
Download
ORIGINAL ARTICLE
PHLDA1 knockdown inhibits inflammation and oxidative stress by regulating JNK/ERK pathway, and plays a protective role in sepsis-induced acute kidney injury
Minmin Gong*, Wenmei Liang, Qinju Lu, Jing Zhang
Department of Critical Care Medicine, Affiliated Hospital of Zunyi Medical University, Zunyi, Guizhou, China
Abstract
Background: Acute kidney injury (AKI), a prevalent complication of sepsis, causes substantial burden on patients’ families as well as the society. More reliable markers are urgently required for the prevention and treatment of AKI. Pleckstrin homology-like domain, family A, member 1 (PHLDA1) was implicated in various diseases, but its involvement in sepsis-induced AKI remains to be explored. The JNK/ERK pathway has been revealed as being involved in progression of sepsis. One previous study demonstrated that PHLDA1 could activate the JNK/ERK pathway in hepatic ischemia/reperfusion injury. Nevertheless, involvement of PHLDA1 in sepsis-triggered AKI through the JNK/ERK pathway has not been probed.
Methods: A cecal ligation and punctured (CLP) mice model of sepsis-induced AKI was established. Reverse transcription-quantitative polymerase chain reaction (RT-qPCR) and immunofluorescence staining were applied to evaluate the expression of PHLDA1. Concentration of blood urea nitrogen (BUN) and serum creatinine (Scr), inflammation markers, including interleukin-6 (IL-6), IL-1β, and tumor necrosis factor-α, as well as oxidative stress-associated proteins (catalase, malondialdehyde, superoxide dismutase, and glutathione), in the kidney tissues of mice were evaluated by enzyme-linked-immunosorbent serologic assay. Western blot analysis was applied for measuring protein expression levels.
Results: The BUN and SCr levels in mice were obviously elevated in the CLP group compared to the sham group. Moreover, the expression of PHLDA1 was also elevated in the CLP group in comparison to the sham group. Down-regulation of PHLDA1 alleviated renal injury, inflammation, and oxidative stress in AKI model. Mechanistic study showed that PHLDA1 knockdown suppressed the activation of c-JUN N-terminal kinase/p38 and extracellular signal-regulated kinase (JNK/ERK) pathway.
Conclusion: Down-regulation of PHLDA1 suppressed inflammation and oxidative stress through the modulation of JNK/ERK pathway in sepsis-induced AKI. The results could offer a novel insight into the treatment of patients with sepsis-induced AKI.
Key words: acute kidney injury, JNK/ERK pathway, PHLDA1, sepsis
*Corresponding author: Minmin Gong, Department of Critical Care Medicine, Affiliated Hospital of Zunyi Medical University, No. 149, Dalian Road, Huichuan District, Zunyi, Guizhou 563003, China. Email address: [email protected]
Received 17 May 2022; Accepted 10 June 2022; Available online 1 November 2022
Copyright: Gong M, et al.
License: This open access article is licensed under Creative Commons Attribution 4.0 International (CC BY 4.0). http://creativecommons.org/licenses/by/4.0/
Introduction
Sepsis is a response to infection presented as a whole-body inflammation, and can lead to severe damage to tissues and organs.1,2 Previous data have revealed that sepsis has become the leading cause of death in intensive care units (ICUs).3 Acute kidney injury (AKI) is a prevalent complication of sepsis, and in ICUs, more than 50% AKI patients were admitted due to sepsis.4,5 Although clinical and intensive care in ICUs provided to AKI patients has improved, still no effective therapy is available for the treatment of AKI patients, and their prognosis remains unfavorable.4,6 Acute renal failure occurs in almost 5% of AKI patients, and morality of AKI patients is approximately 60%; among 15% of survivors, renal replacement therapy (RRT) after discharge is still required.7 AKI causes substantial burden on patients’ families as well as the society,8,9 hence more reliable markers for the prevention and treatment of AKI are urgently required.
Pleckstrin homology-like domain, family A, member 1 (PHLDA1) is found to be a potential transcription factor located on 12q21.2, and is required for Fas expression and mouse T cell hybridoma apoptosis activation.10,11 By interacting with membrane components, PHL protein domain contained in PHLDA1 is involved in eliciting cellular responses by regulating the transduction of cell signaling, trafficking of vesicular, and rearrangement of cytoskeletons.12 Previous studies have revealed that PHLDA1 also participates in several biological processes, including cell proliferation, apoptosis, and differentiation.13 In addition, PHLDA1 is engaged in the oxygen glucose deprivation and reperfusion (OGD/R)-mediated neurons by modulating GSK-3β/Nrf2 pathway.14 PHLDA1 is majorly expressed in ulcerative colitis, which could provide a novel biomarker for managing dysplasia.15 PHLDA1 is involved in human ovarian cancer cells by modulating endoplasmic reticulum stress response as well as oxidative stress.16 PHLDA1 participates in hepatic ischemia/reperfusion injury as a target of miR-194 by regulating TRAF6.17 Although the function of PHLDA1 has been corroborated in various diseases, whether it is implicated in sepsis-induced AKI remains to be explored.
Here, the role and function of PHLDA1 in sepsis--induced AKI was investigated, and it was observed that PHLDA1 silencing suppressed inflammation and oxidative stress in sepsis-induced AKI through modulating the c-JUN N-terminal kinase/p38 and extracellular signal-regulated kinase (JNK/ERK) pathway. The findings of our study might highlight the role of PHLDA1 in the future treatment of -sepsis-induced AKI for improving the prognosis of AKI patients.
Methods
Animal model
The animal experiments were conducted with the approval of the Ethics Committee of Affiliated Hospital of Zunyi Medical University (Approval No. KLLY (A) -2019-047). Vital River (Beijing, China) provided 6–8-week-old C57BL/6 mice (weighing 20–25 g). All animals were kept in individual cages having a temperature of 25 ± 2°C under a 12-h light/dark cycle for one week for environmental adaptation, and were provided with free water and food. Cecal ligation and puncture (CLP) surgery was performed in mice to trigger sepsis.18 The animals were subjected to preheated normal saline, and anesthetized by isoflurane. A 1–2-cm incision was made at the midline of the abdomen after cleaning, and the cecum was removed followed by ligation using a sterile silk suture and double punctured with a 20-gauge needle. Post-squeezing, the cecum was put back into the abdominal cavity and the incision was closed. In the sham group, the procedures were repeated without CLP operation. After 72 h, the blood and kidney tissues of mice were harvested for further use.
The animals (n = 24) were divided into the following four groups (each group, n = 6): (1) Sham group; (2) CLP group; (3) CLP+adeno-associated virus (AAV) group; and (4) CLP+AAV-short hairpin PHLDA1 (shPHLDA1) group.
Detection of blood urea nitrogen (BUN), serum creatinine (Scr), inflammatory markers, and -oxidative stress-associated proteins
Blood samples of mice were collected from jugular vein and centrifuged (10 min at 3000 rpm) to obtain serum for measuring the levels of BUN, Scr, and inflammatory markers.19,20 The BUN and Scr levels were tested, respectively, by the BUN assay kit (Jiancheng Biotech, Nanjing, China) and the creatinine assay kit (Biosino Bio-Technology, Beijing, China).
Inflammation markers, including interleukin-6 (IL-6), IL-1β, and tumor necrosis factor-alpha (TNF-α), were evaluated by specific commercial enzyme-linked-immunosorbent serologic assay (ELISA) kit (Sigma-Aldrich, USA).
The oxidative stress-associated proteins (catalase [CAT], malondialdehyde [MDA], superoxide dismutase [SOD], and glutathione [GSH]) in the kidney tissues of mice were assessed by using commercial kits (Jiancheng Biotech, Nanjing, China) according to respective instructions.21,22
Immunofluorescence (IF) staining
The kidney tissues of mice were maintained at room temperature for 1 h in a box of paraformaldehyde (PFA 1%; Sigma-Aldrich) and Triton X-100 (0.2%; Sigma-Aldrich). Post washing by droplets, the tissues were blocked in bovine serum albumin (BSA, 3%) solution and fetal bovine serum (FBS)/phosphate buffer solution (PBS, 10%) overnight at 4°C. Next, the tissues were incubated with the primary antibodies of PHLDA1 (1:50, ab133654; Abcam, Shanghai, China) overnight at 4°C. The secondary anti-rabbit antibodies (1:400; Sigma-Aldrich) were added and incubated at 4°C post-washing. A fluorescence microscope (Olympus, Japan) was used for observing the images.
Hematoxylin and eosin (H&E) staining
Xylene was used for dewaxing of paraffin-embedded kidney tissues of mice followed by hematoxylin staining for 10 min. The tissues were rinsed in distilled water and subjected to eosin staining for 2 min. Then, 95% and 100% ethanol was added to tissues for 1 min. Subsequently, the sections were mounted with neutral resin, and images were observed under microscope (Olympus, Japan).
RNA extraction and quantitative reverse -transcription-polymerase chain reaction (RT-qPCR)
The TRIzol reagent (Invitrogen, Carlsbad, USA) was applied for RNA extraction from the kidney tissues of mice. The PrimeScript RT reagent kit (Takara, Dalian, China) was used for the synthesis of single-stranded complementary DNA (cDNA). RT-qPCR was performed using the standard SYBR Green PCR kit (Toyobo Life Science, Shanghai, China) and executed on the ABI Prism 7500 Sequence Detection System (PerkinElmer Inc., USA). Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) acted as a control, and the relative level of PHLDA1 was analyzed using the 2-ΔΔCt method.
The primers were as follows:
PHLDA1:
F: 5'-CCAGGACAGATGCTACTTGG-3'
R: 5'-GACTACATAACCTAGCAGTGG-3';
GAPDH:
F: 5'-AACGTGTCAGTGGTGGACCTG-3'
R: 5'-AGTGGGTGTCGCTGTTGAAGT-3'.
Transfection of adenovirus
Adenovirus-mediated AAV-shPHLDA1 was transfected into mice for the knockdown of PHLDA1 level. AAV-shPHLDA1 was transfected into the abdominal cavity of mice 4 days prior to CLP. After anesthetization, the mice were intraperitoneal injected with AAV-shPHLDA1. After transfection, level of PHLDA1 was measured by Western blot analysis.
Western blot analysis
Protein samples were extracted from the kidney tissues and prepared in sodium dodecyl sulfate (SDS) buffer followed by their subjecting to sodium dodecyl sulfate–-polyacrylamide gel electrophoresis (SDS-PAGE). The concentration of proteins was measured by a bicinchoninic acid (BCA) assay kit (Thermo Fisher Scientific, USA). Equal amount of total proteins was loaded for SDS-PAGE in each lane and transferred on polyvinylidene fluoride (PVDF) membranes. Non-fat milk was applied for sealing the membranes, and primary antibodies were added to the membranes for one night. Then horseradish peroxidase (HRP)-conjugated secondary antibodies (Sigma-Aldrich, USA) were supplemented for 2 h. Bands were normalized with GAPDH, and signal intensities of the bands were quantified by Image J (NIH, Bethesda, MD, USA). An enhanced chemiluminesence system (Amersham Biosciences, USA) and the ImageLab software (Bio-Rad, USA) were employed, respectively, for the visualization and quantification of band signals. Primary antibodies were as follows: anti-PHLDA1 (1:10,000, ab133654; Abcam, Shanghai, China), anti-nuclear factor erythroid 2-related factor 2 (Nrf2) (1:1000, ab92946; Abcam), anti--Cyclooxygenase-2 (COX-2) (1:1000, ab179800; Abcam), anti-inducible nitric oxide synthase (iNOS) (1:1000, ab178945; Abcam), anti-p65 (1:1000, ab32536; Abcam), anti-p-p65 (1:1000, ab76302; Abcam), anti-JNK (1:1000, ab76125; Abcam), anti-p-JNK(1:5000, ab76572; Abcam), anti-ERK (1:1000, ab32537; Abcam), anti-p-ERK (1:1000, ab201015; Abcam), and anti-GAPDH (1:1000, ab9483; Abcam).
Statistical analysis
The Statistical Product and Service Solutions 13.0 software (IBM Inc., New York, USA) was applied for statistical analysis expressed as mean ± standard error (SE). Unpaired two-tail Student’s t-test and one-way analysis of variance (ANOVA) were applied for comparison of differences between groups. All the experiments were conducted for at least three times, and P ˂ 0.05 was considered as statistically significant.
Results
PHLDA1 is enhanced in the renal tissues of AKI mice
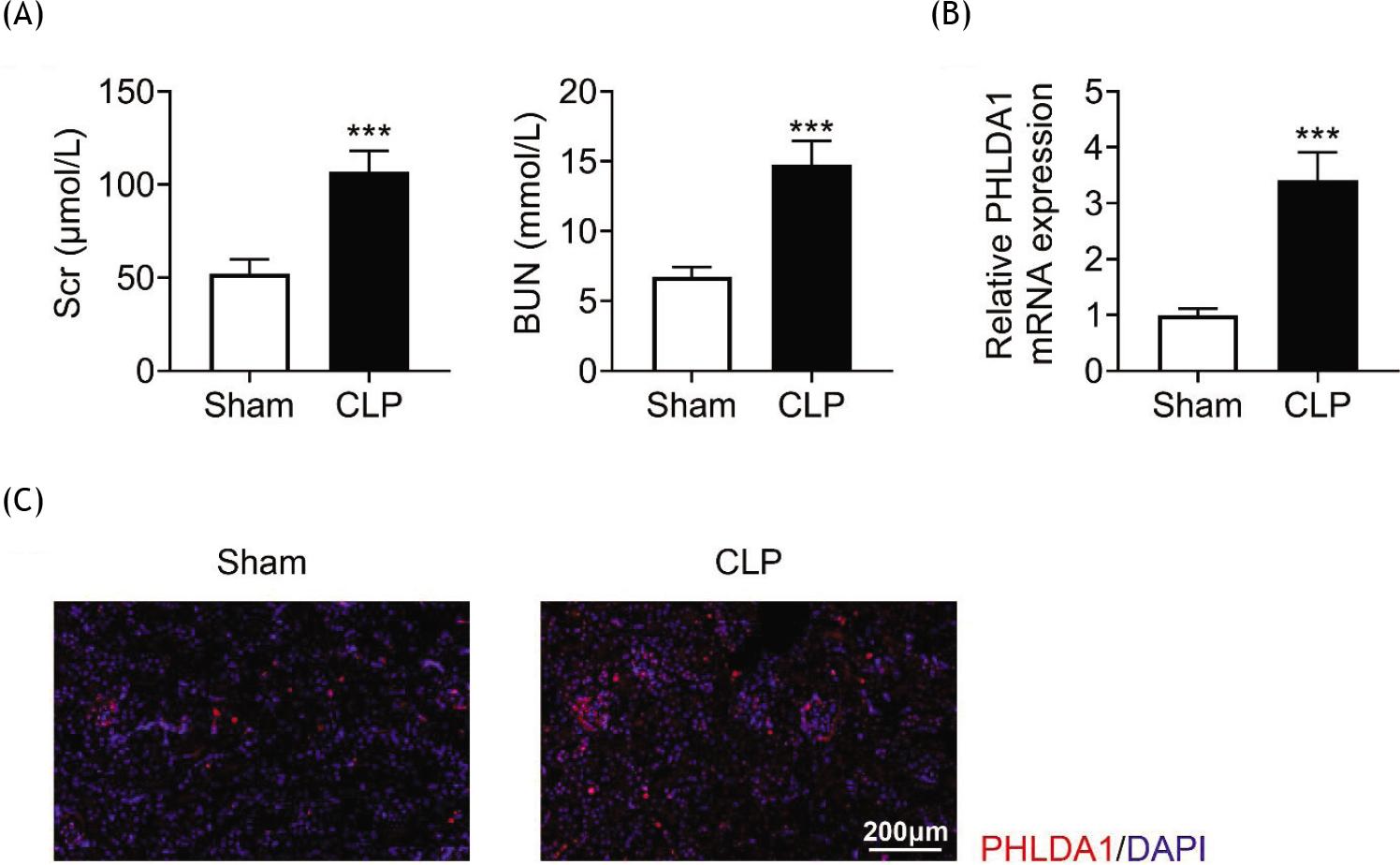
In order to probe the function of PHLDA1 in sepsis-induced AKI, a CLP mouse model of sepsis-induced AKI was established. The BUN and SCr levels of mice were measured. The results revealed that the BUN and SCr levels were elevated in the CLP group compared to the sham group (Figure 1A), suggesting that the AKI model was successfully constructed. RT-qPCR manifested that the PHLDA1 mRNA level was evidently enhanced in the CLP group (Figure 1B). Consistently, IF staining showed that the protein expression of PHLDA1 was also elevated in the CLP group compared to the sham group (Figure 1C). In a nutshell, PHLDA1 was enhanced in the renal tissues of AKI mice.
Figure 1 PHLDA1 is boosted in the renal tissues of AKI mice. Groups were separated as sham group (n = 6) and CLP group (n = 6). (A) The concentrations of BUN and SCr were evaluated via the corresponding commercial kits. (B) The level of PHLDA1 was revealed by RT-qPCR. (C) Immunofluorescence staining expressed the expression of PHLDA1. ***P ˂ 0.001 relative to sham group.
Silencing of PHLDA1 alleviates renal injury
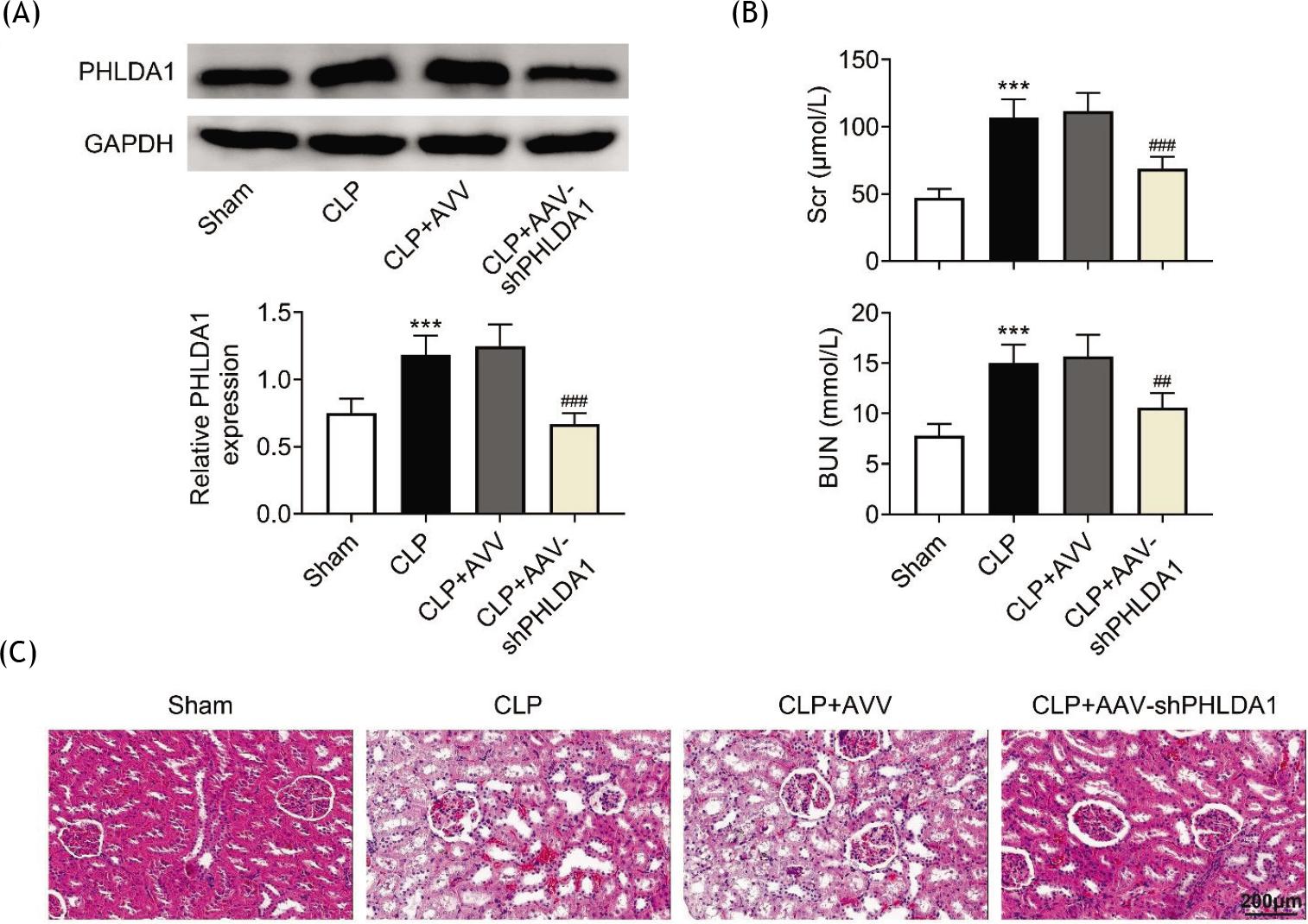
The function of PHILDA1 was studied by silencing it in the renal tissues of mice through the injection of AAV-shPHLDA1. The data demonstrated that PHLDA1 was increased in the CLP model, and the transfection of AAV-shPHLDA1 significantly reduced the level of PHLDA1 in the renal tissues of mouse model (Figure 2A). Moreover, down-regulation of PHLDA1 attenuated the levels of elevated BUN and SCr in the renal tissues of AKI model (Figure 2B). In addition, tubular injuries, including tubule dilation, loss of brush margin, cytoplasmic vacuole, and tubular formation, in the CLP group were alleviated by knockdown of PHLDA1 (Figure 2C). To summarize, silencing of PHLDA1 alleviated renal injury.
Figure 2 Silencing PHLDA1 alleviates renal injury. Groups were separated as sham group (n = 6), CLP group (n = 6), CLP+AAV group (n = 6), and CLP+AAV-shPHLDA1 group (n = 6). (A) The protein level of PHLDA1 was measured by Western blot analysis. (B) The concentrations of BUN and SCr were evaluated by corresponding commercial kits. (C) Hematoxylin and eosin staining uncovered the pathological changes of renal tissues. ***P ˂ 0.001 relative to sham group; ##P ˂ 0.01, ###P ˂ 0.001 relative to CLP+AAV group.
PHLDA1 knockdown reduces inflammation
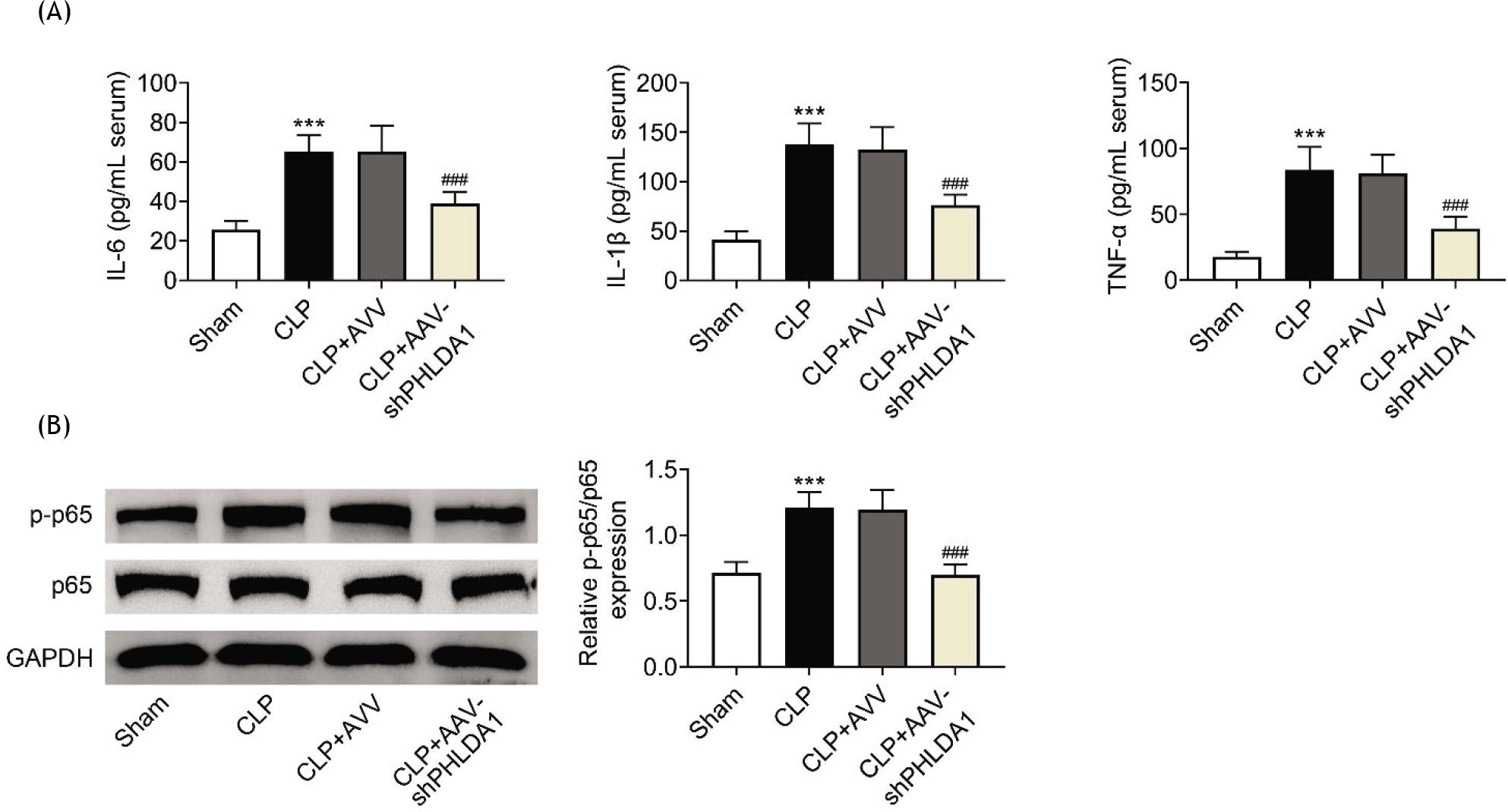
Next, the role of PHLDA1 in the inflammation of AKI was investigated. Concentrations of inflammation markers (IL-6, IL-β, and TNF-α) were evaluated. The results showed that the increased concentrations of IL-6, IL-β, and TNF-α in the CLP group were attenuated by PHLDA1 knockdown (Figure 3A). In addition, the p-p65 protein level was elevated in the CLP model whereas suppression of PHLDA1 inversely changed this effect (Figure 3B). In general, PHLDA1 knockdown reduced inflammation in AKI.
Figure 3 PHLDA1 knockdown reduces inflammation. Groups were separated as sham group (n = 6), CLP group (n = 6), CLP+AAV group (n = 6), and CLP+AAV-shPHLDA1 group (n = 6). (A) The concentrations of inflammation markers were detected by ELISA. (B) The protein levels of p-p65 and p65 were measured by Western blot analysis. ***P ˂ 0.001 relative to sham group; ###P ˂ 0.001 relative to CLP+AAV group.
PHLDA1 alleviation ameliorates oxidative stress
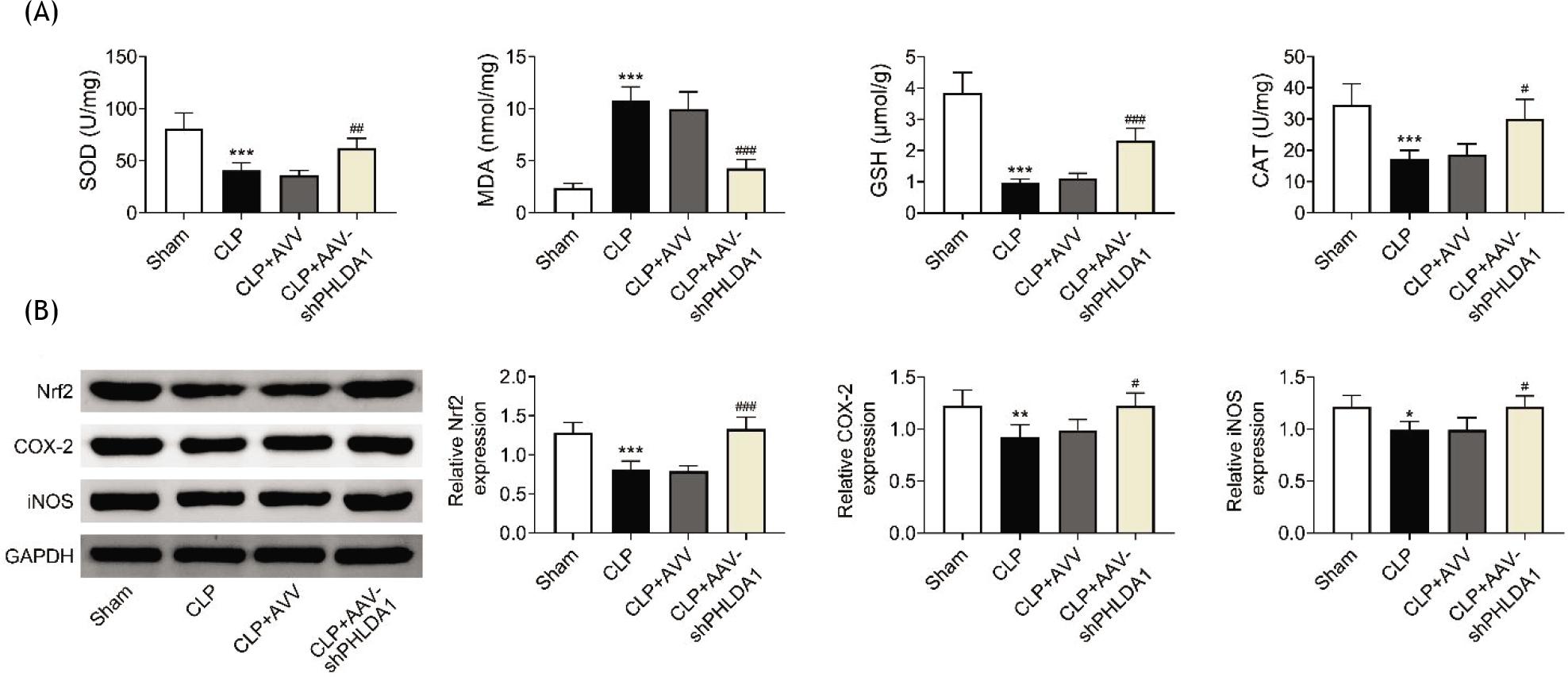
Furthermore, participation of PHLDA1 in oxidative stress in AKI was explored. ELISA was performed to detect the concentrations of oxidative stress-associated proteins (MDA, SOD, GSH, and CAT). CLP treatment decreased the concentrations of SOD, GSH, and CAT but increased the concentration of MDA; however, these effects were counteracted by down-regulation of PHLDA1 (Figure 4A). Moreover, the protein levels of Nrf2, COX-2, and iNOS were decreased in CLP mice, but silencing of PHLDA1 offset these effects (Figure 4B). These results showed that alleviation of PHLDA1 ameliorated oxidative stress in AKI.
Figure 4 PHLDA1 alleviation ameliorates oxidative stress. Groups were separated AS sham group (n = 6), CLP group (n = 6), CLP+AAV group (n = 6), and CLP+AAV-shPHLDA1 group (n = 6). (A) The concentrations of SOD, MDA, GSH, and CAT were assessed through the corresponding commercial kits. (B) Western blot analysis tested the protein levels of Nrf2, COX-2, and iNOS. *P ˂ 0.05, **P ˂ 0.01 ***P ˂ 0.001 relative to sham group; #P ˂ 0.05, ##P ˂ 0.01, ###P ˂ 0.001 relative to CLP+AAV group.
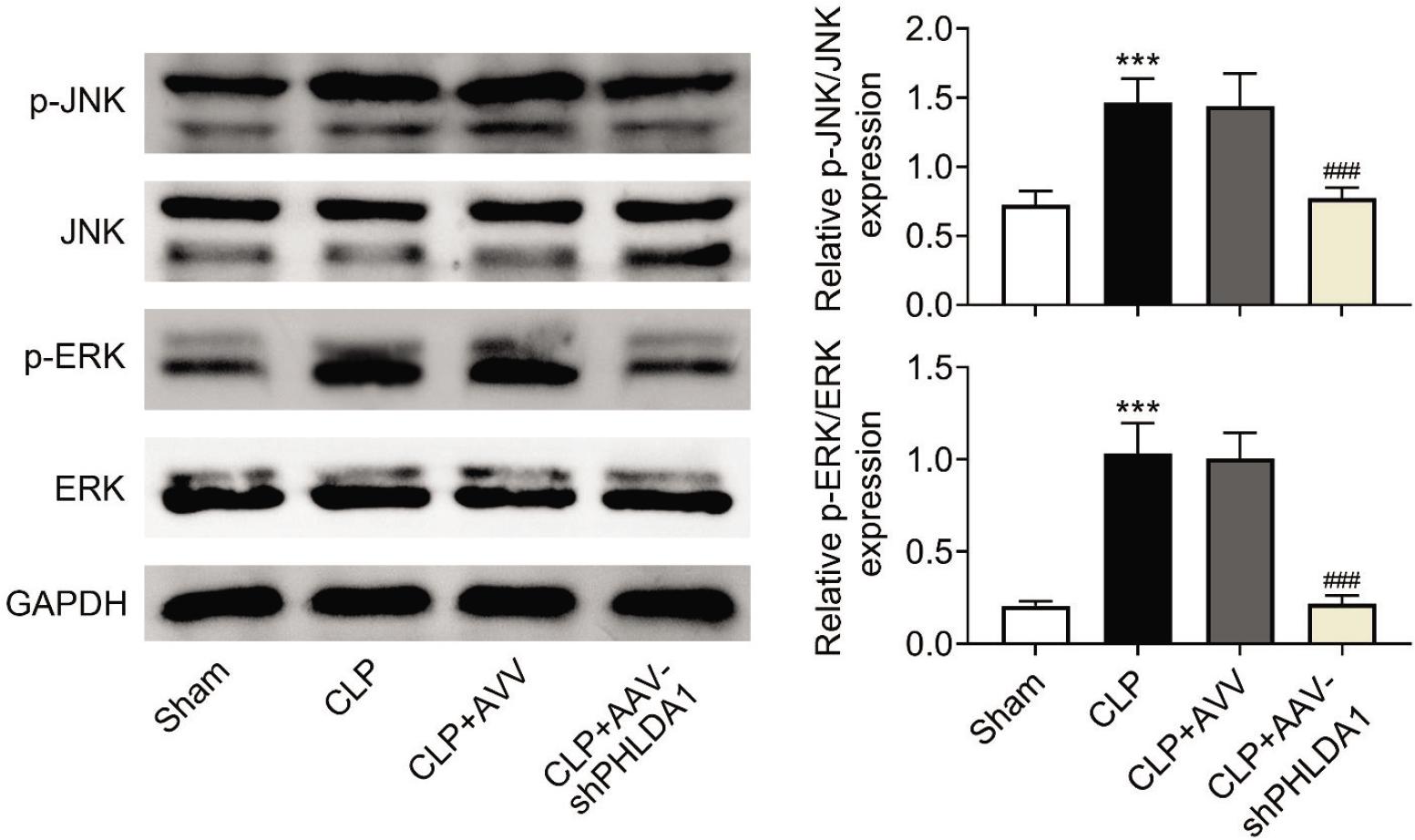
PHLDA1 knockdown suppresses JNK/ERK pathway activation
Finally, the underlying mechanism of PHLDA1 in AKI was investigated. It was observed that down-regulation of PHLDA1 reduced the CLP-induced elevation of the JNK/ERK pathway-related proteins (p-JNK and p-ERK; Figure 5). This result established that knockdown of PHLDA1 suppressed the activation of JNK/ERK pathway.
Figure 5 PHLDA1 knockdown suppresses JNK/ERK pathway activation. Groups were separated as sham group (n = 6), CLP group (n = 6), CLP+AAV group (n = 6), and CLP+AAV-shPHLDA1 group (n = 6). The protein levels of JNK, p-JNK, ERK, and p-ERK were examined through Western blot analysis. ***P ˂ 0.001 relative to sham group; ###P ˂ 0.001 relative to CLP+AAV group.
Discussion
As a prevalent complication of sepsis, AKI affects nearly 50% of patients with sepsis and the mortality rate of up to 60%.23,24 Nonetheless, the detailed pathogenesis of -sepsis-induced AKI is still unclear. In previous studies, multiple proteins were identified to be implicated in the development of sepsis-induced AKI. For instance, Bcl-2 serves as a target of microRNA-543 and is involved in developing sepsis-induced AKI.25 Nicotinamide adenine dinucleotide phosphate (NADPH) oxidase (NOX) 4 modulates sepsis-induced AKI by mediating apoptosis, inflammation, and oxidative stress in kidney tubular epithelial cells by regulating phosphatase and tensin homolog (PTEN) pathway.26 70 Kilodalton heat shock protein (HSP70) alleviates the inflammation-associated apoptosis in sepsis-induced AKI by combining with TNF receptor-associated factor 6 (TRAF6).27 Fatty acid-binding protein 4 mediates inflammatory response and cell apoptosis in sepsis-induced AKI, and might act as a novel therapeutic target for sepsis--induced AKI.28 PHLDA1 is reported to be involved in OGD/R-mediated neurons,14 ulcerative colitis and dysplasia,15 ovarian cancer,16 and hepatic ischemia/reperfusion injury.17 However, participation of PHLDA1 in sepsis-induced AKI remains to be explored. In the current study, a CLP-mice model of sepsis-induced AKI was established to identify the role and function of PHLDA1 in the disease. It was observed that PHLDA1 level was evidently enhanced in the CLP group compared to the sham group. In addition, PHLDA1 knockdown attenuated the elevated levels of BUN and SCr in the renal tissues of AKI model, demonstrating that silencing of PHLDA1 alleviated renal injury. Oxidative stress and inflammation are identified as critical processes that participate in sepsis.29–33 Similarly, in this study, we also discovered that PHLDA1 knockdown reduced inflammation and oxidative stress in sepsis-induced AKI. To summarize, PHLDA1 inhibition had a protective role in sepsis-induced AKI.
A variety of regulatory pathways are involved in the occurrence and development of sepsis. For instance, suppression of Bruton's tyrosine kinase (BTK) signaling alleviates sepsis-induced AKI in mice.34 Moreover, inhibition of IL-2-inducible T-cell kinase (ITK) signaling cuts down AKI progression in mice by modulating inflammatory transcription factor signaling.35 Besides, suppression of spleen tyrosine kinase (Syk) signaling relieves sepsis--induced AKI by modulating inflammatory cytokines.36 The JNK/ERK pathway has been extensively corroborated to participate in a variety of diseases. For example, the JNK/ERK pathway-mediated autophagy attenuation increases the cell death induced by Jaspine B (JB) derivative in gastric cancer by suppressing the p62/Keap1/Nrf2 pathway.37 Columbianadin ameliorates lipopolysaccharide (LPS)-triggered inflammation and oxidative stress in liver injury by modulating JNK/ERK pathway.38 JNK/ERK regulated by paeonol is implicated in anesthesia-mediated hippocampal neurotoxicity and histone acetylation in neonatal rat.39 Moreover, Fang et al. identified that JNK/ERK pathway mediates obesity-induced cardiomyopathy, fibrosis, cardiac injury, and remodeling.40 The JNK/ERK pathway also participates in the progression of sepsis.41–44 In addition, one previous study has demonstrated that PHLDA1 could activate JNK/ERK pathway in hepatic ischemia/reperfusion injury.17 Nevertheless, role of PHLDA1 in sepsis-triggered AKI through JNK/ERK pathway is rarely probed. Results of the present study established that the CLP-induced elevation of the JNK/ERK pathway-related proteins (p-JNK and p-ERK) was suppressed by PHLDA1 down-regulation, thereby resulting in suppressed JNK/ERK pathway activation in sepsis-triggered AKI.
Conclusion
The present study is the first one to demonstrate that PHLDA1 knockdown suppressed inflammation and oxidative stress through the modulation of JNK/ERK pathway in sepsis-induced AKI. The results could offer a novel insight in treating patients with sepsis-induced AKI.
Funding
This work was supported by the Master Research Foundation of Zunyi Medical College of Guizhou Province (Grant No. 2016-40).
Availability of Data and Materials
All data generated or analyzed during this study are included in this published article.
Competing Interests
The authors stated that there was no conflict of interest to disclose.
Author Contributions
Minmin Gong designed the study, completed the experiment and supervised the data collection, Wenmei Liang analyzed the data, interpreted the data Qinju Lu and Jing Zhang prepare the manuscript for publication and reviewed the draft of the manuscript. All authors have read and approved the manuscript.
REFERENCES
1. Souza DC, Jaramillo-Bustamante JC, Céspedes-Lesczinsky M, Quintero EMC, Jimenez HJ, Jabornisky R, et al. Challenges and health-care priorities for reducing the burden of paediatric sepsis in Latin America: A call to action. Lancet Child Adolesc Health. 2022;6(2):129–36. 10.1016/S2352-4642(21)00341-2
2. Li S, Sun Y, Tang X, Wang L, Cheng X. miR-20a attenuates acute lung injury in septic rats via targeting TLR4. Signa Vitae. 2021;17(4):157–62.
3. Zhang T, Xiang L. Honokiol alleviates sepsis-induced acute kidney injury in mice by targeting the miR-218-5p/heme oxygenase-1 signaling pathway. Cell Mol Biol Lett. 2019;24:15. 10.1186/s11658-019-0142-4
4. Kahraman D, Diniz G, Sayham S, Ersavas S, Ayaz D, Keskin E, et al. Over expressions of neutrophil gelatinase-associated lipocalin and kidney injury molecule-1 in human uterine cervical neoplasms enhance tumor invasion. Eur J Gynaecol Oncol. 2021;42(1):148–53. 10.31083/j.ejgo.2021.01.2195
5. Song C, Adili A, Kari A, Abuduhaer A. FSTL1 aggravates-sepsis-induced acute kidney injury through regulating TLR4/MyD88/NF-kappa B pathway in newborn rats. Signa Vitae. 2021;17(3):167–73.
6. Kalantari K, Rosner MH. Recent advances in the pharmacological management of sepsis-associated acute kidney injury. Expert Rev Clin Pharmacol. 2021;14(11):1401–11. 10.1080/17512433.2021.1978287
7. Jia Y, Li Z, Feng Y, Cui R, Dong Y, Zhang X, et al. Methane-rich saline ameliorates sepsis-induced acute kidney injury through anti-inflammation, antioxidative, and antiapoptosis effects by regulating endoplasmic reticulum stress. Oxidat Med Cell Longevity. 2018;2018:4756846. 10.1155/2018/4756846
8. Quaglia M, Merlotti G, Colombatto A, Bruno S, Stasi A, Franzin R, et al. Stem cell-derived extracellular vesicles as potential therapeutic approach for acute kidney injury. Front Immunol. 2022;13:849891. 10.3389/fimmu.2022.84989
9. Tatar B. Assessment of myometrial invasion of endometrial carcinoma with preoperative transvaginal sonography. Eur J Gynaecol Oncol. 2021;42(1):85–9. 10.31083/j.ejgo.2021.01.2236
10. Guo Y, Jia P, Chen Y, Yu H, Xin X, Bao Y, et al. PHLDA1 is a new therapeutic target of oxidative stress and ischemia-reperfusion-induced myocardial injury. Life Sci. 2020;245: 117347. 10.1016/j.lfs.2020.117347
11. Molodecky NA, Soon IS, Rabi DM, Ghali WA, Ferris M, Chernoff G, et al. Increasing incidence and prevalence of the inflammatory bowel diseases with time, based on systematic review. Gastroenterology. 2012;142(1):46–54.e42; quiz e30. 10.1053/j.gastro.2011.10.001
12. Scheffzek K, Welti S. Pleckstrin homology (PH) like domains–Versatile modules in protein–protein interaction platforms. FEBS Lett. 2012;586(17):2662–73. 10.1016/j.febslet.2012.06.006
13. Zhao P, Lu Y, Liu L. Correlation of decreased expression of PHLDA1 protein with malignant phenotype of gastric adenocarcinoma. Int J Clin Exp Pathol. 2015;8(5):5230–5.
14. Yang F, Chen R. Loss of PHLDA1 has a protective role in OGD/R-injured neurons via regulation of the GSK-3β/Nrf2 pathway. Human Exp Toxicol. 2021;40(11):1909–20. 10.1177/09603271211014596
15. Orita F, Ishikawa T, Ishiguro M, Okazaki S, Kikuchi A, Yamauchi S, et al. PHLDA1 expression in ulcerative colitis: A potential role in the management of dysplasia. Mol Clin Oncol. 2021;15(3):192. 10.3892/mco.2021.2354
16. Xu J, Bi G, Luo Q, Liu Y, Liu T, Li L, et al. PHLDA1 modulates the endoplasmic reticulum stress response and is required for resistance to oxidative stress-induced cell death in human ovarian cancer cells. J Cancer. 2021;12(18):5486–93. 10.7150/jca.45262
17. Luo YH, Huang ZT, Zong KZ, Cao ZR, Peng DD, Zhou BY, et al. miR-194 ameliorates hepatic ischemia/reperfusion injury via targeting PHLDA1 in a TRAF6-dependent manner. Int Immunopharmacol. 2021;96:107604. 10.1016/j.intimp.2021.107604
18. Zhang Z, Zhao H, Ge D, Wang S, Qi B. β-Casomorphin-7 ameliorates sepsis-induced acute kidney injury by targeting NF-κB pathway. Med Sci Monit Int Med J Exp Clin Res. 2019;25:121–7. 10.12659/MSM.912730
19. Li X, Zheng P, Ji T, Tang B, Wang Y, Bai S. LINC00052 ameliorates acute kidney injury by sponging miR-532-3p and activating the Wnt signaling pathway. Aging. 2020;13(1):340–50. 10.18632/aging.104152
20. Zhao W, Zhang L, Chen R, Lu H, Sui M, Zhu Y, et al. SIRT3 protects against acute kidney injury via AMPK/mTOR--regulated autophagy. Front Physiol. 2018;9:1526. 10.3389/fphys.2018.01526
21. Zhang N, Zhao W, Hu ZJ, Ge SM, Huo Y, Liu LX, et al. Protective effects and mechanisms of high-dose vitamin C on sepsis--associated cognitive impairment in rats. Sci Rep. 2021;11(1):14511. 10.1038/s41598-021-93861-x
22. Zhang C, Suo M, Liu L, Qi Y, Zhang C, Xie L, et al. Melatonin alleviates contrast-induced acute kidney injury by activation of Sirt3. Oxidat Med Cell Long. 2021;2021:6668887. 10.1155/2021/6668887
23. Plotnikov EY, Pevzner IB, Zorova LD, Chernikov VP, Prusov AN, Kireev II, et al. Mitochondrial damage and mitochondria--targeted antioxidant protection in LPS-induced acute kidney injury. Antioxidants (Basel, Switzerland). 2019;8(6):176. 10.3390/antiox8060176
24. Gao F, Qian M, Liu G, Ao W, Dai D, Yin C. USP10 alleviates sepsis-induced acute kidney injury by regulating Sirt6-mediated Nrf2/ARE signaling pathway. J Inflamm (London). 2021;18(1):25. 10.1186/s12950-021-00291-7
25. Zhang WQ, Wang HJ, Li YZ, Du XF, Hao XL, Jiang HM, et al. Inhibition of microRNA-543 alleviates sepsis-induced acute kidney injury via targeting Bcl-2. Eur Rev Med Pharmacol Sci. 2022;26(7):2305–12.
26. Peng Y, Li Y, Li H, Yu J. Shikonin attenuates kidney tubular epithelial cells apoptosis, oxidative stress, and inflammatory response through nicotinamide adenine dinucleotide phosphate oxidase 4/PTEN pathway in acute kidney injury of sepsis model. Drug Dev Res. 2022. 10.1002/ddr.21936
27. Zhang Y, Song C, Ni W, Pei Q, Wang C, Ying Y, et al. HSP70 ameliorates septic acute kidney injury via binding with TRAF6 to inhibit of inflammation-mediated apoptosis. J Inflamm Res. 2022;15:2213–28. 10.2147/JIR.S352717
28. Wang B, Xu J, Ren Q, Cheng L, Guo F, Liang Y, et al. Fatty acid-binding protein 4 is a therapeutic target for septic acute kidney injury by regulating inflammatory response and cell apoptosis. Cell Death Dis. 2022;13(4):333. 10.1038/s41419-022-04794-w
29. Nadeem A, Siddiqui N, Al-Harbi NO, Attia SM, AlSharari SD, Ahmad SF. Acute lung injury leads to depression-like symptoms through upregulation of neutrophilic and neuronal NADPH oxidase signaling in a murine model. Int Immunopharmacol. 2017;47:218–26. 10.1016/j.intimp.2017.04.010
30. Nadeem A, Al-Harbi NO, Ahmad SF, Ibrahim KE, Siddiqui N, Al-Harbi MM. Glucose-6-phosphate dehydrogenase inhibition attenuates acute lung injury through reduction in NADPH oxidase-derived reactive oxygen species. Clin Exp Immunol. 2018;191(3):279–87. 10.1111/cei.13097
31. Nadeem A, Ahmad SF, Al-Harbi NO, Al-Harbi MM, Ibrahim KE, Kundu S, et al. Inhibition of spleen tyrosine kinase signaling protects against acute lung injury through blockade of NADPH oxidase and IL-17A in neutrophils and γδ T cells respectively in mice. Int Immunopharmacol. 2019;68:39–47. 10.1016/j.intimp.2018.12.062
32. Nadeem A, Al-Harbi NO, Ahmad SF, Al-Harbi MM, Alhamed AS, Alfardan AS, et al. Blockade of interleukin-2-inducible T-cell kinase signaling attenuates acute lung injury in mice through adjustment of pulmonary Th17/Treg immune responses and reduction of oxidative stress. Int Immunopharmacol. 2020;83:106369. 10.1016/j.intimp.2020.106369
33. Nadeem A, Al-Harbi NO, Alfardan AS, Ahmad SF, AlAsmari AF, Al-Harbi MM. IL-17A-induced neutrophilic airway inflammation is mediated by oxidant-antioxidant imbalance and inflammatory cytokines in mice. Biomed Pharmacother Biomed Pharmacother. 2018;107:1196–204. 10.1016/j.biopha.2018.08.123
34. Nadeem A, Ahmad SF, Al-Harbi NO, Ibrahim KE, Alqahtani F, Alanazi WA, et al. Bruton's tyrosine kinase inhibition attenuates oxidative stress in systemic immune cells and renal compartment during sepsis-induced acute kidney injury in mice. Int Immunopharmacol. 2021;90:107123. 10.1016/j.intimp.2020.107123
35. Nadeem A, Ahmad SF, Al-Harbi NO, Ibrahim KE, Sarawi W, Attia SM, et al. Role of ITK signaling in acute kidney injury in mice: Amelioration of acute kidney injury associated clinical parameters and attenuation of inflammatory transcription factor signaling in CD4+ T cells by ITK inhibition. Int Immunopharmacol. 2021;99:108028. 10.1016/j.intimp.2021.108028
36. Al-Harbi NO, Nadeem A, Ahmad SF, Alanazi MM, Aldossari AA, Alasmari F. Amelioration of sepsis-induced acute kidney injury through inhibition of inflammatory cytokines and oxidative stress in dendritic cells and neutrophils respectively in mice: Role of spleen tyrosine kinase signaling. Biochimie. 2019;158:102–10. 10.1016/j.biochi.2018.12.014
37. Xu F, Xie Q, Li YW, Jing QQ, Liu XJ, Xu YC, et al. Suppression of JNK/ERK dependent autophagy enhances Jaspine B derivative-induced gastric cancer cell death via attenuation of p62/Keap1/Nrf2 pathways. Toxicol Appl Pharmacol. 2022;438:115908. 10.1016/j.taap.2022.115908
38. Jayakumar T, Hou SM, Chang CC, Fong TH, Hsia CW, Chen YJ, et al. Columbianadin dampens in vitro inflammatory actions and inhibits liver injury via inhibition of NF-κB/MAPKs: Impacts on (∙)OH radicals and HO-1 expression. Antioxidants (Basel, Switzerland). 2021;10(4):553. 10.3390/antiox10040553
39. Jin H, Wang M, Wang J, Cao H, Niu W, Du L. Paeonol attenuates isoflurane anesthesia-induced hippocampal neurotoxicity via modulation of JNK/ERK/P38MAPK pathway and regulates histone acetylation in neonatal rat. J Matern Fetal Neonatal Med. 2020;33(1):81–91. 10.1080/14767058.2018.1487396
40. Fang Q, Wang J, Zhang Y, Wang L, Li W, Han J, et al. Inhibition of myeloid differentiation factor-2 attenuates obesity--induced cardiomyopathy and fibrosis. Biochim Biophys Acta Mol Basis Dis. 2018;1864(1):252–62. 10.1016/j.bbadis.2017.09.026
41. Kim KM, Kim SY, Mony TJ, Bae HJ, Han SD, Lee ES, et al. Dracocephalum moldavica ethanol extract suppresses LPS-induced inflammatory responses through inhibition of the JNK/ERK/NF-κB signaling pathway and IL-6 production in RAW 264.7 macrophages and in endotoxic-treated mice.Nutrients. 2021;13(12):4501. 10.3390/nu13124501
42. Cohen TS, Gray Lawrence G, Margulies SS. Cultured alveolar epithelial cells from septic rats mimic in vivo septic lung. PloS One. 2010;5(6):e11322. 10.1371/journal.pone.0011322
43. Li P, Chen XR, Xu F, Liu C, Li C, Liu H, et al. Alamandine attenuates sepsis-associated cardiac dysfunction via inhibiting MAPKs signaling pathways. Life Sci. 2018;206:106–16. 10.1016/j.lfs.2018.04.010
44. Pizzino G, Bitto A, Pallio G, Irrera N, Galfo F, Interdonato M, et al. Blockade of the JNK signalling as a rational therapeutic approach to modulate the early and late steps of the inflammatory cascade in polymicrobial sepsis. Mediat Inflamm. 2015;2015:591572. 10.1155/2015/591572