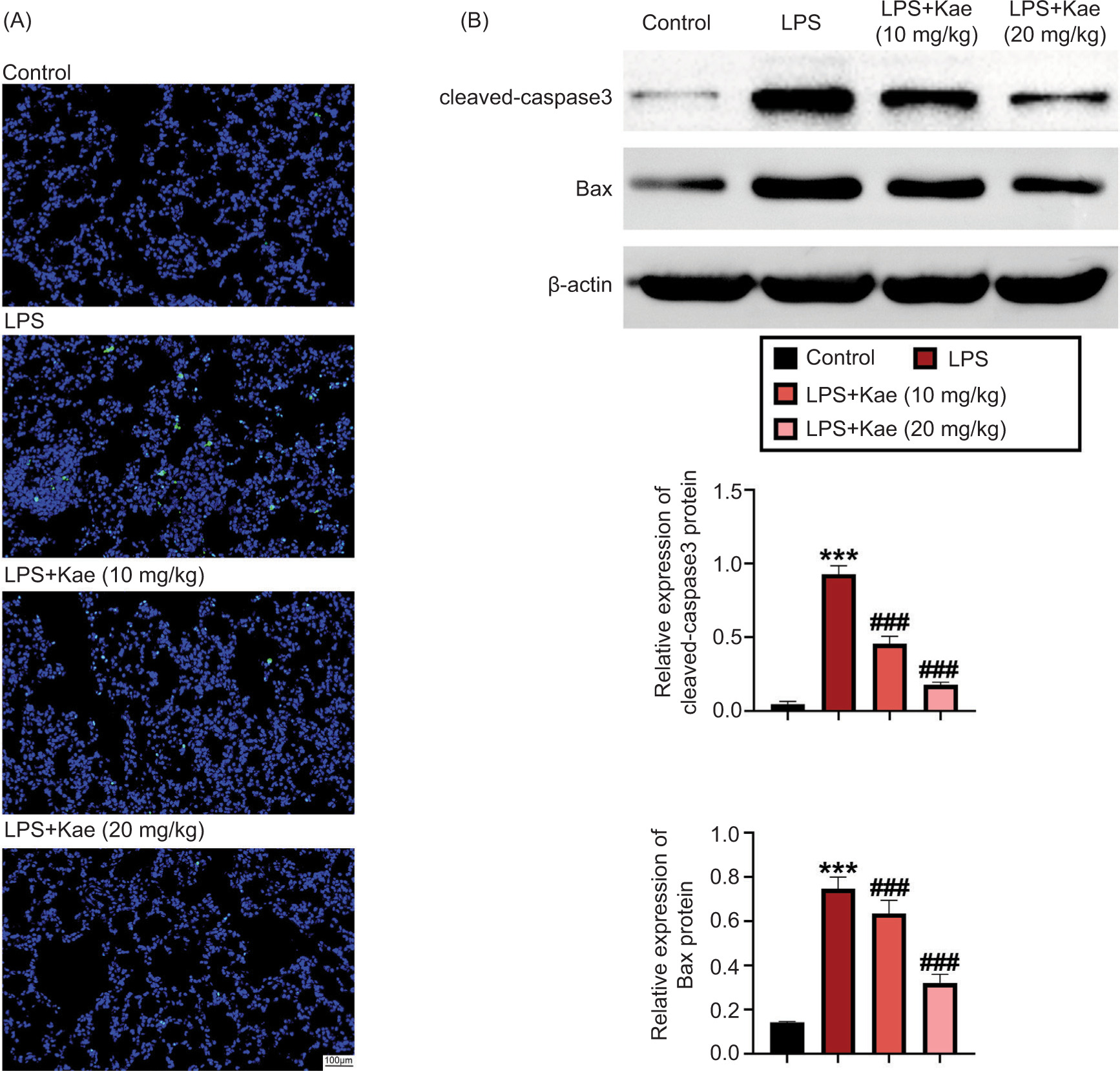
Apoptosis is another important pathological feature of ALI.20 We next explored the effect of Kae on apoptosis in LPS-induced ALI mice. The result showed that the number of TUNEL-positive cells was significantly upregulated by LPS, compared to the control group.
Nevertheless, Kae treatment evidently decreased the number of TUNEL-positive cells induced by LPS (Figure 3A). Cleaved capase-3 and Bax are important markers of apoptosis in the development of ALI.21 Therefore, the levels of cleaved capase-3 and Bax were observed, and the results showed that the expression levels of Bax and cleaved capase-3 were obviously increased in the LPS group, compared to the control group. Nonetheless, both Kae+LPS (Kae 10 mg/kg) and Kae+LPS (Kae 20 mg/kg) groups showed decreased levels of Bax and cleaved capase-3 induced by LPS (Figure 3B). Hence, the results established that Kae attenuated LPS-induced apoptosis in ALI mice.
Figure 3 Kaempferitrin improves LPS-induced apoptosis in mouse lung tissue. (A) Apoptotic cells in lung tissues were observed by TUNEL. (B) The protein expression levels of Bax and cleaved caspase-3 were discovered by Western blot analysis. *P < 0.05 vs control group. *P < 0.005, **P < 0.001, ***P < 0.0001 (n = 3). #P < 0.005, ##P < 0.001, and ###P < 0.0001 (n = 3). *LPS group, compared with the control group; #Kae+LPS group, compared with the LPS group.
Kaempferitrin inhibits activation of NF-κB pathway
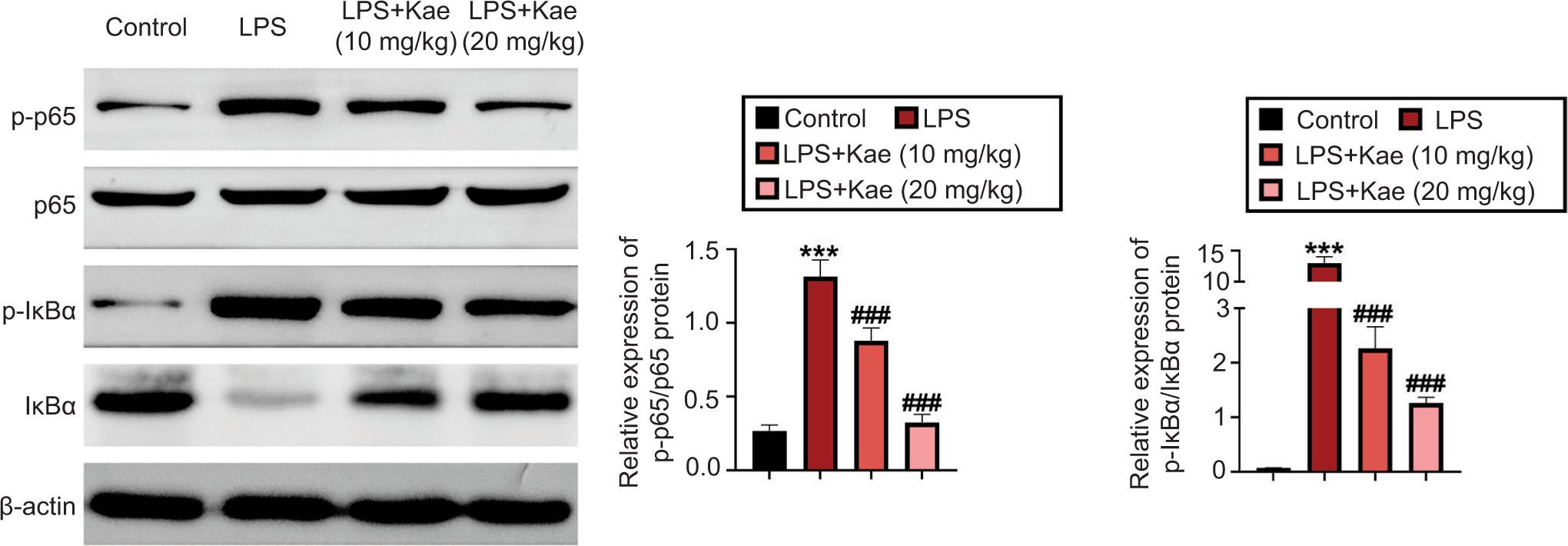
Finally, the mechanism of Kae on the NF-κB pathway was explored in lung tissue. Here, the proteins related to the NF-κB pathway were observed, including transcription nuclear factors p-p65, p65, IκBα, and p-IκBα. The proteins of lung tissues were extracted and subjected to Western blot analysis for testing p-p65, p65, IκBα, and p-IκBα. The protein levels of p-p65/p65 and p-IκBα/IκBα were evidently upregulated in the LPS group, compared to the control group (Figure 4). On the contrary, Kae treatment decreased the levels of p-p65/p65 and p-IκBα/IκBα in both Kae+LPS (Kae 10 mg/kg) and Kae+LPS (Kae 20 mg/kg) groups. These results emphasized that the LPS-induced inflammatory response was suppressed by Kae by diminishing the level of pro-inflammatory cytokines by inhibiting the activation of NF-κB pathway.
Figure 4 The mechanism study of Kae on NF-κB pathway. The protein expression levels of p-P56, p-56, p-IKBα, and IKBα were discovered by Western blot analysis. *P < 0.05 vs control group. *P < 0.005, **P < 0.001, and ***P < 0.0001 (n = 3). #P < 0.005, ##P < 0.001, and ###P < 0.0001 (n = 3). *LPS group, compared with the control group; #Kae+LPS group, compared with the LPS group.
Discussion
In spite of the advantages of understanding the pathogenesis of sepsis, clinical treatments still have limitations. ALI was the most common organ injury in sepsis, which can lead to severe diseases of the lung and lead to uncontrolled and self-amplified pulmonary inflammation.22 The treatments available for ALI in clinical practice are limited to supportive treatment. Therefore, it is of great value and significance to discover a new drug for treating ALI.8 By establishing ALI model using LPS, the action of Kae on ALI was explored by spotting specific characteristic proteins and pathways.
Recently, a number of components extracted from herbs have been demonstrated to attenuate ALI by regulating different pathways. For instance, glycyrrhizic acid can regulate PI3K/AKT/mTOR pathway of LPS-induced inflammation in ALI.23 The factors of inducible nitric oxide synthase (iNOS), NF-κB, and mitogen-activated protein kinase (MAPK) signaling were regulated by Picrasma quassioides in LPS-induced ALI.24 Emodin restrained pulmonary inflammation induced by LPS through regulation of NF-κB pathway.25 Our study showed that Kae could suppress NF-κB and Akt/mTOR pathway activation.26 Meanwhile, inhibition of TLR4/NF-κB pathway attenuated the injury and inflammation of lung tissues in ALI.27 Thus, TLR4/NF-κB pathway plays an important role in LPS infections. In this study, we confirmed that Kae could inhibit the protein expression levels of transcription factors p65, p-p65, IκBα, and p-IκBα, thus attenuating the activating of NF-κB pathway induced by LPS. These findings suggested that Kae attenuated ALI by blocking NF-κB–hedgehog pathway.
Conclusion
The results of this study revealed the multiple functions of Kae in constricting LPS-induced ALI in mice through its anti-inflammatory and anti-apoptosis effects. Mechanistic studies showed that Kae repressed inflammatory response by inhibiting NF-κB pathway activation, indicating that Kae might be a potential therapeutic component to treat ALI.
Funding
No funding was used in this study.
Conflict of Interest
The authors stated that there was no conflict of interest to disclose.
Data Availability
The authors declare that all data supporting the findings of this study are available within the paper and any raw data can be obtained from the corresponding author upon request.
Author Contributions
Xiaoli Zhu, Yongyue Pan, Xin Xu, and Jing Xu designed and carried out the study. All the authors supervised data collection, and analyzed and interpreted the collected data. They prepared the manuscript for publication and reviewed the draft. All the authors read and approved the final manuscript.
REFERENCES
1. Mehdi SF, Pusapati S, Khenhrani RR, Farooqi MS, Sarwar S, Alnasarat A, et al. Oxytocin and related peptide hormones: Candidate anti-inflammatory therapy in early stages of sepsis. Front Immunol. 2022;13:864007. 10.3389/fimmu.2022.864007
2. Molloy EJ, Bearer CF. Paediatric and neonatal sepsis and inflammation. Pediatr Res. 2022;91(2):267–9. 10.1038/s41390-021-01918-4
3. Gustot T. Multiple organ failure in sepsis: Prognosis and role of systemic inflammatory response. Curr Opin Crit Care. 2011;17(2):153–9. 10.1097/MCC.0b013e328344b446
4. O’Toole SA, Beith JM, Millar EK, West R, McLean A, Cazet A, et al. Therapeutic targets in triple negative breast cancer. J Clin Pathol. 2013;66(6):530–42. 10.1136/jclinpath-2012-201361
5. Wu C, Li H, Zhang P, Tian C, Luo J, Zhang W, et al. Lymphatic flow: A potential target in sepsis-associated acute lung injury. J Inflamm Res. 2020;13:961–8. 10.2147/JIR.S284090
6. Ge P, Luo Y, Okoye CS, Chen H, Liu J, Zhang G, et al. Intestinal barrier damage, systemic inflammatory response syndrome, and acute lung injury: A troublesome trio for acute pancreatitis. Biomed Pharmacother. 2020;132:110770. 10.1016/j.biopha.2020.110770
7. Bellani G, Laffey JG, Pham T, Fan E, Brochard L, Esteban A, et al. Epidemiology, patterns of care, and mortality for patients with acute respiratory distress syndrome in intensive care units in 50 countries. JAMA. 2016;315(8):788–800. 10.1001/jama.2016.0291
8. Butt Y, Kurdowska A, Allen TC. Acute lung injury: A clinical and molecular review. Arch Pathol Lab Med. 2016;140(4):345–50. 10.5858/arpa.2015-0519-RA
9. Wang M, Cai Q. Phillygenin attenuates LPS-induced acute lung injury of newborn mice in infantile pneumonia. Signa Vitae 2021;17(4):171–7. 10.22514/sv.2021.085
10. Nezic L, Amidzic L, Skrbic R, Gajanin R, Mandic D, Dumanovic J, et al. Amelioration of endotoxin-induced acute lung injury and alveolar epithelial cells apoptosis by simvastatin is associated with up-regulation of survivin/NF-kB/p65 pathway. Int J Mol Sci. 2022;23(5):2596. 10.3390/ijms23052596
11. Zhu J, Dong X. Decursin alleviates LPS-induced lung epithelial cell injury by inhibiting NF-kappaB pathway activation. Allergol Immunopathol (Madr). 2023;51(1):37–43. 10.15586/aei.v51i1.689
12. Liu FJ, Gu TJ, Wei DY. Emodin alleviates sepsis-mediated lung injury via inhibition and reduction of NF-kB and HMGB1 pathways mediated by SIRT1. Kaohsiung J Med Sci. 2022;38(3):253–60. 10.1002/kjm2.12476
13. Vallabhapurapu S, Karin M. Regulation and function of NF-kappaB transcription factors in the immune system. Annu Rev Immunol. 2009;27:693–733. 10.1146/annurev.immunol.021908.132641
14. Bae HB, Li M, Kim JP, Kim SJ, Jeong CW, Lee HG, et al. The effect of epigallocatechin gallate on lipopolysaccharide-induced acute lung injury in a murine model. Inflammation. 2010;33(2):82–91. 10.1007/s10753-009-9161-z
15. Wang J, Zhao Q. Kaempferitrin inhibits proliferation, induces apoptosis, and ameliorates inflammation in human rheumatoid arthritis fibroblast-like synoviocytes. Phytother Res. 2019;33(6):1726–35. 10.1002/ptr.6364
16. Calderon-Montano JM, Burgos-Moron E, Perez-Guerrero C, Lopez-Lazaro M. A review on the dietary flavonoid kaempferol. Mini Rev Med Chem. 2011;11(4):298–344. 10.2174/138955711795305335
17. Tie F, Ding J, Hu N, Dong Q, Chen Z, Wang H. Kaempferol and kaempferide attenuate oleic acid-induced lipid accumulation and oxidative stress in HepG2 cells. Int J Mol Sci. 2021;22(16):8847. 10.3390/ijms22168847
18. Li J, Lu K, Sun F, Tan S, Zhang X, Sheng W, et al. Panaxydol attenuates ferroptosis against LPS-induced acute lung injury in mice by Keap1-Nrf2/HO-1 pathway. J Transl Med. 2021;19(1):96. 10.1186/s12967-021-02745-1
19. Wang N, Li Q, Liu H, Lin L, Han W, Hao W. Role of C/EBP alpha hypermethylation in diesel engine exhaust exposure-induced lung inflammation. Ecotoxicol Environ Saf. 2019;183:109500. 10.1016/j.ecoenv.2019.109500
20. Ferguson ND, Fan E, Camporota L, Antonelli M, Anzueto A, Beale R, et al. The Berlin definition of ARDS: an expanded rationale, justification, and supplementary material. Intensive Care Med. 2012;38(10):1573–82. 10.1007/s00134-012-2682-1
21. Qu L, Chen C, He W, Chen Y, Li Y, Wen Y, et al. Glycyrrhizic acid ameliorates LPS-induced acute lung injury by regulating autophagy through the PI3K/AKT/mTOR pathway. Am J Transl Res. 2019;11(4):2042–55.
22. Lee JW, Park JW, Shin NR, Park SY, Kwon OK, Park HA, et al. Picrasma quassiodes (D. Don) Benn. attenuates lipopolysaccharide (LPS)-induced acute lung injury. Int J Mol Med. 2016;38(3):834–44. 10.3892/ijmm.2016.2669
23. Xiao M, Zhu T, Zhang W, Wang T, Shen YC, Wan QF, et al. Emodin ameliorates LPS-induced acute lung injury, involving the inactivation of NF-kappaB in mice. Int J Mol Sci. 2014;15(11):19355–68. 10.3390/ijms151119355
24. Cai M, Zhuang W, Lv E, Liu Z, Wang Y, Zhang W, et al. Kaemperfol alleviates pyroptosis and microglia-mediated neuroinflammation in Parkinson’s disease via inhibiting p38MAPK/NF-kappaB signaling pathway. Neurochem Int. 2022;152:105221. 10.1016/j.neuint.2021.105221
25. Ciesielska A, Matyjek M, Kwiatkowska K. TLR4 and CD14 trafficking and its influence on LPS-induced pro-inflammatory signaling. Cell Mol Life Sci. 2021;78(4):1233–61. 10.1007/s00018-020-03656-y