
Download
ORIGINAL ARTICLE
Cornus iridoid glycoside alleviates sepsis-induced acute lung injury by regulating NF-κB and Nrf2/HO-1 pathways
Xuhui Tanga, Haimin Tangb*
aDepartment of Emergency, Jiangxi Provincial People’s Hospital, Nanchang, Jiangxi Province, China
bDepartment of Pathology, Zhejiang Hospital, Hangzhou, Zhejiang Province, China
Abstract
Background Sepsis-induced acute lung injury (ALI) is a syndrome associated with inflammation. Cornus iridoid glycoside (CIG), a bioactive component isolated from Corni Fructus, exhibits anti-inflammatory activities. However, the function and underlying mechanisms of CIG in mice with sepsis-induced ALI remain elusive.
Methods The sepsis-elicited ALI model of mice was established by the induction of cecal ligation and puncture (CLP). The wet/dry (W/D) ratio of lung tissues was examined, and the pathological alterations were determined by hematoxylin and eosin staining. The messenger RNA (mRNA) expressions and serum levels of Interleukin (IL)-1β, IL-6, and tumor necrosis factor-α (TNF-α) were measured by reverse transcription-quantitative polymerase chain reaction and enzyme-linked immunosorbent serologic assay, respectively. The concentrations of malondialdehyde (MDA), superoxide dismutase (SOD), and glutathione peroxidase (GSH-Px) were assessed by biochemical kits. In addition, the relative protein levels of p-p65, p65, phosphorylated- nuclear factor of kappa light polypeptide gene enhancer in B-cells inhibitor, alpha (p-IκBα), IκBα, nuclear factor erythroid 2-related factor 2 (Nrf2), and heme oxygenase-1 (HO-1) gene were analyzed by Western blotting analysis.
Results CLP enhanced W/D ratio and aggravated pathological changes and scores in mice, which were obviously alleviated by the two concentrations of CIG treatment. CIG treatment notably decreased the CLP-induced mRNA expressions and serum levels of IL-1β, IL-6, TNF-α, and MDA, but enhanced the decreased concentrations (caused by CLP) of SOD and GSH-Px. Moreover, CIG treatment significantly decreased the ratios of p65/p-p65 and IκBα/p-IκBα caused by CLP, but aggravated the CLP-induced relative protein levels of Nrf2 and HO-1.
Conclusions CIG obviously ameliorated the sepsis-induced ALI in mice by suppressing inflammation and oxidative stress, which was closely associated with nuclear factor kappa B (NF-κB) and Nrf2–HO-1 signaling pathways.
Key words: acute lung injury, Cornus iridoid glycoside, NF-κB, Nrf2/HO-1, sepsis
*Corresponding author: Haimin Tang, Department of Pathology, Zhejiang Hospital, No. 12 Lingyin Road, Hangzhou City, Zhejiang Province310030, China. Email address: [email protected]
Received 28 March 2022; Accepted 15 April 2022; Available online 1 September 2022
Copyright: Tang X and Tang H
This open access article is licensed under Creative Commons Attribution 4.0 International (CC BY 4.0). http://creativecommons.org/licenses/by/4.0/
Introduction
Sepsis is a syndrome of systemic inflammatory response caused by serious infection that generally results in multiple organ failure,1 which causes prominent morbidity and mortality.2 The lung is the first and most susceptible organ to sepsis.3 It has been demonstrated that over 50% patients with sepsis progress into acute lung injury (ALI) or acute respiratory distress syndrome (ARDS).4 Moreover, patients with ALI exhibit an approximate mortality of 45%, and a poor prognosis.5 ALI is characterized by pulmonary edema, which is caused by massive infiltration of macrophages and neutrophils into the alveolar epithelium and production of abundant inflammatory mediators, eventually causing injury to lung endothelial cells and epithelial cells.6 Thus, inflammatory response plays a remarkable role in sepsis-induced ALI.7 A variety of therapeutic strategies have been used for the treatment of sepsis-induced ALI in clinical setting, such as hemoadsorption8 and unfracioned heparin;9 however, the outcomes of the stated treatments remain unsatisfactory. Therefore, deep understanding of the potential molecular mechanisms, and seeking effective drugs for sepsis-induced ALI are very imperative for the clinical development of sepsis-induced ALI.
Corni Fructus, the fruit of Cornus officinalis Sieb. et Zucc (family Cornaceae) is generally expended as a nutritional food in the form of drinks and wine. It has been demonstrated that Corni Fructus has anti-inflammatory,10 antioxidative,11 antidiabetic,12 immune regulatory,13 antiatherosclerotic,14 and antimicrobial15 activities. Cornus iridoid glycoside (CIG) is a bioactive extract isolated from Corni Fructus; its beneficial role in focal cerebral ischemia has been verified to be closely involved in anti-inflammatory and anti-apoptotic activities.16 Thus, it is speculated that CIG may have a protective effect against sepsis-induced ALI.
Nuclear factor kappa B (NF-κB), a type of transcription factor, is widely expressed in the cytoplasm, participates in inflammatory and immune responses, and regulates the processes of cell growth.17 NF-κB is silenced and binds to IkappaB kinase (IκB) in the cytoplasm under physiological conditions. Upon stimulation, IκB is first phosphorylated and then degraded, which allows shifting of NF-κB into the nucleus, eventually modulating the level of extensive cytokines associated with inflammatory response.18 Nuclear factor erythroid 2-related factor 2 (Nrf2)/heme oxygenase-1 (HO-1) signaling pathway has been indicated to be involved in oxidative damage, including lipid peroxidation and tissue damage.19 Moreover, it has been revealed that morroniside, a main iridoid glycoside isolated from Corni Fructus, suppresses the inflammation and oxidative stress in lipopolysaccharide-treated RAW 264.7 macrophages through regulation of toll-like receptor 4 (TLR4)/NF-κB and Nrf2/HO-1 signaling pathways.20 However, whether CIG regulates ALI through NF-κB and Nrf2/HO-1 signaling pathways remains unclear.
Therefore, the effect and potential mechanism of CIG on mice with sepsis-induced ALI was investigated. The present study provides a theoretical foundation for the clinical development of ALI therapy.
Materials and methods
Animals
Adult male Sprague-Dawley mice (12 weeks, wt. 410–440 g) were obtained from Shanghai Experimental Animal Center (Shanghai, China) and adapted to breeding environment for 1 week. Mice were fed with standard diet and water ad libitum at 25°C ± 2°C with 40–60% humidity and a 12-h light–dark cycle. All procedures were conducted sternly based on the Guide for the Care and Use of Laboratory Animals,21 and following ethical standards approved by the Ethical Committee of Jiangxi Provincial People’s Hospital.
Establishing ALI rat model and groups
Mice were partitioned randomly into the following four groups: sham, CLP, CLP+15-mg/kg CIG, and CLP+30-mg/kg CIG. ALI rat model was established by the induction of cecal ligation and puncture (CLP) as described by Liang et al.22 In brief, mice were anesthetized intraperitoneally by 0.3% pentobarbital sodium (0.1 mL/10 g); then a 2-cm midline incision was generated in the abdomen. Next, the cecum was exposed and jointed with 4.0 silk thread. Subsequently, the cecum was punctured with a 21-gauge needle and a droplet of feces was extruded via the puncture. Finally, the cecum was sent back to the abdominal cavity, and the peritoneum and skins were seamed in turn. The sham mice underwent identical procedures apart from CLP surgery. Mice in the CLP+15-mg/kg CIG and CLP+30-mg/kg CIG groups were administered intragastrically with 15- and 30-mg/kg CIG (66.59% purity; Chengdu Herbpurify Co. Ltd., Chengdu, China), respectively. Mice in all the groups were offered intragastrically the same amount of normal saline. Mice were administrated intraperitoneally with 0.3% pentobarbital sodium (0.1 mL/10 g) at 24 h after CLP. Blood and lung tissues were collected and assigned for subsequent assays.
Examination of wet/dry (W/D) ratio
The lung tissues were separated and weighed immediately. Then wet lung tissues were dried at 60°C for 24 h. The W/D ratio was quantified to evaluate pulmonary edema.
Hematoxylin and eosin (H&E) staining and score
The lung tissues were segregated and fixed in 4% para-formaldehyde, and subjected to dehydration and embedding. Subsequently, these tissues were sliced sequentially (5 µm) and stained with H&E. The stained slices were analyzed using a digital trinocular camera microscope (CX23, Olympus, Tokyo, Japan). The degree of pathology was scored as follows23: 0 indicated no lesions, 1 indicated damage to an area ≤25%, 2 indicated damage to 26–50% area, 3 indicated damage to 51–70% area, 4 indicated damage to 71–90% area, and 5 indicated damage to >90% area.
Reverse transcription-quantitative polymerase chain reaction (RT-qPCR)
Total RNA of serum samples was extracted by TRIzol reagent (TaKaRa Biotechnology Co. Ltd., Dalian, China) and reverse transcription was carried out using Bio-Rad ScripTM cDNA synthesis kit (Bio-Rad Laboratories Inc., Hercules, CA, USA) according to the operating instructions. RT-qPCR was performed by the Bio-Rad CFX Manager software (Bio-Rad Laboratories Inc.). The following primer sequences were obtained from Sangon Biotech (Shanghai, China):
Interleukin (IL)-1β (Forward primer: 5'-ATCCTCTCCAGTCA GGCTTCCTTGTG-3', Reverse primer: 5'-AGCTCTTGTCGAG ATGCTGTGA-3'); IL-6 (Forward primer: 5'-TGGAGCCCACCAAGAA CGATAGTCAA-3', Reverse primer: 5'TGTCACCAGCATCAGTCCC AAGAAGG-3'); tumor necrosis factor-α (TNF-α) (Forward primer: 5'-AGGACAGCAGAGGACCAGCTAAGAGG-3', Reverse primer: 5'-TGCCACGATCAGGAAGGAGAAGAGG-3'); and glyceraldehyde 3-phosphate dehydrogenase (GAPDH) (Forward primer: 5'-ACGGCAAGTTCAACGGCACAGTCA-3', Reverse primer: 5'-CCA CGACATACTCAGCACCAGCATCA-3'). The RT-qPCR specifies were indicated in the following: 94°C for 5 min, followed by 40 cycles including 15 s at 94°C, 58°C for 30 s, and 30 s at 72°C. The relative expression level of genes was determined by the 2-ΔΔCT method and normalized to GAPDH.24
Enzyme linked immunosorbent serological assay (ELISA)
Blood was stored at room temperature for about 30 min and then centrifuged for 10 min at 3500 rpm to harvest the serum. The concentration of TNF-α, IL-1β, and IL-6 was detected by using ELISA kits (IL-1β [PI301], IL-6 [PI326], and TNF-α [PT512]; Beyotime, Shanghai, China) following the operating instructions.
Biochemical examination
The concentration of malondialdehyde (MDA), superoxide dismutase (SOD), and glutathione peroxidase (GSH-Px) was assessed by commercial MDA test kit (A003-1-1), total superoxide dismutase (T-SOD) test kit (A001-1-1), and Glutathione Peroxidase (GSH-Px) test kit (A005-1-2; all from Nanjing Jiancheng Bioengineering Institute, Nanjing, China) based on the operating instructions. The absorbance of wells was respectively analyzed at 532 nm (MDA), 560 nm (SOD), and 412 nm (GSH) by a microplate reader (Thermo Fisher Scientific, Waltham, MA, USA).
Western blotting analysis
The western blotting assay was performed as described by Li et al.25 In brief, total amount of protein was obtained from the lung tissues using a total protein extraction kit (BC3711; Solarbio, Beijing, China), and the concentration of protein was determined by the BCA protein quantification kit (ab102536; Abcam, Cambridge, UK) following the operation manual. Protein samples were isolated by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE), and then electrically shifted to a polyvinylidene fluoride (PVDF) membrane for 4 h at 150 V. Following pre-blocking with tris buffered saline with tween (TBST; Sigma, St. Louis, MO, USA), including 3% bovine serum albumin (BSA; Sangon Biotech), at room temperature for 1 h, the membrane was hatched overnight at 4°C with the following designated primary antibodies: Rabbit anti-p65, 1:1000, 4764; rabbit anti-phosphorylated p65 [p-p65], 1:1000, 3031; rabbit anti-nuclear factor of kappa light polypeptide gene enhancer in B-cells inhibitor, alpha (IκBα), 1:1000, 9242; rabbit anti-phosphorylated (p)-IκBα, 1:1000, 2859; rabbit anti-Nrf2, 1:1000, 33649; rabbit anti-HO-1, 1:1000, 70081; and rabbit anti-GAPDH, 1:1000, 2118. The membrane was washed with TBS, and then hatched with appropriate secondary antibodies at 37°C for 60 min. Imaging of the bands was conducted by 3,3'-Diaminobenzidine (DAB) staining (Sigma) for 10 min and terminated by washing with distilled water; the gray value was determined by the Quantity One software.
Statistical analysis
The evaluation of data normality was conducted using the Shapiro–Wilk test. Data were detected by one-way analysis of variance (ANOVA) among multiple groups by the SPSS 22.0 package (SPSS Inc., Chicago, IL, USA) followed by post hoc Bonferroni test. Least significant difference-t (LSD-t) test was used in case the variance was homogeneous, while Tamhane’s T2 test was applied if the variance was unequal. Nonparametric tests were employed to compare the data that did not conform to a normal distribution between groups. All results were exhibited as mean values ± standard error of mean (SEM), and the differences were taken as statistically significant at P < 0.05.
Results
CIG relieved lung tissue injury in mice with sepsis-induced ALI
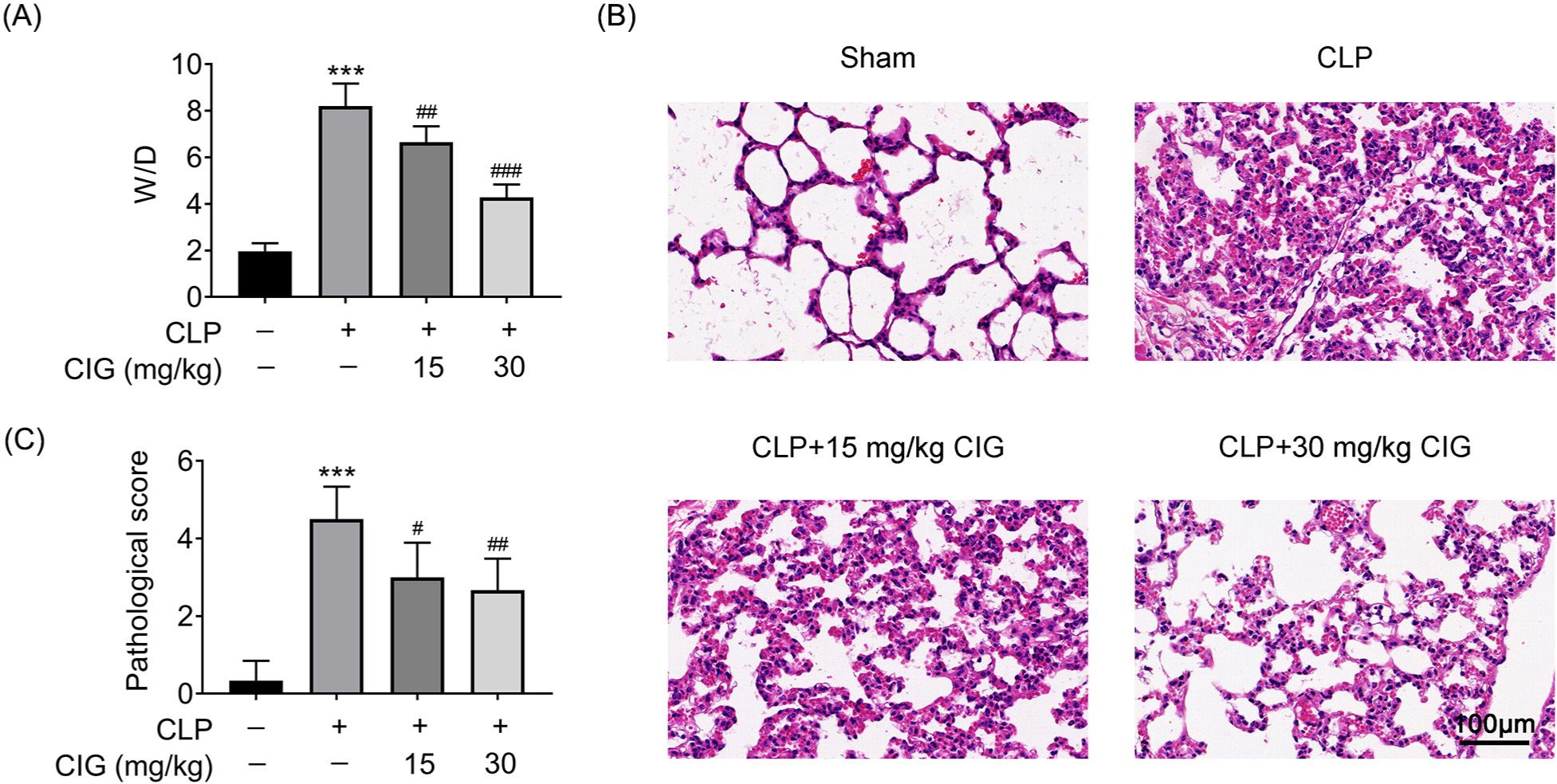
After mice were induced with CLP, the W/D ratio, an indicator of pulmonary edema, increased by nearly four times compared to that in sham mice (P < 0.001; Figure 1A). Moreover, mice treated with CLP exhibited severe damage to alveolar structure, marked as alveolar and interstitial edema, enlarged alveolar spaces, massive inflammatory cell infiltration of alveolar cavity, and capillary congestion in the lungs (Figure 1B). Consistently, the pathological score of mice induced with CLP was significantly higher than that of sham mice (P < 0.001; Figure 1C). Thus, these results suggested that a sepsis-induced ALI rat model was established successfully. However, two different concentrations of CIG prominently alleviated the W/D ratio (P < 0.01; Figure 1A), the pathological manifestation (Figure 1B), and the CLP scores (P < 0.05; Figure 1C). Therefore, these results manifested that CIG ameliorated lung tissue injury in mice with sepsis-induced ALI.
Figure 1 CIG relieved lung tissue injury in mice with sepsis-induced ALI. ALI rat model was established by the induction of cecal ligation and puncture (CLP), and then intragastrically administered 15 mg/kg and 30 mg/kg of CIG, respectively. (A) The W/D ratio was analyzed. (B) The lung tissues were stained with hematoxylin and eosin ×220. (C) The pathological scores were quantified. ***P < 0.001 compared to group without treatment of CLP and CIG, #P < 0.05, ##P < 0.01, and ###P < 0.001 compared to the group with CLP treatment.
CIG inhibited the serum level of inflammatory cytokines in mice with sepsis-induced ALI
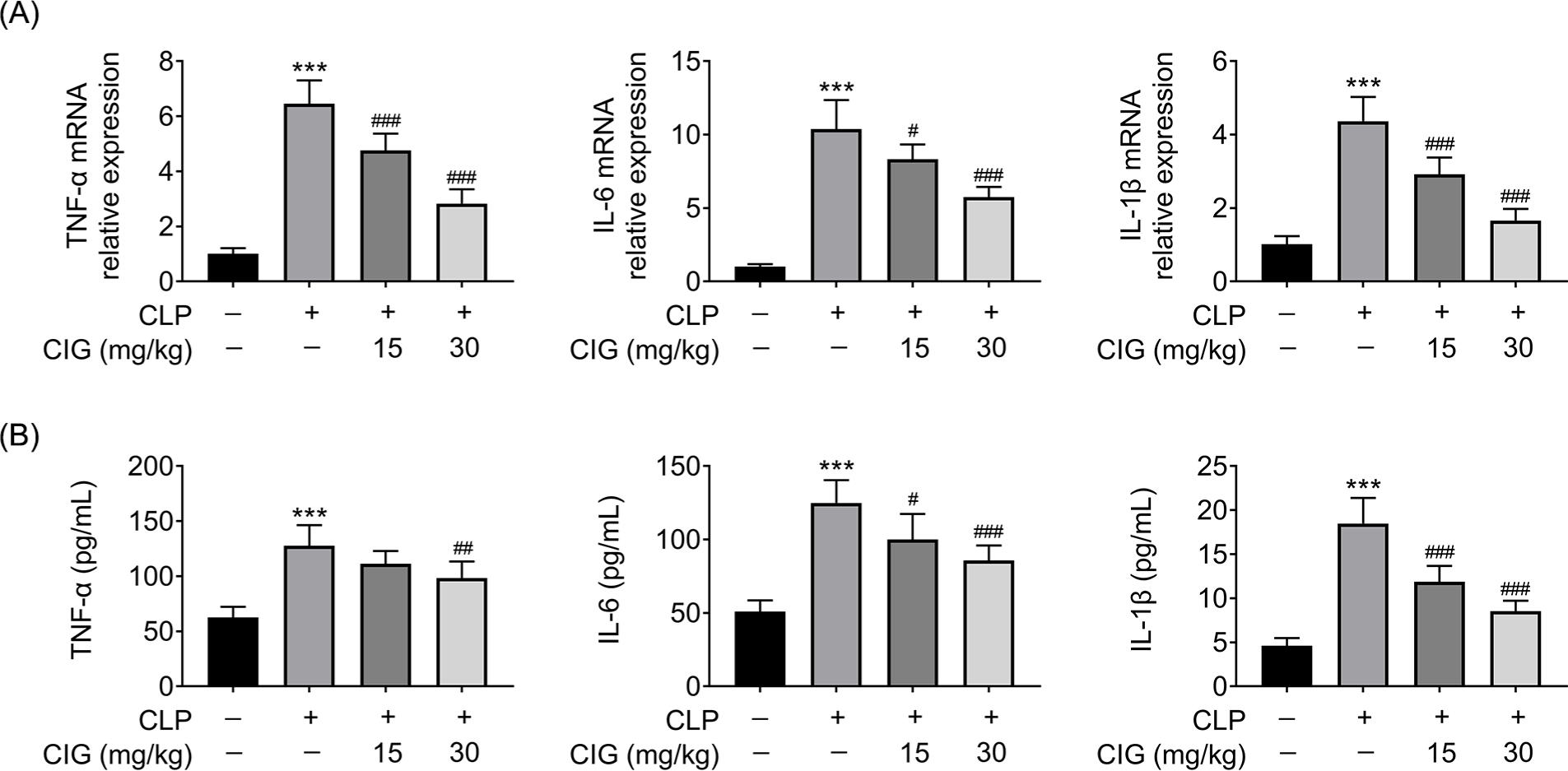
In order to determine the mechanisms inherent to the protection of CIG on lung tissue injury, the serum levels of inflammatory cytokines, including IL-1β, TNF-α, and IL-6, were determined. qRT-PCR results revealed that the relative messenger RNA (mRNA) levels of TNF-α, IL-6, and IL-1β in the serum of ALI mice were observably increased compared to that from sham mice (P < 0.001), which was markedly relieved by treating with 15- or 30-mg/kg CIG (P < 0.05; Figure 2A). Differently, only 30 mg/kg of CIG obviously reduced the CLP-induced serum level of TNF-α (P < 0.01) whereas treatment with 15- or 30-mg/kg CIG notably declined the CLP-induced serum levels of IL-6 and IL-1β (P < 0.05; Figure 2B). Thus, these results elucidated that CIG suppressed the serum level of inflammatory cytokines in mice with sepsis-induced ALI.
Figure 2 CIG repressed the serum level of inflammatory cytokines in mice with sepsis-induced ALI. ALI rat model was established by the induction of cecal ligation and puncture (CLP), and then intragastrically administered 15 mg/kg and 30 mg/kg of CIG, respectively. The blood was collected and serum was harvested via centrifugation. (A) The relative mRNA expressions of TNF-α, IL-6, and IL-1β in serum were examined by qRT-PCR assay. The data were determined after being normalized to GAPDH. (B) The serum levels of TNF-α, IL-6, and IL-1β were measured by ELISA. ***P < 0.001 compared to group without treatment of CLP and CIG, #P < 0.05, ##P < 0.01, and ###P < 0.001 compared to the group with CLP treatment.
CIG attenuated oxidative stress in mice with sepsis-induced ALI
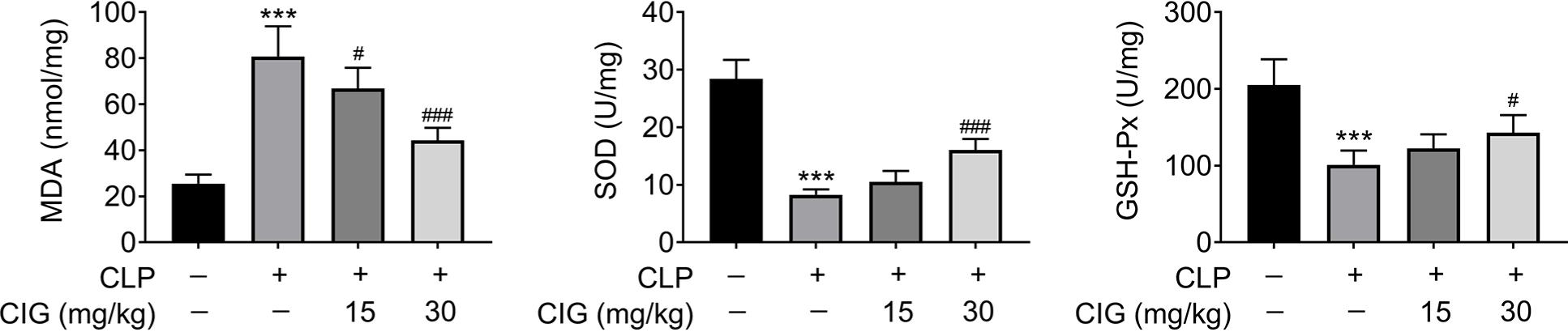
Besides, the level of oxidative stress in the lung tissues of mice with sepsis-induced ALI was also assessed. As shown in Figure 3, the concentration of MDA was prominently enhanced in mice induced with CLP (P < 0.001), which was significantly improved by treatment of 15- or 30-mg/kg CIG (P < 0.05). On the contrary, the concentrations of SOD and GSH-Px were observably declined by administering CLP (P < 0.001), which were notably reversed only by treatment with 30-mg/kg CIG (P < 0.05). Thus, these results indicated that the level of oxidative stress in the lung tissues in CLP-treated mice was obviously suppressed by CIG treatment.
Figure 3 CIG treatment inhibited the oxidative stress of lung tissues in CLP-treated mice. ALI rat model was established by the induction of cecal ligation and puncture (CLP), and then intragastrically administered 15 mg/kg and 30 mg/kg of CIG, respectively. The lung tissues were collected, and the concentration of MDA, SOD, and GSH-Px were examined by commercial kits. ***P < 0.001 compared to the group without treatment of CLP and CIG, #P < 0.05, ##P < 0.01, and ###P < 0.001 compared to the group with CLP treatment.
CIG dampened NF-κB but activated Nrf2/HO-1 signaling pathway in mice with sepsis-induced ALI
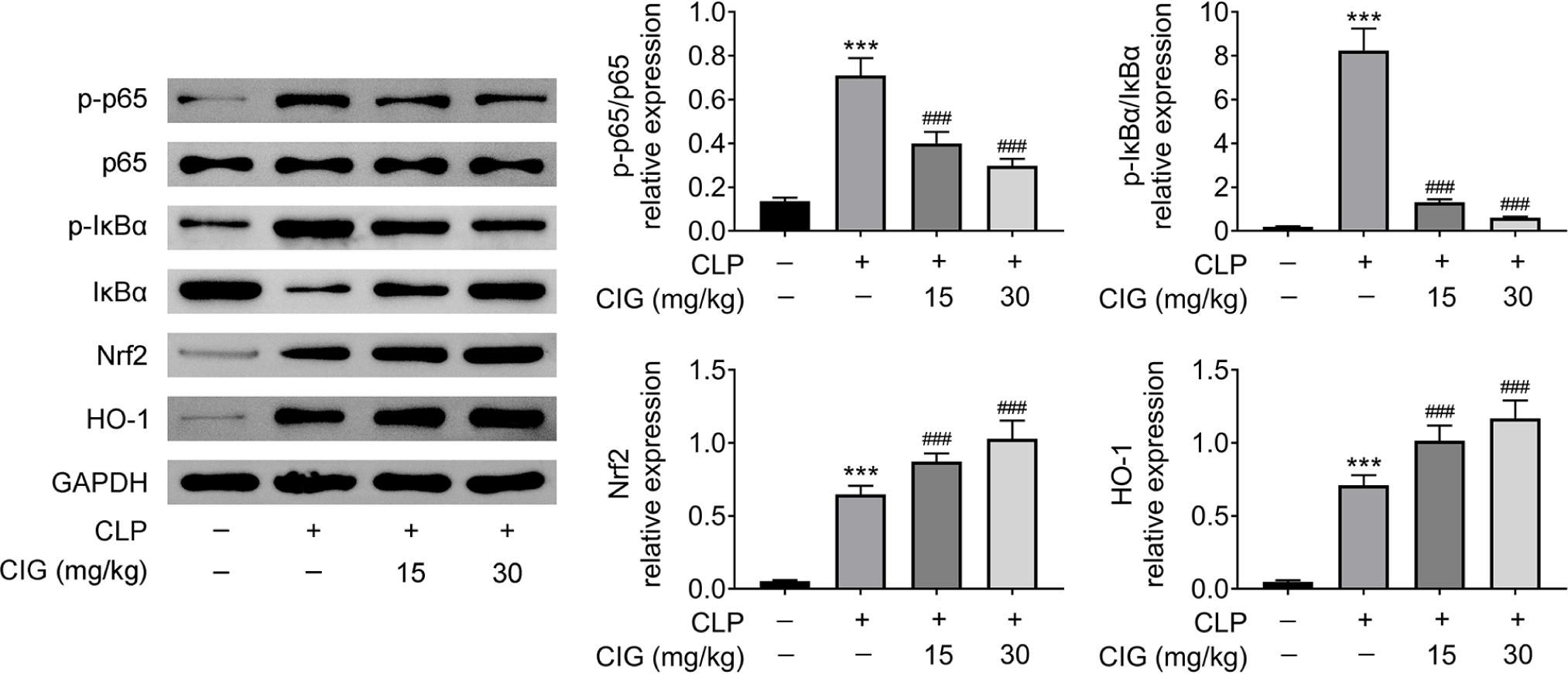
Furthermore, the NF-κB and Nrf2/HO-1 signaling pathways in mice with sepsis-induced ALI were also evaluated by Western blot analysis. The results revealed that treatment with 15- or 30-mg/kg CIG markedly reduced the CLP-induced ratio of p-p65/p65 and p-IκBα/IκBα (P < 0.001; Figure 4), which suggested that CIG treatment notably suppressed NF-κB signaling pathway. However, treatment with 15- or 30-mg/kg CIG memorably further enhanced the CLP-induced relative levels of Nrf2 and HO-1 proteins (P < 0.001; Figure 4), which indicated that CIG treatment obviously promoted Nrf2/HO-1 signaling pathway. There-fore, these results clarified that CIG treatment inhibited NF-κB but activated the Nrf2/HO-1 signaling axis in mice with sepsis-induced ALI.
Figure 4 CIG treatment attenuated NF-κB but prompted Nrf2/HO-1 signaling pathway in mice with sepsis-induced ALI. ALI rat model was established by the induction of cecal ligation and puncture (CLP), and then intragastrically administered 15 mg/kg and 30 mg/kg of CIG, respectively. The lung tissues were collected, and the relative protein levels of p-p65, p65, p-IκBα, IκBα, Nrf2, and HO-1 were determined by Western blot analysis. The data were expressed after being normalized to GAPDH. ***P < 0.001 compared to group without treatment of CLP and CIG, and ###P < 0.001 compared to the group with CLP treatment.
Discussion
It has been demonstrated that sepsis-induced ALI is closely associated with inflammation.26 The anti-inflammatory10 and antioxidative11 functions of Corni Fructus have been reported in the previous studies as well. CIG, a bioactive component of Corni Fructus, has also exhibited anti- inflammatory activities.16 Here, it was discovered that CIG relieved lung tissue injury in mice with sepsis-induced ALI. Mechanically, CIG attenuated inflammation and oxidative stress in mice with sepsis-induced ALI, which might be related to the weakening of NF-κB signaling pathway and the acceleration of Nrf2/HO-1 signaling pathway.
A variety of extracts from traditional Chinese medicine (TCM) have been confirmed to possess protective functions in sepsis-induced ALI, such as steroid ruscogenin extracted from Ophiopogon japonicus (L.f.) Ker-Gawl.,27 artesunate extracted from Artemisia annua L.,28 and norisopoldine (NOR) extracted from Lindera aggregata (Sims) Kosterm.29 Moreover, loganin, one of the best-known iridoid glycosides extracted from Corni Fructus, has been proven to mitigate the sepsis-induced ALI in CLP-induced mice.30 Our results revealed that CIG reduced the CLP-induced W/D ratio, which indicated that CIG improved pulmonary edema. Besides, CIG also relieved lung tissue injury, as evidenced by pathological observation. Therefore, consistent with the previous studies, our results elucidated that CIG ameliorated lung tissue injury in mice with sepsis-induced ALI.
Inflammation is a prominent feature of sepsis-induced ALI.26 Thus, reports have focused on the reduction of inflammation to develop the possible therapy targeting the rat model of sepsis-induced ALI.31 Moreover, the beneficial anti-inflammatory functions of TCM have been demonstrated by Rahmati et al. and Keshvari et al.32,33 In addition to inflammation, oxidative stress also contributed to the development of sepsis-induced ALI.34 Alsharif et al. reported that protocatechuic acid suppressed sepsis-induced ALI in mice via suppressing inflammation and oxidative stress.35 Hu et al. established that flavone isoorientin attenuateed sepsis-induced ALI in mice by the repression of inflammation and oxidative stress.36 The short-term protective effect of digitoxin against sepsis-induced ALI was also associated with inflammation and oxidative stress.37 Moreover, the anti-inflammatory and antioxidative functions of iridoid glycoside from Corni Fructus, including CIG,38 morroniside,39 and loganin,40 have been demonstrated in the type 2 diabetic model. In addition, morroniside also inhibited inflammation and oxidative stress in lipopolysaccharide- administered RAW 264.7 macrophages.20 Furthermore, iridoid glycoside from other natural plants also exhibited the anti-inflammatory and antioxidative activities in different disease models. For instance, aucubin, an iridoid glycoside extracted from natural plants, improved the experimental traumatic brain injury by inhibiting inflammation and oxidative stress.41 Iridoid glycoside isolated from the leaves of Syringa oblata Lindl. relieved inflammation and oxidative stress in ulcerative colitis.42 Here, results demonstrated that CIG treatment notably controlled the CLP-induced mRNA expression levels and serum levels of TNF-α, IL-6, IL-1β, and MDA whereas enhanced the decreased concentrations of SOD and GSH-Px caused by CLP. Thus, our results consistently indicated that CIG attenuated inflammation and oxidative stress in mice with sepsis-induced ALI.
Mechanically, the present study discovered that CIG treatment significantly decreased the CLP-induced ratios of p-p65/p65 and p-IκBα/IκBα whereas further enhanced the decreased relative level of both Nrf2 and HO-1 proteins caused by CLP. Zhao et al. displayed that sulforaphane inhibited sepsis-induced ALI through Nrf2/HO-1 signaling pathway.43 Ni et al. exhibited that nerolidol improved sepsis-induced ALI by attenuating inflammatory response through Nrf-2/HO-1 signaling pathway.19 Moreover, morroniside and loganin could suppress inflammatory response in acute colitis model via NF-κB signaling pathway.44 Morroniside ameliorated diabetic liver complications by inhibiting inflammation and oxidative stress, which was associated with Nrf-2/HO-1 signaling pathway.39 Hence, consistent with the previous studies, our results also established that CIG repressed NF-κB but activated Nrf2/HO-1 signaling axis in sepsis-induced ALI.
Conclusions
The present study elaborated that CIG notably improved the sepsis-induced ALI via inhibiting inflammation and oxidative stress, which was related to NF-κB and Nrf2/HO-1 signaling axis. Although iridoid glycoside from Corni Fructu has been demonstrated to regulate inflammation and oxidative stress through NF-κB and Nrf2/HO-1 signaling axis,39 to our acknowledge, ours were the first evidence elaborating that CIG suppressed inflammation and oxidative stress via NF-κB and Nrf2/HO-1 signaling axis in mice with sepsis-induced ALI. However, there are several limitations to the present study. For instance, other experimental approaches could be applied to validate the effects of CIG on inflammation and oxidative stress. Also, Tthe direct role of NF-κB and Nrf2/HO-1 signaling axis in ALI must be explored in subsequent studies by pharmacological blockade or other effective interference. In brief, our data can offer a theoretical basis for not only treating and prevention of sepsis-induced ALI but also other clinically relevant diseases.
Competing interests
The authors stated that there were no conflicts of interest to disclose.
Data availability
All data supporting the findings of this study are available within the paper, and any raw data can be obtained from the corresponding author upon request.
Author Contributions
Xuhui Tang was involved with conceptualization, methodology, software, and data curation of the manuscipt. Haimin Tang was involved with writing–original draft preparation, visualization, validation, writing and reviewing, and editing of the manuscript. Both authors approved the final manuscript.
REFERENCES
1. Chiu C, Legrand M. Epidemiology of sepsis and septic shock. Curr Opin Anaesthesiol. 2021;34(2):71–6. 10.1097/ACO.0000000000000958
2. Biason L, Teixeira C, Haas JS, Cabral CDR, Friedman G. Effects of sepsis on morbidity and mortality in critically Ill patients 2 years after intensive care unit discharge. Am J Crit Care. 2019;28(6):424–32. 10.4037/ajcc2019638
3. Rubenfeld GD, Caldwell E, Peabody E, Weaver J, Martin DP, Neff M, et al. Incidence and outcomes of acute lung injury. N Engl J Med. 2005;353(16):1685–93. 10.1056/NEJMoa050333
4. Villar J, Sulemanji D, Kacmarek RM. The acute respiratory distress syndrome: Incidence and mortality, has it changed? Curr Opin Crit Care. 2014;20(1):3–9. 10.1097/MCC.0000000000000057
5. Matthay MA, Zemans RL, Zimmerman GA, Arabi YM, Beitler JR, Mercat A, et al. Acute respiratory distress syndrome. Nat Rev Dis Primers. 2019;5(1):18. 10.1038/s41572-019-0069-0
6. Zhang Y, Yu W, Han D, Meng J, Wang H, Cao G. L-lysine ameliorates sepsis-induced acute lung injury in a lipopolysaccharide-induced mouse model. Biomed Pharmacother. 2019;118:109307. 10.1016/j.biopha.2019.109307
7. Ding X, Tong Y, Jin S, Chen Z, Li T, Billiar TR, et al. Mechanical ventilation enhances extrapulmonary sepsis-induced lung injury: role of WISP1-αvβ5 integrin pathway in TLR4-mediated inflammation and injury. Crit Care. 2018;22(1):302. 10.1186/s13054-018-2237-0
8. Sazonov V, Abylkassov R, Tobylbayeva Z, Saparov A, Mironova O, Poddighe D. Case series: Efficacy and safety of hemoadsorption with HA-330 adsorber in septic pediatric patients with cancer. Front Pediatr. 2021;9:672260. 10.3389/fped.2021.672260
9. Fu S, Yu S, Wang L, Ma X, Li X. Unfractionated heparin improves the clinical efficacy in adult sepsis patients: A systematic review and meta-analysis. BMC Anesthesiol. 2022;22(1):28. 10.1186/s12871-021-01545-w
10. Li HB, Feng QM, Zhang LX, Wang J, Chi J, Chen SQ, et al. Four new gallate derivatives from wine-processed corni fructus and their anti-inflammatory activities. Molecules. 2021;26(7):1851. 10.3390/molecules26071851
11. Wang X, Wu Y, Wang F, Yi J, Zhang Y. Optimization of polysaccharide process from Fructus corni with box-behnken design and antioxidant capacity. Pak J Pharm Sci. 2019;32(4):1537–44. PMid: 31608872
12. Bi H, Niu D, Guo C, Li J, Chen X, Zhang Y, et al. Comparative study of crude and wine-processing Corni Fructus on chemical composition and antidiabetic effects. Evid Based Complement Alternat Med. 2019;2019:3986964. 10.1155/2019/3986964
13. Wu Y, Wang X, Shen B, Kang L, Fan E. Extraction, structure and bioactivities of the polysaccharides from Fructus corni. Recent Pat Food Nutr Agric. 2013;5(1):57–61. 10.2174/2212798411305010009
14. Zhang XW, Sui Y, Liu XX, Fu CY, Qiao YH, Liu WJ, et al. Structures and anti-atherosclerotic effects of 1,6-α-glucans from Fructus corni. Int J Biol Macromol. 2020;161:1346–57. 10.1016/j.ijbiomac.2020.08.038
15. Mau J, Chen C, Hsieh P. Antimicrobial effect of extracts from Chinese chive, cinnamon, and corni fructus. J Agric Food Chem. 2001;49(1):183–8. 10.1021/jf000263c
16. Ya BL, Li CY, Zhang L, Wang W, Li L. Cornel iridoid glycoside inhibits inflammation and apoptosis in brains of rats with focal cerebral ischemia. Neurochem Res. 2010;35(5):773–81. 10.1007/s11064-010-0134-2
17. Silverman N, Maniatis T. NF-kappaB signaling pathways in mammalian and insect innate immunity. Genes Dev. 2001;15(18):2321–42. 10.1101/gad.909001
18. Haddad JJ, Abdel-Karim NE. NF-κB cellular and molecular regulatory mechanisms and pathways: Therapeutic pattern or pseudo regulation? Cell Immunol. 2011;271(1):5–14. 10.1016/j.cellimm.2011.06.021
19. Ni YL, Shen HT, Su CH, Chen WY, Huang-Liu R, Chen CJ, et al. Nerolidol suppresses the inflammatory response during lipopolysaccharide-induced acute lung injury via the modulation of antioxidant enzymes and the AMPK/Nrf-2/HO-1 pathway. Oxid Med Cell Longev. 2019;2019:9605980. 10.1155/2019/9605980
20. Park C, Cha HJ, Lee H, Kim GY, Choi YH. The regulation of the TLR4/NF-κB and Nrf2/HO-1 signaling pathways is involved in the inhibition of lipopolysaccharide-induced inflammation and oxidative reactions by morroniside in RAW 264.7 macrophages. Arch Biochem Biophys. 2021;706:108926. 10.1016/j.abb.2021.108926
21. National Research Council (US) Committee for the Update of the Guide for the Care and Use of Laboratory Anumals. Guide for the care and use of laboratory animals. 8th ed. The National Academies collection: Reports funded by National Institutes of Health. Washington, DC: National Academies Press; 2011. PMid: 21595115 Copyright ©2011, National Academy of Sciences.
22. Liang W, Guo L, Liu T, Qin S. MEF2C alleviates acute lung injury in cecal ligation and puncture (CLP)-induced sepsis rats by up-regulating AQP1. Allergol Immunopathol (Madr). 2021;49(5):117–24. 10.15586/aei.v49i5.477
23. Wang ZF, Yang YM, Fan H. Protective effect of S-nitrosoglutathione pretreatment on acute lung injury in septic rats. Iran J Basic Med Sci. 2020;23(8):1059–64. 10.22038/ijbms.2020.43590.10240
24. Chen S, Chen X, Xiu Y-L, Sun K-X, Zong Z-H, Zhao Y. microRNA 490-3P enhances the drug-resistance of human ovarian cancer cells. J Ovarian Res. 2014;7:84. 10.1186/s13048-014-0084-4
25. Li S, Sun Y, Tang X, Wang L, Cheng X. miR-20a attenuates acute lung injury in septic rats via targeting TLR4. Signa Vitae. 2021;17(4):157–62.
26. da-Palma-Cruz M, da Silva RF, Monteiro D, Rehim H, Grabulosa CC, de Oliveira APL, et al. Photobiomodulation modulates the resolution of inflammation during acute lung injury induced by sepsis. Lasers Med Sci. 2019;34(1):191–9. 10.1007/s10103-018-2688-1
27. Wang Y, Xue L, Wu Y, Zhang J, Dai Y, Li F, et al. Ruscogenin attenuates sepsis-induced acute lung injury and pulmonary endothelial barrier dysfunction via TLR4/Src/p120-catenin/VE-cadherin signalling pathway. J Pharm Pharmacol. 2021;73(7):893–900. 10.1093/jpp/rgaa039
28. Zhang E, Wang J, Chen Q, Wang Z, Li D, Jiang N, et al. Artesunate ameliorates sepsis-induced acute lung injury by activating the mTOR/AKT/PI3K axis. Gene. 2020;759:144969. 10.1016/j.gene.2020.144969
29. Chen Q, Shao X, He Y, Lu E, Zhu L, Tang W. Norisoboldine attenuates sepsis-induced acute lung injury by modulating macrophage polarization via PKM2/HIF-1α/PGC-1α pathway. Biol Pharm Bull. 2021;44(10):1536–47. 10.1248/bpb.b21-00457
30. Zhang J, Wang C, Wang H, Li X, Xu J, Yu K. Loganin alleviates sepsis-induced acute lung injury by regulating macrophage polarization and inhibiting NLRP3 inflammasome activation. Int Immunopharmacol. 2021;95:107529. 10.1016/j.intimp.2021.107529
31. Oliveira-Junior IS, Brunialti MK, Koh IH, Junqueira VB, Salomão R. Effect of pentoxifylline on lung inflammation and gas exchange in a sepsis-induced acute lung injury model. Braz J Med Biol Res. 2006;39(11):1455–63. 10.1590/S0100-879X2006001100009
32. Rahmati M, Keshvari M, Mirnasouri R, Chehelcheraghi F. Exercise and Urtica dioica extract ameliorate hippocampal insulin signaling, oxidative stress, neuroinflammation, and cognitive function in STZ-induced diabetic rats. Biomed Pharmacother. 2021;139:111577. 10.1016/j.biopha.2021.111577
33. Keshvari M, Rahmati M, Mirnasouri R, Chehelcheraghi F. Effects of endurance exercise and Urtica dioica on the functional, histological and molecular aspects of the hippocampus in STZ-induced diabetic rats. J Ethnopharmacol. 2020;256:112801. 10.1016/j.jep.2020.112801
34. Guo RF, Ward PA. Role of oxidants in lung injury during sepsis. Antioxid Redox Signal. 2007;9(11):1991–2002. 10.1089/ars.2007.1785
35. Alsharif KF, Almalki AA, Alsanie WF, Alzahrani KJ, Kabrah SM, Elshopakey GE, et al. Protocatechuic acid attenuates lipopolysaccharide-induced septic lung injury in mice: The possible role through suppressing oxidative stress, inflammation and apoptosis. J Food Biochem. 2021;45(10):e13915. 10.1111/jfbc.13915
36. Hu M, Yang J, Xu Y. Isoorientin suppresses sepsis-induced acute lung injury in mice by activating an EPCR-dependent JAK2/STAT3 pathway. J Mol Histol. 2022 Feb;53(1):97–109. 10.1007/s10735-021-10039-5
37. Bora E-S, Erdoğan M-A, Özkul B, Sever İ-H, Söğüt İ, Hürdağ C, et al. Short-term protective effect of digitoxin in sepsis-induced acute lung injury. Biocell. 2022;46(2):433-9. 10.32604/biocell.2022.018510
38. Niu D, Chen X, Wang T, Wang F, Zhang Q, Xue X, et al. Protective effects of iridoid glycoside from corni fructus on type 2 diabetes with nonalcoholic fatty liver in mice. Biomed Res Int. 2021; 2021:3642463. 10.1155/2021/3642463
39. Park CH, Noh JS, Kim JH, Tanaka T, Zhao Q, Matsumoto K, et al. Evaluation of morroniside, iridoid glycoside from corni fructus on diabetes-induced alterations such as oxidative stress, inflammation, and apoptosis in the liver of type 2 diabetic db/db mice. Biol Pharm Bull. 2011;34(10):1559–65. 10.1248/bpb.34.1559
40. Park CH, Tanaka T, Kim JH, Cho EJ, Park JC, Shibahara N, et al. Hepato-protective effects of loganin, iridoid glycoside from Corni Fructus, against hyperglycemia-activated signaling pathway in liver of type 2 diabetic db/db mice. Toxicology. 2011;290(1):14–21. 10.1016/j.tox.2011.08.004
41. Wang H, Zhou XM, Wu LY, Liu GJ, Xu WD, Zhang XS, et al. Aucubin alleviates oxidative stress and inflammation via Nrf2-mediated signaling activity in experimental traumatic brain injury. J Neuroinflamm. 2020;17(1):188. 10.1186/s12974-020-01863-9
42. Zhang Y, Han D, Yu S, An C, Liu X, Zhong H, et al. Protective effect of iridoid glycosides of the leaves of Syringa oblata Lindl. on dextran sulfate sodium-induced ulcerative colitis by inhibition of the TLR2/4/MyD88/NF-κB signaling pathway. Biomed Res Int. 2020;2020:7650123. 10.1155/2020/7650123
43. Zhao B, Gao W, Gao X, Leng Y, Liu M, Hou J, et al. Sulforaphane attenuates acute lung injury by inhibiting oxidative stress via Nrf2/HO-1 pathway in a rat sepsis model. Int J Clin Exp Pathol. 2017;10(8):9021–8. PMid: 31966772
44. Yuan J, Cheng W, Zhang G, Ma Q, Li X, Zhang B, et al. Protective effects of iridoid glycosides on acute colitis via inhibition of the inflammatory response mediated by the STAT3/NF-κB pathway. Int Immunopharmacol. 2020;81:106240. 10.1016/j.intimp.2020.106240