
Download
ORIGINAL ARTICLE
6-Gingerol, a functional polyphenol of ginger, reduces pulmonary fibrosis by activating Sirtuin1
Li Liua, Nan Yub, Wei Lengb, Yun Lua, Xinxin Xiac, Hui Yuanb*
aHospital of Chengdu University of Traditional Chinese Medicine, Chengdu, China
bThe Affiliated Hospital of Shaanxi University of Traditional Chinese Medicine, Xianyang, China
cThe First Affiliated Hospital of Xi’an Jiaotong University, Xi’an, China
Abstract
Pulmonary fibrosis in general is the final common outcome of various interstitial lung diseases. In recent years, the incidence of pulmonary fibrosis has been rising with poor prognosis. 6-gingerol is deemed as a functional polyphenol of ginger. The aim of the present study was to investigate the effect of 6-gingerol, on pulmonary fibrosis. Mice were randomly divided into four groups: control, bleomycin, bleomycin + 6-gingerol 100 mg/kg, bleomycin + 6-gingerol 250 mg/kg, and the survival rates of the groups were recorded. Pathological and fibrotic changes in the lungs were identified by H&E and Masson staining, respectively. The levels of hydroxyproline and protein deposited in lung tissues were then, respectively, determined by colorimetry and western blotting. Subsequently, the proportion of cells and inflammatory factors in the alveolar lavage fluid were estimated. Following the identification of the possibility of Sirtuin1 (SIRT1) in the pharmacological mechanism through molecular docking and western blotting, human embryonic lung fibroblasts MRC-5 were treated with TGF-β1 and SIRT1 inhibitor to study the role of SIRT1 in the regulatory effect of 6-gingerol. From the results, 6-gingerol was found to increase the survival rate of mice and reduce lung pathology and fibrosis in mice. And, it significantly reduced the levels of hydroxyproline and the proteins deposited in lung tissues. Moreover, the number of neutrophils, basophils, monocytes, and the levels of inflammatory factors in the alveolar lavage fluid were also reduced. SIRT1 inhibitor blocked the function of 6-gingerol to inhibit fibrosis. To sum up, 6-gingerol relieves pulmonary fibrosis via activating SIRT1. This finding expands the pharmacological effect of 6-gingerol, and it is expected to advance the development of treatments for pulmonary fibrosis.
Key words: 6-gingerol, SIRT1, pulmonary fibrosis, MRC-5 cells, ginger
*Corresponding author: Yuan H, The Affiliated Hospital of Shaanxi University of Traditional Chinese Medicine, Xianyang, China. Email address: [email protected]
Received 2 November 2021; Accepted 7 December 2021; Available online 1 March 2022
Copyright: Liu L, et al.
License: This open access article is licensed under Creative Commons Attribution 4.0 International (CC BY 4.0). http://creativecommons.org/licenses/by/4.0/
Introduction
The lung is composed of bronchi, alveoli, blood vessels, nerves, and lymph and connective tissue.1 Among them, alveoli and bronchi belong to the lung parenchyma, and the rest are parts of the pulmonary interstitium, which is distributed between the parenchyma and plays a role in connection, fixation, and nutrition.2 Viral infection, air pollution, smoking, and immune system abnormalities are all potential risk factors for interstitial lung diseases.3 Due to the unpredictability of interstitial lung diseases,4 daily attention should be paid to reducing the erosion of pathogenic factors. Moreover, pulmonary fibrosis is often the final common outcome of various interstitial lung diseases.5 In recent years, the incidence of pulmonary fibrosis has been on the rise, and its poor prognosis is manifested by the low average survival time of less than 5 years after diagnosis.6,7 Delaying the progress of pulmonary fibrosis and reducing its incidence are major needs for disease prevention and treatment due to the lack of effective radical treatment.8 It is well documented that the accumulation of inflammatory factors, the collapse of blood vessels, the increase of myofibroblasts, the deposition of extracellular matrix, and the final fibrosis are pivotal factors contributing to the pathogenesis of pulmonary fibrosis.9,10 Therefore, seeking effective treatment targets and lead compounds have great clinical significance for reducing the incidence of pulmonary fibrosis.
Ginger is a kind of rhizome-type spice seasoning that is widely cultivated worldwide. It is extensively applied in food processing and food preparation.11 Notably, people in numerous countries are already accustomed to treating flu, typhoid, and cough by making ginger tea. Contemporary scientific researchers have confirmed that the component gingerol accounts for the pharmacological properties of ginger.12 The gingerol isolated from ginger is a mixture of multiple substances. According to the chemical structure, gingerol is divided into two categories: single aromatic ring type and double aromatic ring heptane type. The main difference between them is the aromatic ring substituent.13 There are currently more than 10 specific gingerols, all of which have similar properties and structures.14 Among them, 6-gingerol, the content of which accounts for more than 80% of the total gingerol, is the most important component.15 Multiple studies have demonstrated that 6-gingerol may be capable of ameliorating hypoglycemia and exerting anti-hyperlipidemia, antioxidant, and anti-inflammatory roles.16–19 Furthermore, 6-gingerol has been found to reduce isoproterenol-induced myocardial fibrosis.20 Therefore, the hypothesis that 6-gingerol could also reduce pulmonary fibrosis was proposed in this work.
In the present study, for the purpose of studying the effect of 6-gingerol on pulmonary fibrosis, mice and MRC-5 cells were separately induced with bleomycin (BLM)21 or TGF-β122 to establish pulmonary fibrosis models. The experimental protocol is shown in Figure 1. The findings of this research were expected to expand the pharmacological effect of 6-gingerol, and advance the development of treatments for pulmonary fibrosis.
Figure 1 The experimental protocol of this study.
Materials and Methods
Mice and in vivo model establishment
All animal experiments were carried out in strict compliance with the National Institutes of Health guidelines for the handling of laboratory animals and approved by the Ethics Committee of our Hospital. A total of 80 male C57BL/6 mice (aged 6–8 weeks; Cyagen Biosciences, Suzhou, China) were adaptively housed in an SPF environment with a 12-h light/dark cycle, relative humidity of 55%, and free access to food and water at 25°C for a week.
The mice were randomly divided into four groups: control, BLM, BLM + 6-gingerol 100 mg/kg, BLM + 6-gingerol 250 mg/kg. BLM group: for the induction of fibrosis, BLM (purity > 98.00%, GlpBio, United States) was dissolved in 50 μL sterile PBS, and mice were intratracheally administered (2 mg/kg body weight) with a single dose of BLM. This induction lasted 21 days, once a day. BLM + 6-gingerol group: in addition to being treated with BLM, mice were given 6-gingerol (100 or 250 mg/kg, purity > 98.00%, Yuanye Biotechnology, Shanghai, China) for 21 days, once a day. The control group was given normal saline gavage. When 21 days or the ethical endpoints were reached, the mice were euthanized using an overdose of sodium pentobarbital (100 mg/kg, intraperitoneal injection) followed by cervical dislocation.
Cell lines and in vitro model establishment
Human embryonic lung fibroblasts MRC-5 were purchased from Procell Life Science & Technology (Wuhan, China) and cultured in DMEM (Gibco, United States) supplemented with 10% fetal bovine serum (FBS, Invitrogen, CA, United States), 1% penicillin/streptomycin (Invitrogen) in 5% CO2 atmosphere at 37°C.
The cells were randomly divided into different groups: control, TGF-β1 (Abmole, Shanghai, China), TGF-β1 + 6-gingerol, TGF-β1 + 6-gingerol + EX-527 [Sirtuin1 (SIRT1) inhibitor, purity > 98.00%, GlpBio, United States]. To induce fibrosis, cells were exposed to 10 ng/mL TGF-β1 for 48 h. Other experimental groups were exposed to 10 ng/mL TGF-β1 and 6-gingerol (10 or 20 μM) or EX-527 (10 μM). Untreated cells were used as the control group.
Hematoxylin-eosin (H&E) staining
H&E staining was performed using H&E staining kit (#C0105, Beyotime Institute of Biotechnology, China) according to the manufacturer’s instructions. After the mice were sacrificed, the left lung tissues were collected and fixed in 4% paraformaldehyde at 4°C for 6 h, dehydrated with gradient alcohol, embedded in paraffin, and cut into 4-µm slices. The sections were deparaffinized in xylene, and rehydrated in gradient alcohols followed by being stained with hematoxylin solution for 5 min at room temperature. After the sections were washed with distilled water and stained with eosin for 30 s at room temperature, they were dehydrated further with gradient alcohol and became transparent using xylene for 5 min. Physiological changes of the samples were observed under a light microscope (magnification 400×, Leica, Germany).
Masson staining
This assay was performed using Masson staining kit (#DC0032, Leagene, Anhui, China) according to the manufacturer’s instructions. Tissue sections were obtained as aforementioned. Following the staining of sections with Weigert’s iron hematoxylin for 5 min, they were differentiated with acidic ethanol for 10 s. Then, sections were treated with the aniline blue solution for 1 min, followed by ponceau red magenta dyeing solution for 5 min. Subsequently, sections were washed with the phosphomolybdic acid solution for 1 min and counterstained with the aniline blue dyeing solution for 1 min, the results were observed after dehydration under a light microscope (magnification 400×, Leica, Germany).
Determination of hydroxyproline concentration
This assay was performed using an assay kit (#A030-2-1, Jiancheng Bioengineering Institute, Nanjing, China) according to the manufacturer’s instructions. Lung tissue (50 mg) was taken in a tube, and 1 mL of hydrolysis solution was added to the tube. The tube was placed in boiling water for 20 min, and the PH value of the solution was adjusted to 6.0–6.8. After the sample was bathed in 60°C water for 15 min, centrifuged at 3500 r/min for 10 min, the supernatant was collected to measure the absorbance at 550 nm using a microplate reader (Dojindo, Rockville, United States).
Enzyme-linked immunosorbent (ELISA) assay
The necks of mice were dissected and the tracheae were exposed, lavage was performed with 0.8 mL PBS for three times, and the alveolar lavage fluid was collected as described previously.23 The collected lavage fluid was centrifuged at 3500 r/min and 4°C for 10 min. The expression levels of TNF-α (#88-7324) and IL-1β (#88-7013; both Thermo Fisher Scientific, Shanghai, China) in the supernatant were determined using commercial kits according to the manufacturer’s instructions. The absorbance at 450 nm was measured using a microplate reader (Dojindo).
Western blotting analysis
Cultured cells and lung tissue samples from experimental mice were lysed in RIPA lysis buffer (Beyotime Institute of Biotechnology, China). After the determination of protein concentration with a BCA protein assay kit (#A045-3-2, Nanjing JianCheng Bioengineering Institute, Nanjing, China), 20 µg of protein per lane was separated by 10% SDS-PAGE. The separated proteins were transferred onto PVDF membranes, and the membranes were blocked with 5% skimmed milk at room temperature for 2 h. Subsequently, they were continued to incubate with primary antibodies against α-SMA (#ab5694, 1:200), collagen IV (#ab6586, 1:1000), SIRT1 (#ab189494, 1:1000), and GAPDH (#ab9485, 1:2500) at 4°C overnight, followed by HRP-conjugated anti-rabbit antibody (#ab97051; 1: 10,000) at 37°C for 1.5 h. All antibodies were acquired from Abcam. The membranes were developed with an ECL reagent (Millipore) at room temperature for 2 min. The blots were quantified using ImageJ software (v1.8; National Institutes of Health).
Reverse transcription-quantitative PCR (RT-qPCR)
Total RNA was extracted from cultured cells using TRIzol® reagent (Invitrogen). The extracted RNA was reverse transcribed into cDNA using a PrimeScript RT reagent kit (Takara, Japan) according to the manufacturer’s protocol. The mRNA levels were quantified using a QuantiTect SYBR Green PCR Kit (Qiagen) with the ABI7500 system (Applied Biosystems). The PCR amplification conditions were as follows: 95°C for 10 min and then 40 cycles of 95°C for 30 s, 64°C for 34 s, and 72°C for 30 s. The relative mRNA level was normalized to GAPDH and calculated using the 2−ΔΔCq method. The used primers for PCR are listed in Table 1.
Table 1 Primer sequences used for real-time PCR analysis.
| Gene | Sequence (5’ → 3’) |
|---|---|
| α-SMA | F: TCCGCTTCAATTCCTGTCCG |
| R: CAGGATTCCCGTCTTAGTCCC | |
| collagen IV | F:CCGGATCACATTGACATGAAACC |
| R:TGGAAACCAGTCCATGCTCG | |
| TNF-α | F: AGAACTCACTGGGGCCTACA |
| R: GCTCCGTGTCTCAAGGAAGT | |
| IL-6 | F: CTTCGGTCCAGTTGCCTTCTC |
| R: GGCATTTGTGGTTGGGTCAG | |
| IL-1β | F: AGCCATGGCAGAAGTACCTG |
| R: TGAAGCCCTTGCTGTAGTGG | |
| GAPDH | F: GACTCATGACCACAGTCCATGC |
| R: AGAGGCAGGGATGATGTTCTG |
Cell counting kit-8 (CCK-8) assay
Cell viability was determined using a CCK-8 assay. Cells (1 × 104 cells/well) were seeded in 96-well plates. After cultivation for 24 h, the normal medium was replaced with DMEM medium containing TGF-β1 (10 ng/mL), 6-gingerol (10 or 20 μM), or EX-527 (10 μM). Cells were continued to be incubated in a humidified incubator at 37°C for 24, 48, or 72 h. Afterwards, 10 μL of CCK-8 solution was added to each well for a further incubation for 2 h at 37°C. The optical density values were recorded at 450 nm using a microplate reader (Dojindo).
Molecular docking
Molecular docking was used to evaluate the binding force between 6-gingerol and SIRT1 protein. AutoDock v4.2 software (Scripps Institute) was used in this study to perform molecular docking simulation. The three-dimensional (3D) structure of 6-gingerol was imported into the software and adjusted to display hydrogen atoms. The 3D structure of the protein was downloaded from the Protein Data Bank (PDB) website (https://www.rcsb.org/) and imported into the software. The water molecules and excess residues or ligands in the protein file were removed. After setting the docking position, the molecular docking was operated, and the docking energy value of the two was displayed after the operation was over.
Statistical analysis
Data were presented as means ± standard deviations (SD) of three experiments. One-way ANOVA with Tukey’s post-hoc test was used to compare differences among multiple groups. Statistical analysis was performed using GraphPad Prism 8.0 software, and the value of p < 0.05 was considered statistically significant.
Results
6-Gingerol improves the survival rate of bleomycin-induced mice and reduces fibrosis
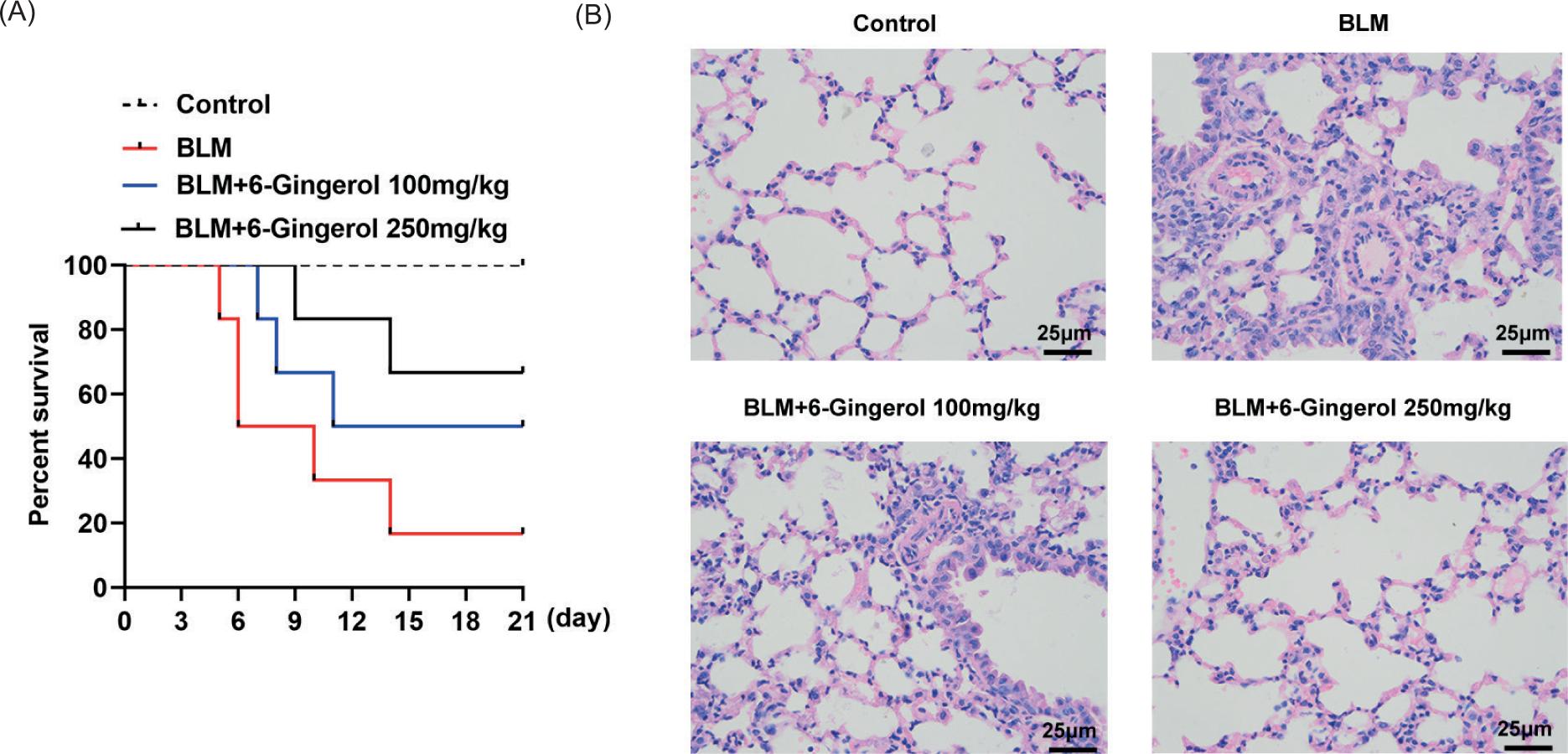
The number of surviving mice in the four groups was recorded every day for a total of 21 days, and the survival rate was exhibited in the form of a line graph. The red line represented the survival rate of the BLM group, and blue and black lines represented the 6-gingerol treatment group at different concentrations. The daily survival rate of the 6-gingerol group was higher than that of the BLM group, and the high-concentration group was more effective in improving the survival of mice (Figure 2A). And, the changes of lung pathology in each group were observed by H&E staining. In the control group, the alveolar morphology was normal, the wall was slender, and there was no inflammatory cell infiltration in the lung interstitium. However, obvious pulmonary fibrosis changes were seen in the BLM group, manifested as sheet consolidation, with much inflammatory cell infiltration, fibroblast proliferation, and collagen deposition in the consolidation area. Simultaneously, less consolidation and fewer inflammatory cells were observed in the 6-gingerol 100 mg/kg treatment group than in the BLM group. More importantly, in the 6-gingerol 250 mg/kg treatment group, there was no obvious fibrosis and only a weak inflammatory cell infiltration (Figure 2B).
Figure 2 6-Gingerol improves the survival rate of bleomycin-induced mice and reduces lung pathology. (A) The survival rate of different groups of mice is exhibited in a line graph. (B) The changes of lung pathology in each group were observed by H&E staining.
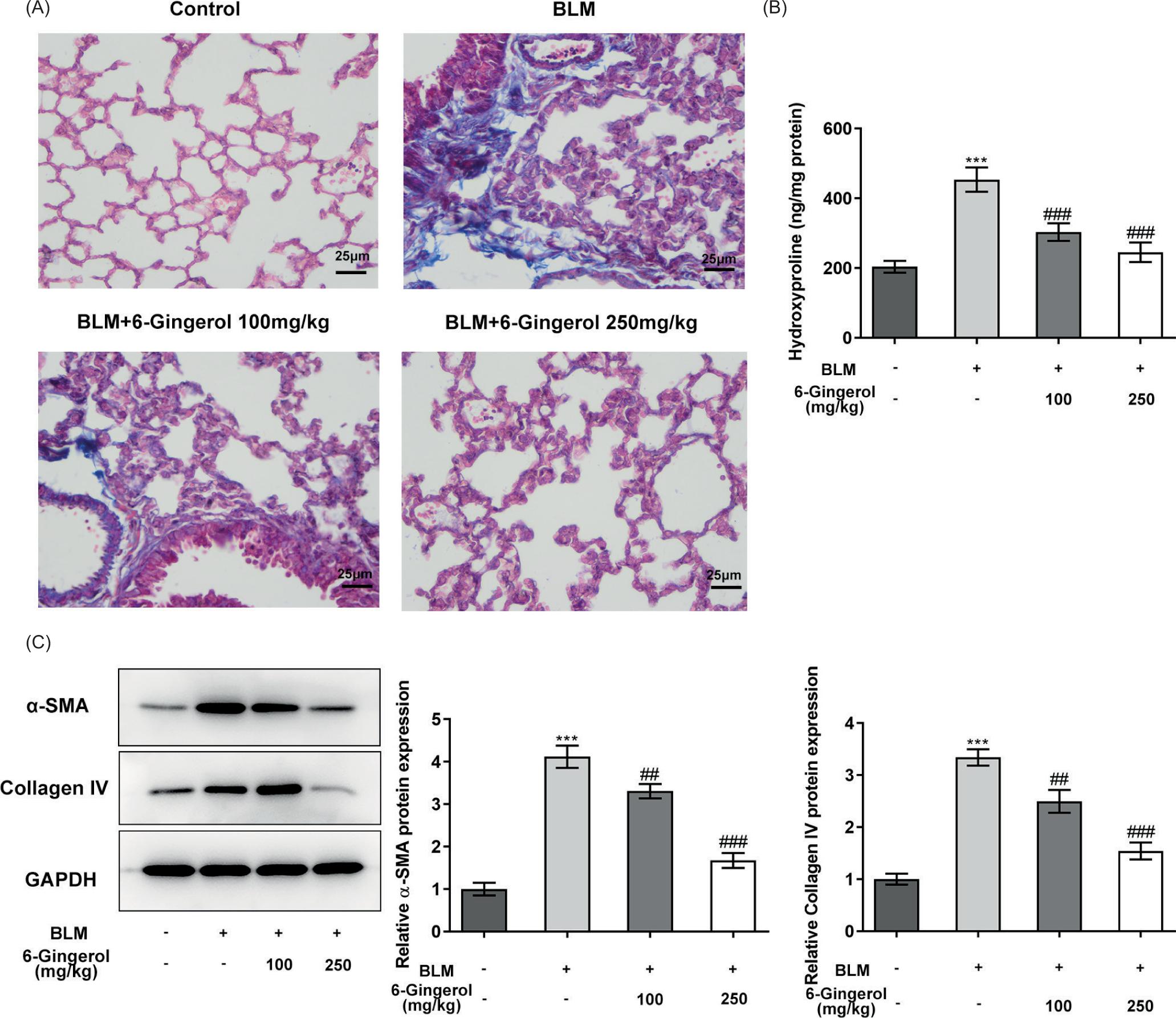
Moreover, the masson staining results indicated that the lung tissue of the control group had no inflammatory cell infiltration and fibrosis, whereas there were obvious inflammatory cells and blue collagen fibers in the BLM group. Meanwhile, the 6-gingerol 100 mg/kg treatment group only had a small amount of collagen fibers, and the 6-gingerol 250 mg/kg treatment group had no significant difference compared with the control group. Briefly, the degree of inflammation and fibrosis of lung tissue in the 6-gingerol intervention group was better than that of the BLM group and worse than that of the control group (Figure 3A). Subsequently, the levels of hydroxyproline in the lung tissues were determined. The level of hydroxyproline in the BLM group was significantly higher than that of the control group, and hydroxyproline in the lung tissues of mice were significantly reduced by the administration of 6-Gingerol (p < 0.001; Figure 3B). The levels of fibrotic deposition proteins were then determined by western blotting. The levels of α-SMA and collagen IV in the BLM group were significantly higher than those in the control group (p < 0.001), and 6-gingerol intervention could significantly reduce the expression levels of these two proteins (p < 0.01; Figure 3C).
Figure 3 6-Gingerol reduces lung fibrosis in bleomycin-induced mice. (A) Fibrotic changes in lung tissue were observed by masson staining. (B) The levels of hydroxyproline in the lung tissues were determined using colorimetry. (C) The levels of fibrotic deposition proteins were determined by western blotting. ***p < 0.001 vs. control; ##p < 0.01, ###p < 0.001 vs. BLM.
6-Gingerol regulates the ratio of cells and inhibits the release of inflammatory factors in the alveolar lavage fluid
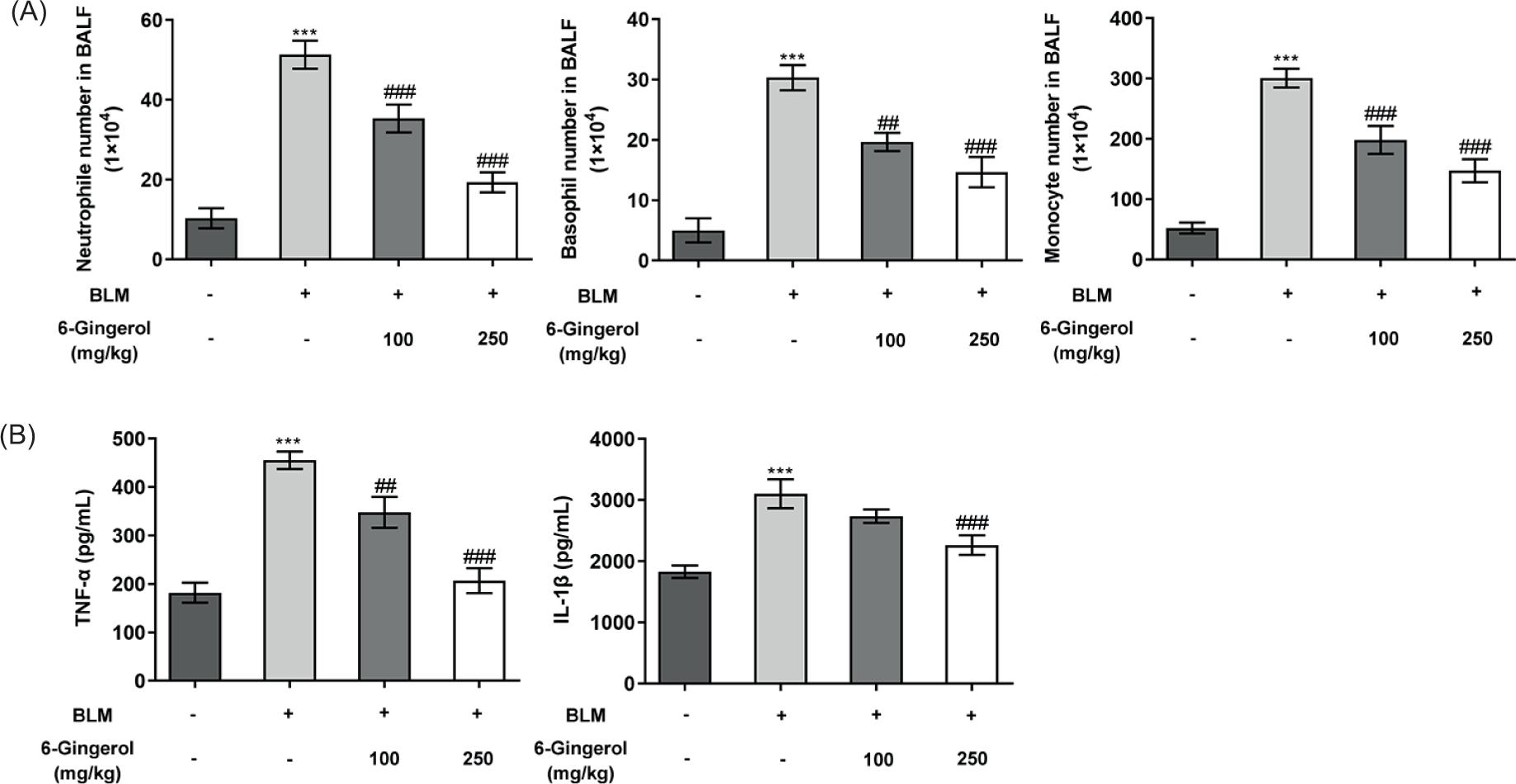
Elastase produced by neutrophils activates TGF-β and recruits inflammatory cells to the lungs, thereby promoting lung fibrosis.22 The number of basophils and monocytes is positively correlated with the progression of pulmonary fibrosis.24,25 From the experimental results, it could be seen that the number of neutrophils, basophils, and monocytes in the BLM group rose sharply (p < 0.001), while the number of these cells in the 6-gingerol intervention groups was smaller than those in the BLM group, and the difference became more obvious in the high-dose group (p < 0.01; Figure 4A). Moreover, the expression levels of TNF-α and IL-1β in the alveolar lavage fluid were determined using ELISA assay. The results revealed that the levels of TNF-α and IL-1β in the BLM group were significantly upregulated compared with the control group (p < 0.001), while 6-gingerol intervention again cut down TNF-α and IL-1β levels (p < 0.01), and the finding was more apparent in the high-dose group (p < 0.001; Figure 4B).
Figure 4 6-Gingerol regulates the ratio of cells and inhibits the release of inflammatory factors in the alveolar lavage fluid. (A) The numbers of neutrophils, basophils, and monocytes in the alveolar lavage fluid of each group of mice were calculated. (B) The levels of TNF-α and IL-1β in the alveolar lavage fluid were determined using ELISA assay. ***p < 0.001 vs. control; ##p < 0.01, ###p < 0.001 vs. BLM.
SIRT1 is the probable target of 6-gingerol
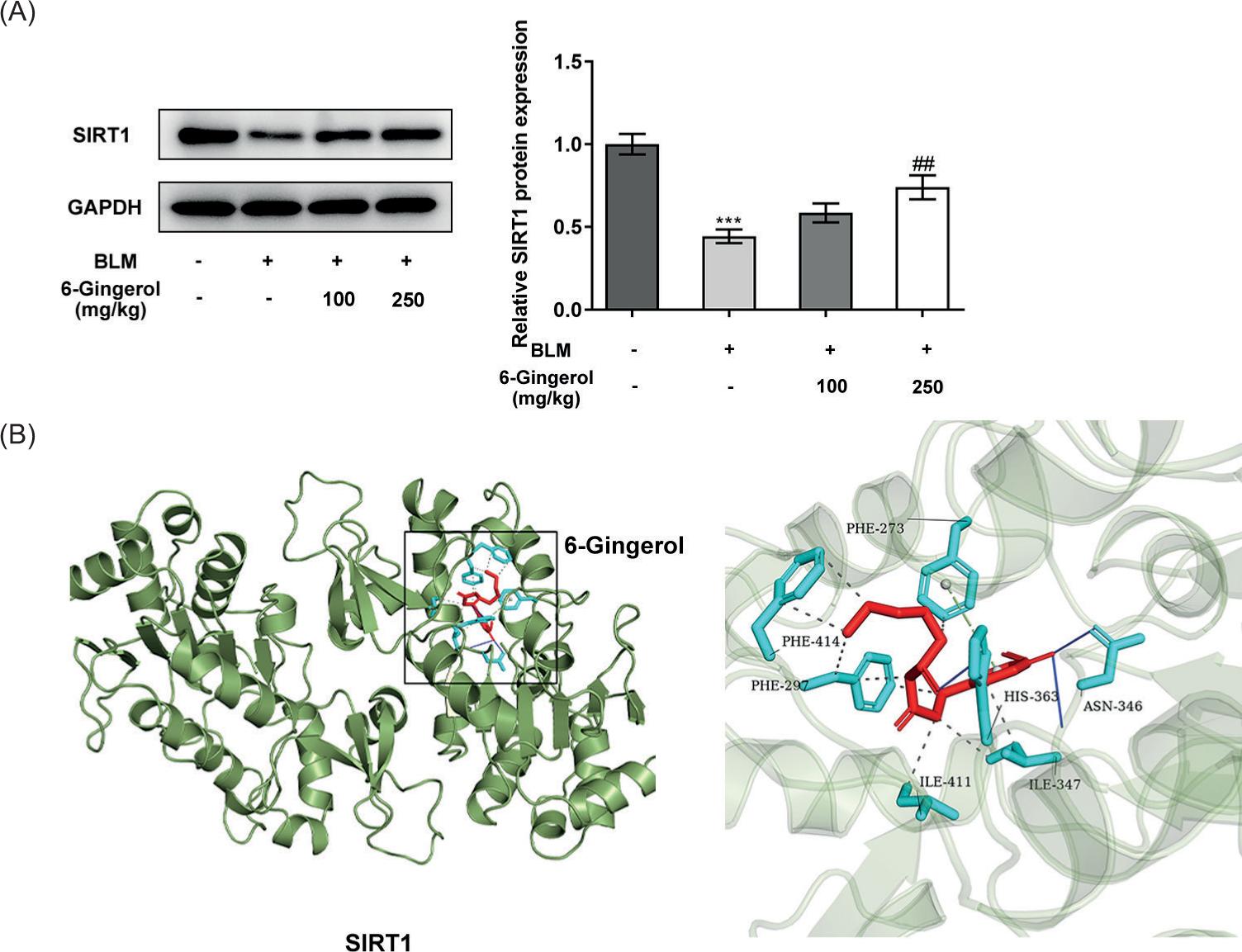
Furthermore, the expression of SIRT1 in the lung tissues at the protein level was determined using western blotting. It was noticed that SIRT1 exhibited low expression in the BLM group relative to the control group (p < 0.001). And, the upregulation of SITR1 in the 6-gingerol 250 mg/kg treatment group was distinctly highlighted compared with the BLM group (p < 0.01; Figure 5A). In addition, the results of molecular docking revealed that 6-gingerol was located in the hole of SIRT1 protein and bonded to the amino acid ASN in the form of hydrogen bonds, forming a π-stacking effect with PHE. These results suggested that 6-gingerol stably interacted with SIRT1 through noncovalent bonding (Figure 5B).
Figure 5 SIRT1 is the probable target of 6-gingerol. (A) The expression levels of SIRT1 in the lung tissues were determined using western blotting. (B) Overall image and partial enlarged image of the results of molecular docking. ***p < 0.001 vs. control; ##p < 0.01 vs. BLM.
6-Gingerol reduces TGF-β1-induced lung fibroblast proliferation and inflammatory and fibrotic matrix protein deposition by activating SIRT1
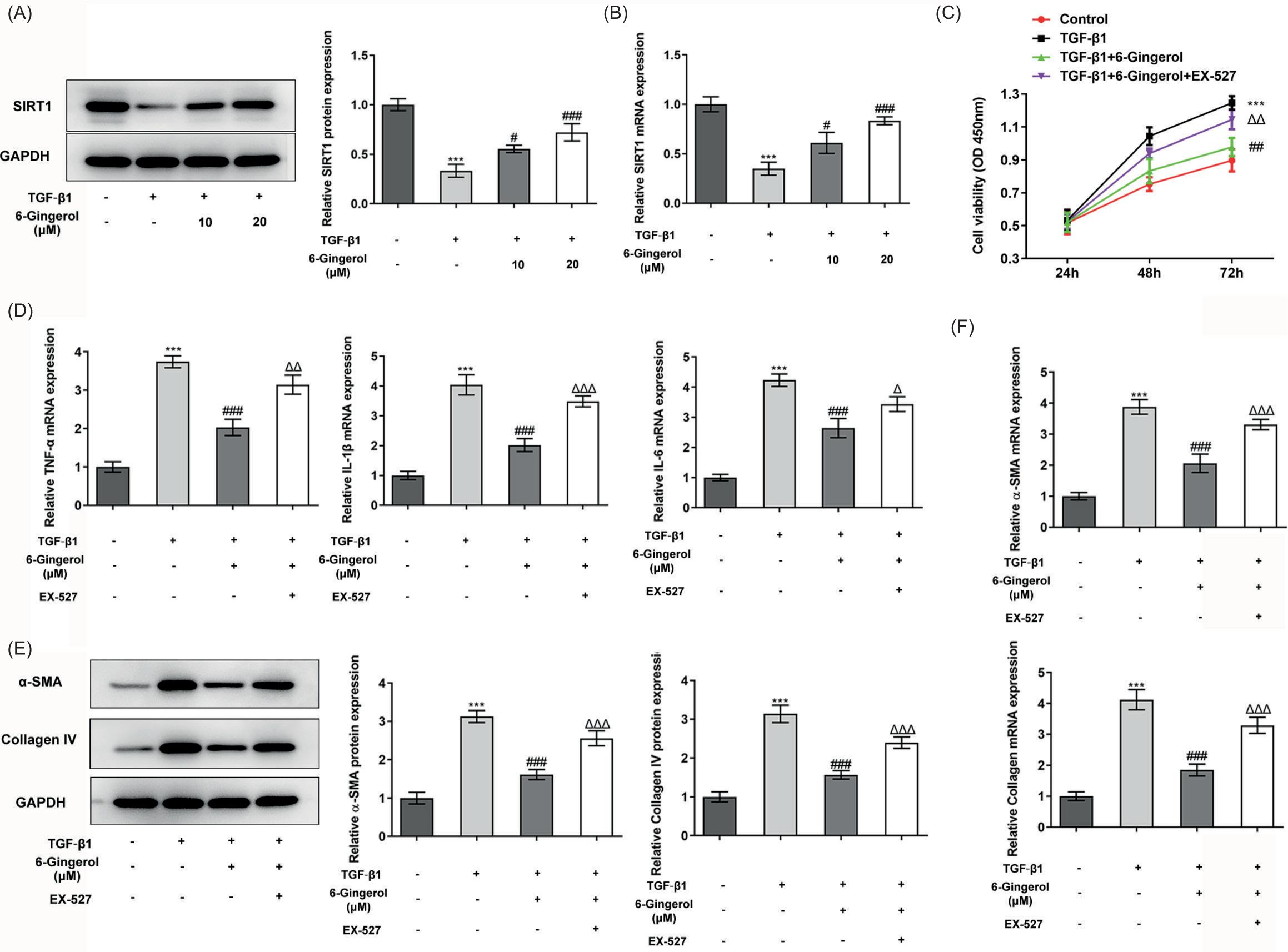
TGF-β1 is a central mediator in the pathogenesis of pulmonary fibrosis.26 Through our investigation, SIRT1 protein level in TGF-β1-stimulated cells was downregulated compared with the control group (p < 0.001), while its protein level was enhanced in the 6-gingerol intervention group than that in the TGF-β1 group (p < 0.05; Figure 6A-B). Afterwards, cell viability was determined using CCK-8 assay. The results demonstrated that TGF-β1 promoted the viability of fibroblasts, while 6-gingerol exerted the opposite role in cell viability as TGF-β1. However, SITR1 inhibitor, EX-527, eased the inhibitory effect of 6-gingerol on cell viability (p < 0.01; Figure 6C). Besides, RT-qPCR analysis showed that the levels of inflammatory factors including TNF-α, IL-1β, and IL-6 were greatly increased in the TGF-1β group (p < 0.001), and this effect was restored by 6-gingerol. Moreover, the addition of EX-527 could increase the levels of inflammatory factors again (p < 0.05; Figure 6D). Also, 6-gingerol significantly inhibited the overexpression of α-SMA and collagen IV induced by TGF-1β (p < 0.001), whereas this outcome was counteracted by EX-527 (p < 0.001; Figure 6E–F).
Figure 6 6-Gingerol reduces TGF-β1-induced lung fibroblast proliferation and inflammatory and fibrotic matrix protein deposition by activating SIRT1. (A) The expression levels of SIRT1 in the different groups of MRC-5 cells were determined using western blotting and (B) RT-qPCR. (C) Cell viability was determined using CCK-8 assay. (D) The expression levels of TNF-α, IL-1β, and IL-6 in the different groups of cells were determined using RT-qPCR. The expression levels of α-SMA and collagen IV in the different groups of cells were determined using (E) western blotting and (F) RT-qPCR. ***p < 0.001 vs. control; #p < 0.05, ###p < 0.001 vs. TGF-1β; Δp < 0.05, ΔΔp < 0.01, ΔΔΔp < 0.001 vs. TGF-1β + 6-gingerol.
Discussion
Pulmonary fibrosis has the highest morbidity in population aged 40–50 years, and it tends to occur in men.27 The clinical manifestations are dyspnea, dry cough, and fatigue, accompanied by clubbing digits and cyanosis.28,29 The disease can be diagnosed based on clinical manifestations, imagological examination, and lung function tests.30 Currently, the treatment of patients with pulmonary fibrosis is basically not limited to one method, and a comprehensive treatment strategy is usually adopted, that is, the combination of anti-pulmonary fibrosis drugs and nonpharmacological methods.31 Pirfenidone and nintedanib are the most commonly used anti-fibrosis drugs approved for marketing, which can delay pulmonary fibrosis to a certain extent and improve the lung function of patients.32 Nonpharmacological methods include pulmonary rehabilitation (such as aerobic training, respiratory muscle training), oxygen therapy, invasive/noninvasive mechanical ventilation, and lung transplantation.33,34 However, the application of these methods is limited due to a list of disadvantages including high cost, low curative effect, insufficient transplantation donors, immune rejection, or poor patient compliance.8 Therefore, it is necessary to actively explore the pathogenesis of pulmonary fibrosis and develop novel intervention drugs.
The 6-gingerol used in this study is a natural medicinal component that exists in ginger. It was found to increase the survival rate of experimental mice and reduce pathological damage to the lungs in the mice models. Previous studies have revealed that 6-gingerol can reduce the damage of dextran sodium sulfate–induced ulcerative colitis,35 and relieve liver damage induced by sepsis.36 Moreover, 6-gingerol is believed to reduce the release of pro-inflammatory and pro-fibrotic factors during liver fibrosis.37 Although the target organs of the study are different, the finding in our study that 6-gingerol alleviated fibrosis coincides with the results of previous studies. Furthermore, higher concentration of 6-gingerol illustrated more effective inhibition of pulmonary fibrosis and improvement of survival rate of mice. This suggested that the effect of 6-gingerol on pulmonary fibrosis was concentration-dependent. Although, as a natural extract, 6-gingerol has superior safety in contrast to synthetic drugs, the dose of drug used is not negligible for the treatment. Whether elevating the experimental concentration of 6-gingerol will adversely affect the survival of mice requires further research.
In addition, we conducted a preliminary study on the mechanism of 6-gingerol on pulmonary fibrosis. Through computer simulation calculations, this paper found that 6-Gingerol and SIRT1 had a stable binding through intermolecular forces. And, 6-gingerol markedly increased SIRT1 level in the lung tissue according to the western blot analysis. SIRT1 therein is a member of the Sirtuin family, which is a type of histone deacetylase that depends on NAD+.38 Interestingly, Conti et al. found that SIRT1 expression was downregulated in the lung tissue of patients with chronic obstructive pulmonary emphysema.39 Li et al. found that SIRT1 activation could alleviate lung injury caused by lipopolysaccharide.40 These findings indicated that the activation of SIRT1 was beneficial for normal lung function. Herein, 6-gingerol was found to inhibit lung fibroblast proliferation, reduce inflammation, and ensure fibrotic matrix protein deposition via activating SIRT1. The results of this article also illustrated the importance of SIRT1 in the lungs, and the thought of applying drugs to activate SIRT1 is an important and innovative approach to treat pulmonary fibrosis. Moreover, natural extract generally has little adverse effects to the human body, which is a significant advantage of its clinical application in the future.
Last but not least, SIRT1 has a variety of substrates, including transcription factors such as p53, p65, and Forkhead box O41; nuclear receptors such as estrogen receptor and peroxisome proliferators–activated receptor42; DNA repair protein components such as apurinic/apyrimidinic endonuclease and NBS1; and intracellular signaling molecules such as signal transducer and activator of transcription.43 This article is limited to the discovery that SIRT1 is the target of 6-gingerol; however, what kind of substrate SIRT1 acts on to regulate pulmonary fibrosis will be the direction of our future research.
Conclusion
In conclusion, 6-gingerol was found to alleviate pulmonary fibrosis by activating SIRT1. This article revealed the role of 6-gingerol in suppressing pulmonary fibrosis for the first time. This discovery is expected to underline the importance of 6-gingerol and provide credible evidence that 6-gingerol may be used as a potential medicine.
Funding
This study was supported by The Hospital of Chengdu University of Traditional Chinese Medicine Scientific Research Capacity to enhance the “Hundred Talents Plan” (Li Liu, No. 20-Q06), The Discipline Innovation Team Project of Shaanxi University of Traditional Chinese Medicine Innovation Team (Nan Yu, No. 2019-QN09), Provincial Unit Chinese Medicine Development Fund-Provincial Famous Traditional Chinese Medicine Studio (Yun Lu, No. 2100601), Scientific Research Project of Affiliated Hospital of Shaanxi University of Chinese Medicine (Hui Yuan, No. 2020QN011) and National Natural Science Foundation of China Youth Fund (Xinxin Xia, No.81904073).
REFERENCES
1. Hsia CC, Hyde DM, Weibel ER. Lung structure and the intrinsic challenges of gas exchange. Comprehens Physiol. 2016 Mar;6(2):827–895. 10.1002/cphy.c150028
2. Li XH, Luo ZQ. [Research progress of endogenous bone marrow-derived mesenchymal stem cells in pulmonary fibrosis]. Sheng li xue bao: [Acta Physiol Sinica]. 2020 Oct;72(5):597–604.
3. Belloli EA, Beckford R, Hadley R, Flaherty KR. Idiopathic non-specific interstitial pneumonia. Respirology (Carlton, Vic). 2016 Feb 21(2):259–268. 10.1111/resp.12674
4. McLean-Tooke A, Moore I, Lake F. Idiopathic and immune-related pulmonary fibrosis: diagnostic and therapeutic challenges. Clin Transl Immunol. 2019;8(11):e1086. 10.1002/cti2.1086
5. Adegunsoye A, Vij R, Noth I. Integrating genomics into management of fibrotic interstitial lung disease. Chest. 2019 May;155(5):1026–1040. 10.1016/j.chest.2018.12.011
6. Cottin V, Wollin L, Fischer A, Quaresma M, Stowasser S, Harari S. Fibrosing interstitial lung diseases: knowns and unknowns. Eur Respir Rev. 2019 Mar;28(151):180100. 10.1183/16000617.0100-2018
7. Kolb M, Bondue B, Pesci A, Miyazaki Y, Song JW, Bhatt NY, et al. Acute exacerbations of progressive-fibrosing interstitial lung diseases. Eur Respir Rev. 2018 Dec;27(150):180071. 10.1183/16000617.0071-2018
8. Nwafor EO, Lu P, Liu Y, Peng H, Qin H, Zhang K, et al. Active components from traditional herbal medicine for the potential therapeutics of idiopathic pulmonary fibrosis: a systemic review. Am J Chin Med. 2021;49(5):1093–1114. 10.1142/S0192415X2150052X
9. Snijder J, Peraza J, Padilla M, Capaccione K, Salvatore MM. Pulmonary fibrosis: a disease of alveolar collapse and collagen deposition. Exp Rev Respir Med. 2019 Jul;13(7):615–619. 10.1080/17476348.2019.1623028
10. Hewitt RJ, Maher TM. Idiopathic pulmonary fibrosis: new and emerging treatment options. Drugs Aging. 2019 Jun;36(6):485–492. 10.1007/s40266-019-00647-y
11. Haniadka R, Saldanha E, Sunita V, Palatty PL, Fayad R, Baliga MS. A review of the gastroprotective effects of ginger (Zingiber officinale Roscoe). Food Funct. 2013 Jun;4(6):845–855. 10.1039/c3fo30337c
12. Semwal RB, Semwal DK, Combrinck S, Viljoen AM. Gingerols and shogaols: important nutraceutical principles from ginger. Phytochemistry. 2015 Sep;117:554–568. 10.1016/j.phytochem.2015.07.012
13. Kato A, Higuchi Y, Goto H, Kizu H, Okamoto T, Asano N, et al. Inhibitory effects of Zingiber officinale Roscoe derived components on aldose reductase activity in vitro and in vivo. J Agric Food Chem. 2006 Sep;54(18):6640–6644. 10.1021/jf061599a
14. Wei QY, Ma JP, Cai YJ, Yang L, Liu ZL. Cytotoxic and apoptotic activities of diarylheptanoids and gingerol-related compounds from the rhizome of Chinese ginger. J Ethnopharmacol. 2005 Nov;102(2):177–184. 10.1016/j.jep.2005.05.043
15. Yao J, Du Z, Li Z, Zhang S, Lin Y, Li H, et al. 6-Gingerol as an arginase inhibitor prevents urethane-induced lung carcinogenesis by reprogramming tumor supporting M2 macrophages to M1 phenotype. Food Funct. 2018 Sep;9(9):4611–4620. 10.1039/C8FO01147H
16. Hasani H, Arab A, Hadi A, Pourmasoumi M, Ghavami A, Miraghajani M. Does ginger supplementation lower blood pressure? A systematic review and meta-analysis of clinical trials. Phytother Res. 2019 Jun;33(6):1639–1647. 10.1002/ptr.6362
17. Tramontin NDS, Luciano TF, Marques SO, de Souza CT, Muller AP. Ginger and avocado as nutraceuticals for obesity and its comorbidities. Phytother Res. 2020 Jun;34(6):1282–1290. 10.1002/ptr.6619
18. Choi JG, Kim SY, Jeong M, Oh MS. Pharmacotherapeutic potential of ginger and its compounds in age-related neurological disorders. Pharmacol Therap. 2018 Feb;182:56–69. 10.1016/j.pharmthera.2017.08.010
19. Wang X, Shen Y, Thakur K, Han J, Zhang JG, Hu F, et al. Antibacterial activity and mechanism of ginger essential oil against Escherichia coli and Staphylococcus aureus. Molecules (Basel, Switzerland). 2020 Aug 25(17):3955. 10.3390/molecules25173955
20. Han X, Liu P, Liu M, Wei Z, Fan S, Wang X, et al. [6]-Gingerol ameliorates ISO-induced myocardial fibrosis by reducing oxidative stress, inflammation, and apoptosis through inhibition of TLR4/MAPKs/NF-κB pathway. Mol Nutr Food Res. 2020 Jul;64(13):e2000003. 10.1002/mnfr.202000003
21. Liu T, De Los Santos FG, Phan SH. The bleomycin model of pulmonary fibrosis. Methods Mol Biol. 2017;1627:27–42. 10.1007/978-1-4939-7113-8_2
22. Kolahian S, Fernandez IE, Eickelberg O, Hartl D. Immune mechanisms in pulmonary fibrosis. Am J Respir Cell Mol Biol. 2016 Sep;55(3):309–322. 10.1165/rcmb.2016-0121TR
23. Lee YS, Min D, Park SY, Lee J, Bae H. Standardized herbal extract PM014 alleviates fine dust-induced lung inflammation in mice. BMC Complement Med Ther. 2020 Sep;20(1):270. 10.1186/s12906-020-03060-w
24. Kreuter M, Lee JS, Tzouvelekis A, Oldham JM, Molyneaux PL, Weycker D, et al. Monocyte count as a prognostic biomarker in patients with idiopathic pulmonary fibrosis. Am J Respir Crit Care Med. 2021 Jul;204(1):74–81. 10.1164/rccm.202003-0669OC
25. Gieseck RL, 3rd, Wilson MS, Wynn TA. Type 2 immunity in tissue repair and fibrosis. Nat Rev Immunol. 2018 Jan;18(1):62–76. 10.1038/nri.2017.90
26. Nanri Y, Nunomura S, Terasaki Y, Yoshihara T, Hirano Y, Yokosaki Y, et al. Cross-talk between transforming growth factor-β and periostin can be targeted for pulmonary fibrosis. Am J Respir Cell Mol Biol. 2020 Feb;62(2):204–216. 10.1165/rcmb.2019-0245OC
27. Kishaba T. Acute exacerbation of idiopathic pulmonary fibrosis. Medicina (Kaunas, Lithuania). 2019 Mar 55(3):70. 10.3390/medicina55030070
28. Patterson KC, Strek ME. Pulmonary fibrosis in sarcoidosis. Clinical features and outcomes. Ann Am Thoracic Soc. 2013 Aug 10(4):362–370. 10.1513/AnnalsATS.201303-069FR
29. Glass DS, Grossfeld D, Renna HA, Agarwala P, Spiegler P, Kasselman LJ, et al. Idiopathic pulmonary fibrosis: molecular mechanisms and potential treatment approaches. Respir Investig. 2020 Sep;58(5):320–335. 10.1016/j.resinv.2020.04.002
30. Wakwaya Y, Brown KK. Idiopathic pulmonary fibrosis: epidemiology, diagnosis and outcomes. Am J Med Sci. 2019 May;357(5):359–369. 10.1016/j.amjms.2019.02.013
31. Maher TM, Strek ME. Antifibrotic therapy for idiopathic pulmonary fibrosis: time to treat. Respir Res. 2019 Sep;20(1):205. 10.1186/s12931-019-1161-4
32. Somogyi V, Chaudhuri N, Torrisi SE, Kahn N, Müller V, Kreuter M. The therapy of idiopathic pulmonary fibrosis: what is next? Eur Respir Rev. 2019 Sep;28(153):190021. 10.1183/16000617.0021-2019
33. Faverio P, De Giacomi F, Sardella L, Fiorentino G, Carone M, Salerno F, et al. Management of acute respiratory failure in interstitial lung diseases: overview and clinical insights. BMC Pulm Med. 2018 May;18(1):70. 10.1186/s12890-018-0643-3
34. Schaffer JM, Singh SK, Reitz BA, Zamanian RT, Mallidi HR. Single-vs double-lung transplantation in patients with chronic obstructive pulmonary disease and idiopathic pulmonary fibrosis since the implementation of lung allocation based on medical need. JAMA. 2015 Mar;313(9):936–948. 10.1001/jama.2015.1175
35. Sheng Y, Wu T, Dai Y, Xu L, Zhong Y, Xue Y, et al. 6-gingerol alleviates inflammatory injury in DSS-induced ulcerative colitis mice by regulating NF-κB signaling. Ann Palliat Med. 2020 Jul;9(4):1944–1952. 10.21037/apm-20-903
36. Hong MK, Hu LL, Zhang YX, Xu YL, Liu XY, He PK, et al. 6-Gingerol ameliorates sepsis-induced liver injury through the Nrf2 pathway. Int Immunopharmacol. 2020 Mar;80:106196. 10.1016/j.intimp.2020.106196
37. Algandaby MM, El-Halawany AM, Abdallah HM, Alahdal AM, Nagy AA, Ashour OM, et al. Gingerol protects against experimental liver fibrosis in rats via suppression of pro-inflammatory and profibrogenic mediators. Naunyn Schmiedebergs Arch Pharmacol. 2016 Apr;389(4):419–428. 10.1007/s00210-016-1210-1
38. Chen C, Zhou M, Ge Y, Wang X. SIRT1 and aging related signaling pathways. Mech Ageing Dev. 2020 Apr;187:111215. 10.1016/j.mad.2020.111215
39. Conti V, Corbi G, Manzo V, Malangone P, Vitale C, Maglio A, et al. SIRT1 activity in peripheral blood mononuclear cells correlates with altered lung function in patients with chronic obstructive pulmonary disease. Oxid Med Cell Longev. 2018;2018:9391261. 10.1155/2018/9391261
40. Li T, Zhang J, Feng J, Li Q, Wu L, Ye Q, et al. Resveratrol reduces acute lung injury in a LPS-induced sepsis mouse model via activation of Sirt1. Mol Med Rep. 2013 Jun;7(6):1889–1895. 10.3892/mmr.2013.1444
41. Liu X, Zhao H, Jin Q, You W, Cheng H, Liu Y, et al. Resveratrol induces apoptosis and inhibits adipogenesis by stimulating the SIRT1-AMPKα-FOXO1 signalling pathway in bovine intramuscular adipocytes. Mol Cell Biochem. 2018 Feb;439(1–2):213–223. 10.1007/s11010-017-3149-z
42. Wang W, Lin Q, Lin R, Zhang J, Ren F, Zhang J, et al. PPARα agonist fenofibrate attenuates TNF-α-induced CD40 expression in 3T3-L1 adipocytes via the SIRT1-dependent signaling pathway. Exp Cell Res. 2013 Jun;319(10):1523–1533. 10.1016/j.yexcr.2013.04.007
43. Su GC, Yeh HY, Lin SW, Chung CI, Huang YS, Liu YC, et al. Role of the RAD51-SWI5-SFR1 ensemble in homologous recombination. Nucl Acids Res. 2016 Jul;44(13):6242–6251. 10.1093/nar/gkw375