
Download

Research Article
Distribution, molecular characterization, and treatment options for dermatophyte and non-dermatophyte fungi isolated from human and animal samples: An integrated computational and experimental approach
Saba Sanaa*, Ahsaan Ahmedb, Layyaba Nazira, Usman Shehzadc, Rukhmaa, Muhammad Nauman Aftaba, Najeeb Ullahd, Nawal Al-Hoshanie, Maher S. Alwethaynanif, Hanan Abdulrahman Saginig, Shaza N. Alkhatibh, Fakhria A. Al-Joufii
aDepartment of Microbiology, Dr. Ikram-ul-Haq Institute of Industrial Biotechnology, Government College University, Lahore, Punjab, Pakistan
bDepartment of Pharmaceutical Chemistry, Institute of Pharmaceutical Sciences, Jinnah Sindh Medical University, Karachi, Pakistan
cDepartment of Pathology, Shoaukat Khanam Memorial Cancer Hospital & Research Centre, Lahore, Punjab, Pakistan
dFood and Biotechnology Research Center, PCSIR Laboratories Complex, Lahore, 54590 Punjab, Pakistan
eDepartment of Biology, College of Science, Princess Nourah bint Abdulrahman University, P.O. Box 84428, Riyadh 11671, Saudi Arabia
fDepartment of Clinical Laboratory Sciences, College of Applied Medical Sciences, Shaqra University, Alquwayiyah, Riyadh, Saudi Arabia
gDepartment of Biological Sciences, College of Sciences, University of Jeddah, 21959, Jeddah Saudi Arabia
hDepartment of Biological Sciences, Collage of Sciences and Arts Khulais, University of Jeddah, Jeddah, Saudi Arabia
iDepartment of Pharmacology, College of Pharmacy, Jouf University, 72341 Aljouf, Saudi Arabia
Abstract
Background: Cutaneous infections caused by dermatophytes and non-dermatophytes fungi have become significant public health and veterinary concern.
Objective: The current study deals with the distribution, characterization, and antifungal susceptibility profile of these fungi.
Materials and Methods: A total of 40 sample were screened by KOH mount; positive samples were cultured on SDA. Purified isolates were identified by macroscopic and microscopic characters. Molecular characterization by PCR and sanger sequencing of ITS region confirmed the fungi. Evolutionary trees were constructed using MEGA 11. Different treatment options were evaluated, including amphotericin B, nystatin, fluconazole, itraconazole, and essential oil of Nigella sativa. Initially molecular docking was performed against selected fungal targets (5eqb and 5v5z) using molecular-operating environment (MOE v.2018.01) and Maestro modeling interface 12.3. In vitro antimycotic activity was performed by agar disc diffusion, followed by micro-broth dilution method.
Results: KOH mount revealed that 65% (13/20) samples positive for fungal components. Culture positivity for samples was 23.07% for dermatophytes and 76.92% for non-dermatophytes. Dermatophytes were detected in skin samples. However, non-dermatophytes were isolated from all types of samples. Sequence analysis confirmed the distribution of Arthroderma multifidum, Aspergillus sydowii, and Aspergillus sublatus. In-silico analysis revealed itraconazole as the most effective antimycotic agent with -7.93 kcal/mol (5eqb) and -10.89 (5v5z) kcal/mol. In vitro analysis strengthens docking analysis, itraconazole was the most effective agent with the highest mean ZOI against A. multifidum (72.00±3.46 mm), A. sydowii (62.00±3.00 mm), and A. sublatus (64.00±1.73 mm). Moreover, essential oil of N. sativa was the most effective antifungal against A. sydowii (ZOI; 31.00±3.46 mm, MIC; 3.125±0.00 mg/mL).
Conclusion: The current study is the first report of A. sydowii and A. sublatus in onychomycosis and hair infections in Pakistan. This study provides awareness about the existence of new fungal pathogens and highlights the availability of effective treatment options for emerging fungal infections.
Key words: Arthroderma multifidum, Aspergillus sublatus, Aspergillus sydowii, Azole antifungals, Cutaneous infections, ITS, Molecular docking, Thymoquinone
*Corresponding authors: Saba Sana, Department of Microbiology, Dr. Ikram-ul-Haq Institute of Industrial Biotechnology, Government College University, Lahore, Pakistan. Email address: [email protected]
Received 11 July 2025; Accepted 25 September 2025; Available online 1 January 2026
Copyright: Sana S, et al.
This open access article is licensed under Creative Commons Attribution 4.0 International (CC BY 4.0). http://creativecommons.org/licenses/by/4.0/
Introduction
Cutaneous infections are caused by dermatophyte and non-dermatophyte fungal species. These are pathogens of public health concern. Cutaneous fungal infections infect the keratinized tissue of skin, hair, and nails1. Approximately 20–25% of the world’s population is affected by cutaneous fungal infections with 40–60% incidence of dermatophytes. However, non-dermatophytes are reported for 0.6–47% in different regions. Among dermatophytes, following are the commonly reported nine genera: Trichophyton, Epidermophyton, Nannizia, Paraphyton, Lopophyton, Microsporum, Arthroderma, Ctenomyces, and Guarromyces. However, most of the pathogenic species belongs to the genus Trichophyton, Microsporum, Epidermophyton, and Nannizzia. Dermatophytes can be grouped as zoophilic, geophilic, and anthropophilic.2
Non-dermatophyte molds (NDMs) are soil saprophyte and are recognized as plant pathogens. Recent reports have acknowledged the presence of NDMs in onychomycosis3. However, exclusion of environmental contaminants is important to avoid false diagnosis.4 Most of the NDMs belong to the genera Alternaria, Acremonium, Fusarium, Scopulariopsis, Scytalidium, and Aspergillus. The causative agent of the disease may vary depending upon geographical region. Irrational use of antimicrobials, immunosuppression, invasive procedures, and chemotherapy are the leading causes for rise in infections caused by NDMs.5
Aspergilli are the second most common cause of opportunistic fungal infections after Coandida albicans.6 Aspergillus is the most commonly isolated genus from human clinical samples. Among aspergilli, the most common fungal pathogen is Aspergillus fumigates, followed by Aspergillus flavus, Aspergillus niger, and Aspergillu terreus. Aspergilli are reported to cause respiratory infections, followed by skin infections. Cutaneous aspergillosis may occur as primary as well as secondary infection. Immunocompromised and immunocompetent individuals are equally susceptible to aspergillosis.6 However, immunocompromised individuals are more likely to acquire fungal infection. Cutaneous infections, particularly in immunocompetent individuals, are a rare phenomenon. Aspergillosis primarily affects the lungs, sinuses, and central nervous system (CNS).7
In developing countries, such as Pakistan, socioeconomic constraints and other common health issues have contributed to a lack of awareness about fungal infections. Thus, even in the presence of good personal hygiene, it continued to persist.5 One of the risk factors of fungal infections is close contact with animals. Therefore, individuals in veterinary professions or those rearing the animals may acquire fungal infections easily9. Some dermatophytes also have zoonotic potential.10 Good hygienic measures, immune competency, and safe animal handling are the practices to prevent fungal infections. Furthermore, accurate diagnosis, better treatment options, and knowledge about pathogens play an important role in controlling fungal infections. Hence, limited awareness and treatment challenges have become major contributors of rising fungal infections.
The current scenario of antifungal resistance, limited efficacy, and toxicity urges the urgent need for development of new drugs.11 In the current situation, molecular docking has gained importance, which predicts interaction between antifungal compounds and fungal targets, such as enzymes, proteins, and DNA. To evaluate a ligand’s binding affinity, the technique involves sampling of every potential ligand conformation within a target’s binding pocket and grading each with respect to scoring functions. This approach assists investigators to discover new drug targets and medicinal compounds by allowing these to comprehend molecular interactions, which are necessary for disease pathways. Its use in the drug development process reduces the need for resource-intensive experimental techniques while simultaneously accelerating the discovery of potential drug candidates.12 To the best of present authors’ knowledge, the current study is the first investigation conducted in Pakistan about the presence of dermatophyte molds (DMs) and NDMs in cutaneous infections, their molecular characterization, computational analysis of antifungals, and in vitro evaluation of different antifungals as treatment options to control these infections.
Materials and Methods
Cutaneous infections may result from dermatophyte or non-dermatophyte fungi. The current study deals with distribution and molecular characterization of dermatophyte and non-dermatophyte fungi from human and animal samples. Further In-silico and In vitro effects of different antifungals was evaluated against isolated fungi. The study was conducted from June 2021 to May 2022.
Samples
Clinical samples (n=40) of infected skin, hair and nails were collected in Lahore, Pakistan. Skin samples (n=20) were collected from animals (cats), and hair (n=10) and nail cutting (n=10) samples were of human origin. The cat samples were collected from pet center, University of Veterinary and Animal Sciences Lahore. Nail clippings and hair cuttings were obtained from a community in Lahore. Infected skin was decontaminated with 70% ethyl alcohol to remove dirt and environmental contamination. Skin scraps were collected from the periphery of lesions with sterile blunt end surgical blades. The scraps were collected on black paper and shifted to sterile vials. Hair samples were plucked with disinfected forceps, and nail clippings were obtained with a sterile cutter. The samples were properly labeled and brought to laboratory (BSL-2) for further processing.13 Experiments were performed under sterile containment in BSC Class 2.
Direct microscopic examination
The microscopic examination of each sample was performed. Potassium hydroxide (KOH) mounts were prepared using 40% solution. The mounts were observed under brightfield compound microscope at 100× and 400× magnification. Approximately 50–100 microscopic fields were observed for each sample. Positive samples for fungal elements (hyphae and spores) were selected for culturing on selective agar.14
Culturing and purification of fungi
For the isolation of fungi, positive samples were inoculated on Sabouraud dextrose agar (SDA) supplemented with chloramphenicol (0.05 g/L) and cycloheximide (0.4 g/L) to avoid bacterial contamination. After inoculation, plates were incubated at 25°C for 5–10 days. The colonies that appeared on SDA were purified by single spot technique on the same media.15
Identification of Fungi
Fungi were identified by their macroscopic and microscopic characters. Macroscopic characters of fungi were observed on the obverse and reverse side of the culture plate. The characters observed were growth pattern, colony textures, colony color, and diffusible pigments. Microscopic characters were studied by the slide culture technique. Prepared fungal slides were observed for type of hyphae, shape, arrangement of spores, shapes, and presence or absence of special structures using a compound microscope.16
Molecular Characterization of Fungi
Fungal mycelia were used for the extraction of DNA. The fungi were cultured in SDB in a shaking incubator and mycelia were harvested by filtration at the early stage of growth. The fungal mycelia (100 mg) were used for the extraction of DNA using commercially available DNA extraction kit (Thermo Scientific GeneJET Genomic DNA purification kit) by following manufacturer’s instructions. DNA was quantified by nanodrop and observed on 1% agarose. Polymerase chain reaction (PCR) was carried out in MiniAmp™ Thermal Cycler. The Internal Transcribed Spacer (ITS) was targeted to amplify the genome. ITS1 and ITS2 primer sequences were used:
Forward primer: (ITS1) F:5′-TCCGTAGGTGAACCTGCGG-3′)
Reverse primer: (ITS4) R:5′-TCCTCCGCTTATTGATATGC-3′)
PCR reaction mixture (25 µL) was prepared in PCR tubes using 2X master mix (12.5 µL), primers (ITS1 and ITS4; 1 µL each), nuclease-free water (6 µL) with template DNA (5 µL). PCR was carried by as described by Dalis et al. in 2018.17 The initial denaturation was carried out at 94°C for 3 min, denaturation at 94°C for 45 s, annealing at 52°C for 1 min, extension at 72°C for 1 min, and the final extension at 72°C for 7 min and for infinite (∞) time at 4°C. After completion of the process, agarose gel electrophoresis was carried out, and amplicons along with DNA ladder that was run on 2% agarose gel and gel was observed in UV transilluminator.
Sequence analysis
Polymerase chain reaction for positive samples was re-conducted and amplicons with primers were sent to Lab Genetics for sequence analysis. Sanger sequencing was performed, and results were further processed by the Bioedit software and phylogenetic analysis was performed using the MEGA software. Sequences were submitted to the National Center for Biotechnology Information (NCBI), and accession numbers were obtained.
Computational Analysis for Antifungal Susceptibility
The structure of reference standards was fetched online from PubChem (https://pubchem.ncbi.nlm.nih.gov/) and imported to the Molecular-Operating Environment (MOE) v.2018.01 software to get charged, add missing hydrogen atoms and optimization were done by applying MMFF94x force field to clean up the geometry and attain a stable pose.18
Macromolecular targets were identified and extracted from the protein data bank (https://www.pdb.org/) based on the mechanisms of action of standard drugs. Then Quickprep tool was used to perform energy minimization after removal of water molecules, heteroatoms (other than carbon and hydrogen atoms), and extra chains. The active binding site was marked and isolated by the site finder before proceeding toward molecular docking. The molecular docking technique was employed to examine interactions between the target protein and ligands. To closely investigate potential residual interactions between ligand and the target proteins 5eqb and 5v5z19,20, both ligand and macromolecules were uploaded onto the MOE software, incorporating induced fit molecular docking to account for structural adjustments in the receptor. The residual interactions of standards with the target enzyme were evaluated by observing the lowest energy pose with significant interaction via Maestro modeling interface v.12.3.
Antifungal susceptibility testing
Antifungal susceptibility of isolated dermatophyte and non-dermatophyte species was evaluated for different antifungals by agar well diffusion, followed by minimum inhibitory concentration (MIC). Experiments were performed in triplicate. Antifungal drugs (fluconazole, nystatin, amphotericin B. itraconazole), and essential oil of N. sativa were selected for antifungal activity.21 For antifungal activity, an inoculum was prepared in 10 mL of normal saline by mixing purified fungal spores. Inoculum was standardized to 106 spores/mL. Spores were counted using Neubauer chamber. The Muller Hinton agar (MHA) was used for agar well diffusion. For inoculation, a pour plate technique was used. The inoculum (100 µL) was taken in sterilized Petri plates with sterilized micropipettes. Then, molten medium (45–50°C) was poured into the plates. Antibiotic chloramphenicol (0.05 g/L) was added in medium prior to use so that the chances of contamination were reduced as much as possible. It was allowed to solidify. After solidification, wells were formed with well borers and the base of each well was sealed with sterilized molten agar. Then antifungals (100 µL) were added in wells and incubated at 25°C for 5–10 days in upright position in an incubator. Zone of inhibition (ZOI) was measured in millimeter (mm). For essential oil of N. sativa, a stock solution of 100 mg/mL was prepared and 100 µL of this stock was poured into respective well, followed by incubation at the above-mentioned conditions.22 MIC of essential oil of N. sativa was determined by the micro broth dilution method.
In micro broth dilution, Muller Hinton broth (MHB) was used in 96 well micro-titration plates. First, 100 µL of broth was added up to 12th well. Two-fold serial dilutions of 100 µL were prepared in 96 wells flat bottom micro-titration plates. The two-fold serial dilution of antifungal was performed by taking 100 µL of antifungal in the first well, then shifted to the secondwell and dilution was continued till the 10th well, and from the 10th well, remaining dilution was discarded. Then 100 µL of freshly prepared inoculum were added up to 11th well having different dilution of antifungals. The 11th well was positive control with MHB and fungal suspension, and the 12th well was negative/broth control containing MHB only. Micro-titration plates were placed in an incubator at 25°C for 5–10 days. MIC was determined by the absence of growth in micro-titration plates.23
Statistical analysis
The Statistical significance of experimental results was evaluated by Statistical Package for Social Sciences (SPSS version 20) by One-way Analysis of Variance (ANOVA), followed by Duncan’s multiple range test (DMRT).
Results
Mycological Examination of Samples
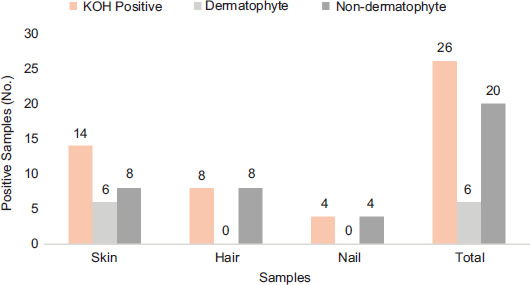
In all, 40 different infected samples were collected from different origins (skin = 20, hair = 10, and nails = 10). Out of these 40 samples, 65% (26/40) were positive by direct microscopy. Among these, four nail samples, 14 skin samples, and eight hair samples were positive. Under low- (100×) and high-power (400×) magnification, hyphal firmaments and spores were detected in these samples. Out of 40 samples, 14 (35%) were negative for the presence of fungal components. Positive samples were the cultures on SDA, and distribution of dermatophytes and non-dermatophytes fungi was determined. According to the culturing of samples (n=26), which were positive in direct microscopic examination, 23.07% samples were positive for dermatophytes, and 76.92% samples were found positive for non-dermatophyte fungal pathogens. The highest prevalence of dermatophytes was found in skin samples (six were positive for dermatophytes and 20 were positive for non-dermatophytes). No nail and hair sample showed the growth of dermatophytes. Hair samples (08) were positive for the non-dermatophyte group, and non-dermatophyte species were detected in four nail samples. Results are shown in Figure 1 and Table 1.
Figure 1 Distribution of fungal groups in selected samples. Samples positive in KOH mounts are represented by blue bars, and culture-positive dermatophytes and non-dermatophytes are represented by yellow and gray bars, respectively, in each category of sample.
Table 1 Distribution of fungal species in selected samples.
| Fungal group | Fungal species | Skin | Hair | Nail | Total |
|---|---|---|---|---|---|
| Dermatophytes | Arthroderma multifidum | 06 | 00 | 00 | 06 |
| Non-dermatophytes | Aspergillus sydowii | 06 | 08 | 04 | 18 |
| Aspergillus sublatus | 02 | 00 | 00 | 02 | |
| Total | 14 | 08 | 04 | 26 | |
Conventional Identification of Fungi
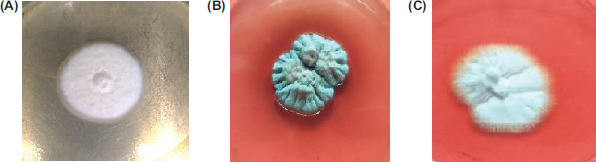
Fungi were identified by their macroscopic and microscopic characters. Macroscopic characters of dermatophytes and non-dermatophytes were observed on SDA. Microscopic characters were observed by slide culture technique. Representative pictures of isolated colonies are shown in Figure 2.
Figure 2 Representative pictures of isolated fungal colonies on Sabouraud’s dextrose agar after 5 days of incubation at 25°C. (A) Arthroderma multifidum; (B) Aspergillus sydowii; and (C) Aspergillus sublatus.
Arthroderma multifidum: According to macroscopic analysis, the colonies were pure white but turned to beige color on maturation. The texture of colonies was cottony. The colony had a uniform shape. On the reverse side of the plate, light yellow color pigmentation was observed in a matured colony. Microscopic examination by slide culture revealed septate hyaline hyphae, and small ovoid and pyriform macroconidia (Figure 2A).
Aspergillus sydowii: Initially, colonies were white in color, turning to bluish green with velvety texture on the obverse side of the plate. The reverse side showed yellowish brown pigment on matured growth. Microscopic observations showed the presence of hyaline septate hyphae spherical vesicle and conidia. Conidia were arranged in chains around the vesicle (Figure 2B).
Aspergillus sublatus: According to macroscopic character, the colonies were white in color with cotton texture on the obverse side of the plate. While on the reverse side, no pigment production was observed. The reverse side was also white. Microscopic characters observed in this species were hyaline septate hyphae and unicellular conidia arranged in chains around the vesicle (Figure 2C).
Distribution of dermatophytes and non-dermatophytes
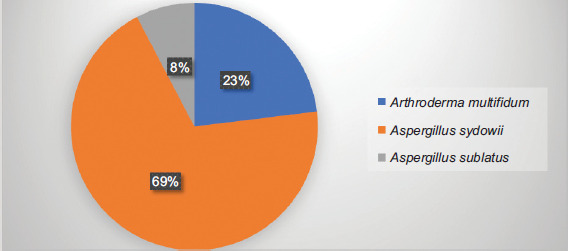
Different fungi from dermatophytes and non-dermatophytes were identified from positive samples. Among dermatophytes, A. multifidum were identified in skin samples. No fungi from dermatophyte group were detected in hair and nail samples. Among non-dermatophyte group, A. sydowii and A. sublatus were isolated from skin, hair, and nail samples (Table 1). Among dermatophytes, A. multifidum was the most prevalent species and detected only in skin samples of animal origin. Among non-dermatophytes, A. sydowii and A. sublatus were the prevalent species and detected in all type of samples (Figure 3). Fungi were identified by their macroscopic and microscopic characters. In all, 13 fungal species were identified. The percentage distribution of these fungi is shown in Figure 3. Some fungi remained unidentified at genus level and declared as fungi.
Figure 3 Percentage distribution of fungal species isolated from skin (n=20), hair cutting (n=10), and nail clipping (n=10) samples.
Molecular Characterization and Phylogenetic Analysis
Discrete fungi were selected for molecular characterization and phylogenetic analysis. According to PCR results, isolates produced an amplicon between 600 bps and 700 bps were observed at 2% agarose gel. According to sequence analysis using NCBI, the ITS region similarity analysis of selected isolates was discovered more with A. multifidum, A. sydowii, and A. sublatus on the available data. The sequences were submitted to NCBI database, and their accession numbers were OP102681 (A. multifidum), OP120769 (A. sydowii), OP132848 (A. sublatus), and OP351722 (unidentified fungi). The phylogenetic tree of these fungi is shown in Figure 4. According to phylogenetic analysis, A. multifidum (OP102681) was closely related to A. multifidum isolate 1 (OP102681). A. sydowii (OP120769), A. sublatus (OP132848), and unidentified fungi (OP351722) were close to A. sydowii ASH-01 (OP120769), A. sublatus ASS-03 (OP132848), and fungi MMJ-04 (OP351722).
Figure 4 Phylogenetic tree of dermatophyte and non-dermatophyte fungi.
In the phylogenetic tree, one of the isolates, A. sublatus (OP132848) has a close relation with Emericella striata (AB248980.1). The genus Emericella is now classified as a subgenus of Aspergillus. Close resemblance between both these strains was determined by the ITS1 5.8S rRNA region genes. Another isolated nucleotide sequence, that is, Aspergillus sydowii isolate ASH-01 (OP120769.1) was connected by nodes in the phylogenetic tree with Aspergillus sp. voucher (MH655008.1) according to the NCBI database. It may or may not be closely related to Aspergillus sydowii because of lack of species identification. Because it is a “voucher” strain, it could be from a separate region of the genus Aspergillus, a unique species, or an unusual strain of an existing species.
The third highlighted strain in the current study was Arthroderma multifidum isolate 1 (OP102681.1), which is directly related to Arthroderma multifidum (AB861736.1). It is indicated by short branch length and direct node joining in the phylogenetic tree. Both strains belong to Arthrodermataceae family and to the same dermatophyte category.24 Arthroderma multifidum (AB861736.1) was submitted by Yamaguchi et al., in 2013.25
The fourth isolated strain from the collected samples was an unidentified fungal species with a taxonomic identification No. OP351722. In the phylogenetic tree, this fungal strain is closely related with another unidentified fungal sp. strain OTU55, which has KT923221.1 nucleotide sequence number in the NCBI database. It was isolated and reported by Clay in 2015 by the Great Lakes Science Center, Cleveland, OH, US from the host Phragmites australis var. australis. Among the identified isolates, A. multifidum, A. sublatus, and A. sydowii were selected for further experiments.
Evolutionary history was inferred using the Neighbor–Joining method.26 The optimal tree is shown in the figure. The percentage of replicate trees in which the associated taxa clustered together in the bootstrap test (500 replicates) is shown next to branches27. Evolutionary distances were computed using the Maximum Composite Likelihood method28 as the number of base substitutions per site. This analysis involved 40 nucleotide sequences. All ambiguous positions were removed for each sequence pair (pairwise deletion option). In all, 713 positions were in the final dataset. Evolutionary analyses were conducted in MEGA11.29
Computational Analysis for Antifungal Susceptibility
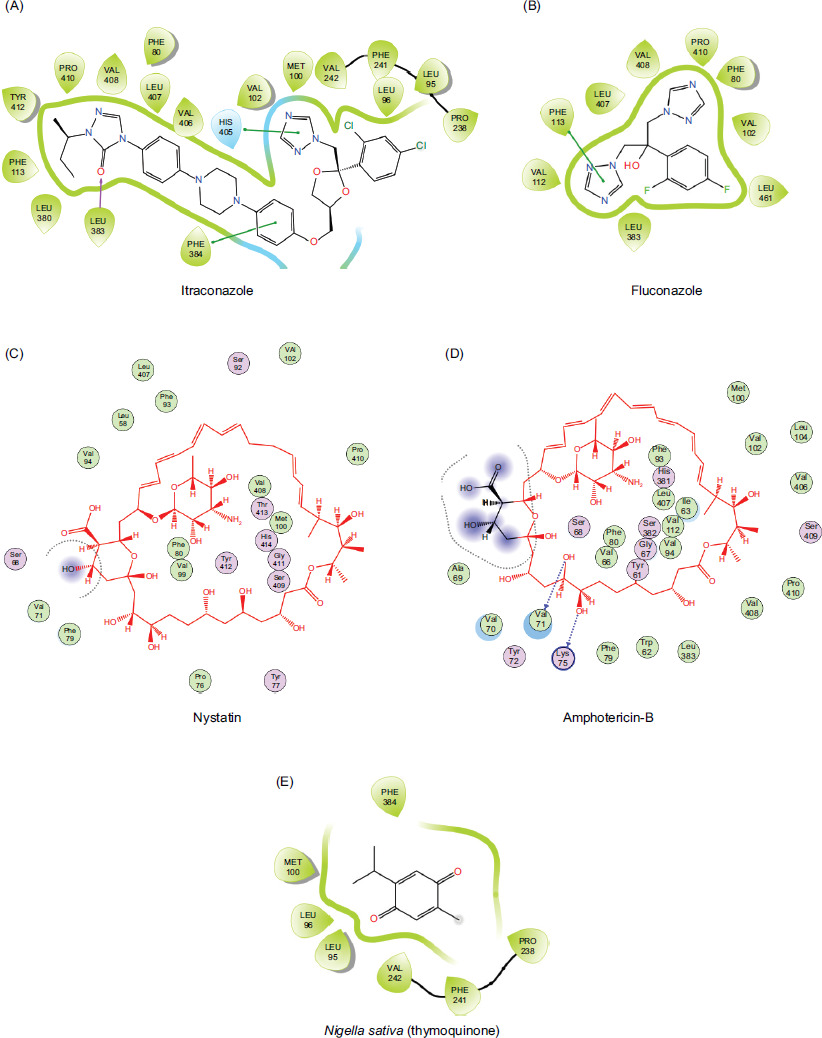
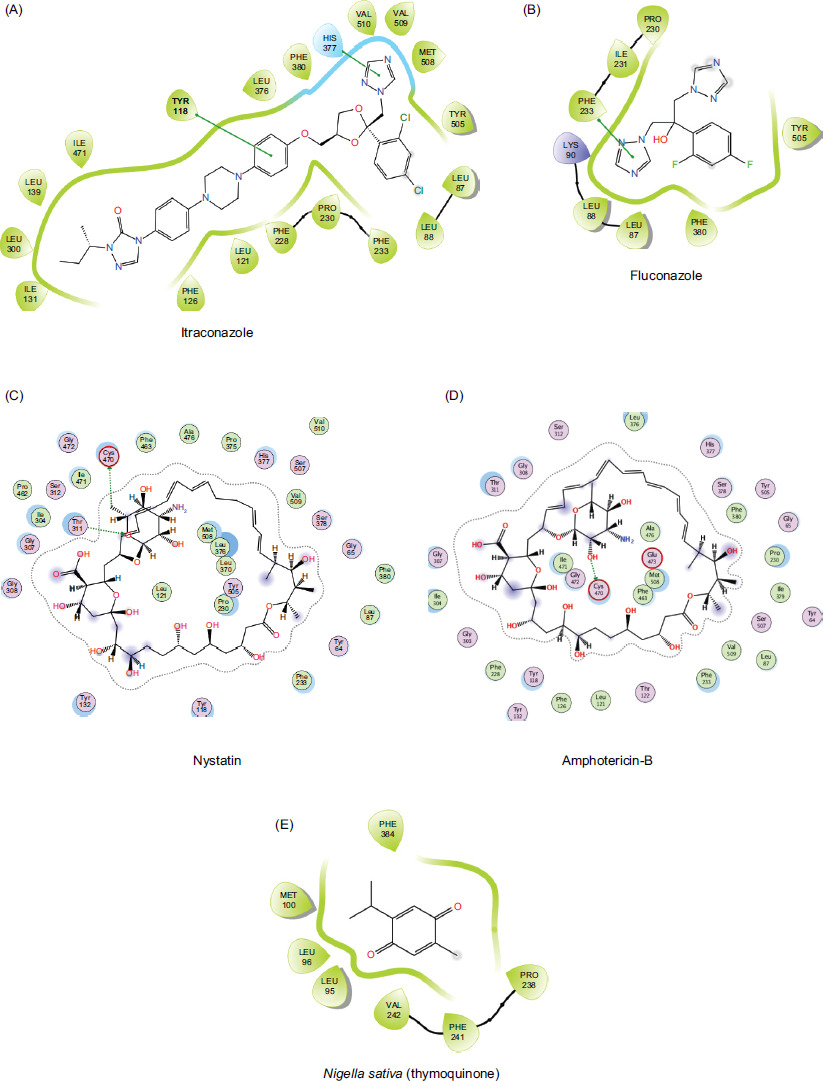
The molecular docking study predicts interactions between two structures (receptor protein and a ligand), using methods such as electrostatic, Van der Waals, Coulombic, and hydrogen bonds. This approach entails determining the energy necessary to bind at various candidate sites. In this study, negative ligand–receptor interaction energy is at molecular targets, demonstrating affinity mainly for standards (fluconazole, itraconazole, nystatin, amphotericin B) and fungi (Aspergillus sydowii). The results are shown in Tables 2 and 3 and Figures 5 and 6.
Table 2 Molecular-operating environment (MOE) docking score and interaction profile (PDB: 5eqb).
| Standard | MOE docking score (kcal/mol) | Interaction residues | |
|---|---|---|---|
| Hydrophobic | Hydrogen bonding | ||
| Itraconazole | −7.93 | Phe80, Leu95, Leu96, Met100, Val102, Phe113, Pro238, Phe241, Val242, Leu380, Leu383, Phe384, His405, Val406, Leu407, Val408, Pro410, Tyr412 | Leu383 |
| Fluconazole | −4.40 | Phe80, Val102, Val112, Phe113, Leu383, Leu407, Val408, Pro410, Leu461 | Phe113, Leu383 |
| Nystatin | −6.56 | Leu58, Val59, Tyr72, Pro76, Phe79, Phe80, Phe93, Val 102, Phe113, Leu407, Val408, Pro410, Tyr412, Thr413, His414, Leu461 | Pro76, Phe80, Met100, Ser409, Tyr412 |
| Amphotericin B | −6.49 | Ile63, Val66, Val70, Val71, Tyr72, Lys75, Pro76, Phe79, Pjhe80, Phe93, Met100, Val 102, Leu104, Val112, Leu407, Val408 | Trp62, Ile63, Val71, Lys75, Phe79, Ser409 |
| Nigella sativa (thymoquinone) | −4.66 | Leu95, Leu96, Met100, Pro238, Phe241, Val242, Phe384 | - |
Table 3 Molecular-operating environment (MOE) docking score and interaction profile (PDB: 5v5z).
| Standard | MOE docking score (kcal/mol) | Interaction residues | |
|---|---|---|---|
| Hydrophobic | Hydrogen bonding | ||
| Itraconazole | -10.89 | Leu88, Leu87, Tyr118, Leu121, Phe126, Ile131, Leu139, Phe228, Pro230, Phe233, Leu300, Leu376, His377, Phe380, Ile471, Tyr505, Met508, Val509, Val510 | - |
| Fluconazole | -5.35 | Leu88, Leu87, Pro230, Ile231, Phe233, Phe380, Tyr505 | Lys90 |
| Nystatin | -6.40 | Tyr64, Leu87, Tyr118, Pro230, Phe233,Thr311, Leu376, Phe380, Ala476, Val509, Val510 | Ilu304, Thr311, Ser378, Pro462, His468 |
| Amphotericin B | -5.09 | Tyr64, Leu87, Tyr118, Thr122, Tyr132, Pro230, Phe233,Thr311, Leu376, Phe380, Ala476, Val509 | Gly303, Thr311, Ser378,Phe380, Cys470 |
| Nigella sativa (thymoquinone) | -5.12 | Tyr118, Pro230, Phe233, Leu376, His377, Ile379, Tyr505, Met508 | His377 |
Figure 5 (A–D) Two-dimensional (2D) interaction of antifungal drugs and (E) Nigella sative compound docked into active region of protein (PDB: 5eqb) showing hydrogen bonding, hydrophobic contacts, and other key molecular interactions.
Figure 6 (A–D) Two-dimensional (2D) interaction of antifungal drugs and (E) Nigella sative compound docked into active region of protein (PDB: 5v5z) showing hydrogen bonding, hydrophobic contacts, and other key molecular interactions.
Antifungal susceptibility profile
A. multifidum showed the highest ZOI against itraconazole (72.00±3.46) and the lowest ZOI of 25.66±2.51 against Amphotericin B (p < 0.05; Table 4). A. sydowii showed the highest ZOI against itraconazole (62.00±3.00) and the lowest ZOI of 22.66±2.51 against fluconazole (p < 0.05; Table 5).
Table 4 Antifungal susceptibility of Arthroderma multifidum.
| Antifungals | ZOI (mm) | Mean±SD | p value | ZOI for dermatophytes (CLSI standard in mm) | ZOI for dermatophytes (EUCAST standard in mm) |
|---|---|---|---|---|---|
| Nystatin | 30 | 31.66±2.88a | 18 | 10-14 | |
| (25 mg/mL) | 35 | ||||
| 30 | |||||
| Itraconazole | 70 | 72.00±3.46b | 18-21 | 15-19 | |
| (2 mg/mL) | 76 | ||||
| 70 | 0.00 | ||||
| Amphotericin B | 23 | 25.66±2.51a | 10-13 | 10-14 | |
| (50 mcg) | 26 | ||||
| 28 | |||||
| Fluconazole | 26 | 30.66±4.50a | 14-17 | 15-19 | |
| (2 mg/mL) | 31 | ||||
| 35 |
Notes: Different superscripts are different significantly, and the same superscripts are different non-significantly.
ZOI: zone of inhibition; CLSI: Clinical and Laboratory Standards Institute; EUCAST: European Committee on Antimicrobial Susceptibility Testing.
Table 5 Antifungal susceptibility of Aspergillus sydowii.
| Antifungals | ZOI (mm) | Mean±SD | p value | ZOI for dermatophytes (CLSI standard in mm) | ZOI for dermatophytes (EUCAST standard in mm) |
|---|---|---|---|---|---|
| Nystatin | 25 | 27.33±2.51a,b | 18 | 10-14 | |
| (25 mg/mL) | 30 | ||||
| 27 | |||||
| Itraconazole | 62 | 62.00±3.00c | 18-21 | 15-19 | |
| (2 mg/mL) | 59 | ||||
| 65 | 0.00 | ||||
| Amphotericin B | 25 | 29.66±4.50b | 10-13 | 10-14 | |
| (50 mcg) | 30 | ||||
| 34 | |||||
| Fluconazole | 23 | 22.66±2.51a | 14-17 | 15-19 | |
| (2 mg/mL) | 25 | ||||
| 20 |
Notes: Different superscripts are different significantly, and the same superscripts are different non-significantly.
ZOI: zone of inhibition; CLSI: Clinical and Laboratory Standards Institute; EUCAST: European Committee on Antimicrobial Susceptibility Testing.
A. sublatus showed the highest ZOI against itraconazole (64.00±1.73) and the lowest ZOI of 26.66±2.08 against Amphotericin B (p < 0.05; Table 6). Furthermore, isolates were selected to check the antimicrobial activity of essential oil of N. sativa against A. multifidum, A. sydowii, and A. sublatus. Essential oil of N. sativa showed activity against all the isolates resistant to antifungals. This essential oil produced the best results, and sensitivity was noted for all isolates o f A. multifidum, A. sydowii, and A. sublatus. Table 7 presents the antifungal activity of essential oil of N. sativa. In the table, A. multifidum shows the ZOI of 24.66±2.51 mm, and ZOI for A. sydowii (31.00±3.46 mm) and A. sublatus (30.00±6.08 mm) are non-significantly different from the ZOI values with the same superscripts (p < 0.05). According to MIC results, the least mean MIC (3.125±0.00 mg/mL) of oil was observed against A. sydowii. Among selected fungi, essential oil of N. sativa was found to be most effective against A. sydowii (Table 8).
Table 6 Antifungal susceptibility of Aspergillus sublatus.
| Antifungals | ZOI (mm) | Mean±SD | p value | ZOI for dermatophytes (CLSI standard in mm) | ZOI for dermatophytes (EUCAST standard in mm) |
|---|---|---|---|---|---|
| Nystatin | 29 | 30.00±1.00a | 18 | 10-14 | |
| (25 mg/mL) | 30 | ||||
| 31 | |||||
| Itraconazole | 65 | 64.00±1.73b | 18-21 | 15-19 | |
| (2 mg/mL) | 62 | ||||
| 65 | 0.00 | ||||
| Amphotericin B | 26 | 26.66±2.08a | 10-13 | 10-14 | |
| (50 mcg) | 25 | ||||
| 29 | |||||
| Fluconazole | 23 | 26.66±4.04a | 14-17 | 15-19 | |
| (2 mg/mL) | 26 | ||||
| 31 |
Notes. Different superscripts are different significantly, and the same superscripts are different non-significantly.
ZOI: zone of inhibition; CLSI: Clinical and Laboratory Standards Institute; EUCAST: European Committee on Antimicrobial Susceptibility Testing.
Table 7 Antifungal susceptibility of essential oil of Nigella sativa.
| Antifungals | ZOI (mm) | Mean±SD | p value |
|---|---|---|---|
| Arthroderma | 27 | 24.66±2.51a,b | |
| multifidum | 25 | ||
| 22 | |||
| Aspergillus | 29 | 31.00±3.46b | |
| sydowii | 35 | 0.029 | |
| 29 | |||
| Aspergillus | 23 | 30.00±6.08b | |
| sublatus | 33 | ||
| 34 |
Different superscripts are different significantly, and the same superscripts are different non-significantly.
ZOI: zone of inhibition
Table 8 Minimum inhibitory concentration (MIC) of Nigella sativa (essential oil) against fungal species.
| Antifungals | MIC (mg/mL) | Mean±SD | p value |
|---|---|---|---|
| Arthroderma | 6.25±0.00 | 6.25±0.00b | |
| multifidum | 6.25±0.00 | ||
| 6.25±0.00 | |||
| Aspergillus | 3.125 | 3.125±0.00c | |
| sydowii | 3.125 | 0.00 | |
| 3.125 | |||
| Aspergillus | 25 | 25.00±0.00a | |
| sublatus | 25 | ||
| 25 |
Different superscripts are different significantly, and the same superscripts are different non-significantly.
Discussion
The cutaneous infections target the skin, hair and nails2 and are recognized as a major health problem, affecting approximately 25% of the global population.30 These infections are equally pathogenic in animals. Diagnosis of cutaneous infections based on symptoms is a challenge, so in developing countries direct microscopic examination and culturing are conventional methods for disease diagnosis. In the current study, 65% of samples were found positive for fungal infections. The results were in contrast to Araya et al. (2020),31 who reported 41.8% positivity in direct microscopy examination. The culture positivity rate in the current study was 23.07 for dermatophyte molds (DMs) and 76.92 for non-dermatophyte molds (NDMs). In contrast to the current study, a comparatively high culture positivity rate for dermatophytes was reported by Araya et al. Previous studies conducted in Addis Ababa, Ethiopia, reported 85.2%, 74.0%, and 42.6% positivity in culture of dermatophytes, compared to the current results.32,33,34 In some studies, 44.8% of culture was reported for the presence of non-dermatophytes.35 These data indicate natural variations in samples for the presence of fungal content. These variations may be associated with differences in sample types, number of samples, host-related factors (immunity/health status), geographical location, climate, and methods used for diagnosis.
The distribution of dermatophytes and etiological agents varies with geographical location. Some species are widely distributed, whereas others are restricted geographically. Most of the fungal infections are transmitted from animals to humans. According to Ridzuan et al. (2021)36, Microsporum canis was the most frequently isolated species, followed by Trichophyton mentagrophytes, and Trichophyton spp. in infected dogs. It is documented that 80–90% of dermatophytosis infections in cats are caused by Microsporum canis. However, studies on new pathogens are also reported.37 In the current study, we isolated A. multifidum from the skin scraps of infected cats. In the United Kingdom, Long et al. (2020)38 reported M. canis and Trichophyton mentagrophytes as the most prevalent species in cats and dogs, in contrast to the current findings. In another study, Moskaluk et al. (2022)39 reported two species, Arthroderma lilyanum and Arthroderma mcgillisianum, as potential pathogens in domestic cats. These results somewhat strengthen the current findings. A. multifidum is a fungus of zoonotic potential, isolated from a 63-year-old immunocompromised patient. It was the first report of human infection.10 Arthroderma is a diverse genus, and it was first isolated in 1963 from rabbit holes and rabbit hair. Previous studies reported its isolation from caves and animals, such as rabbits and horses40. However, there are several risk factors that may contribute to the man-to-man and animal-to-human transmission of dermatophytes. Sharing of clothing, utensils, barbershop material, and dense population play a crucial role in man-to-man transmission.8 On the other hand, rearing of and having a close contact with animals (pets) are reported as a significant risk factor for animal-to-human exposure of dermatophytes.9 The documented evidences highlight the dynamic distribution of emerging fungal pathogens, such as A. multifidum, which may act as a zoonotic pathogen. Therefore, there is a dire need to monitor human and animal reservoirs for better understanding of transmission and prevention of disease.
Similar to other countries, detection of dermatophytes in the Northern and Western parts of Pakistan are reported. Most of the studies described the prevalence of DM species. In a study, 154 samples were collected from different hospitals of Khyber Pakhtunkhwa, Pakistan, and of these, 136 samples were positive. Tinea corporis (35%) was the most predominant type of infection, followed by Tinea capitis (22%). The fungi identified in Tinea corporis infection were Trichophyton rubrum, Trichophyton mentagrophytes, and Epidermophyton floccosum. In a study conducted at University of Health Sciences (UHS) Lahore, Pakistan,42 Tinea corporis was the most common clinical type discovered (27.69%), followed by Tinea capitis (21.53%) and Tinea cruris (12.30%). T. mentagrophytes was the common species isolated (32%), followed by Trichophyton violaceum (28%) and T. rubrum (12%). All studies reported the highest prevalence rate of DMs in contrast to the current results. Among non-dermatophytes, Aspergillus is the most prevalent specie-isolated genus. Results of the current study described similar findings and correlated with the results of Arora et al. (2020)43, Nouripour-Sisakht et al. (2015),44 and Straten et al. (2002)45. A. sydowii is the prevalent species in selected samples of the current study. Aspergillus in the current study is A. sublatus. Both these species were isolated from humans. This indicated that non-dermatophytes are equally important as dermatophytes in cutaneous infections. The present study is the first to report the presence of three fungal isolates A. multifidum (DMs), A. sydowii and A. sublatus (both NDMs) in cutaneous infections. In Pakistan, health facilities are limited, and awareness about infectious diseases, in general, and about fungal infections, in particular, is also limited. Consequently, fungal diseases prevail due to poor hygienic practices and low socioeconomic status8. This study provided awareness about the existence of new fungal pathogens and emphasized the continuous monitoring and surveillance to track the prevalence, spread, and impact on human and animal health.
Phylogenetic analysis based on ITS region of isolated species is performed to construct a phylogenetic tree. By interpreting a tree, we came to know that A. sublatus isolated in the current study closely resembled Emericella nidulans. Both species are non-dermatophytes and belong to the group “Aspergillus section Nidulantes” (formerly Emericella). It is a diverse group of microscopic fungi whose species play essential roles in mycotoxin production, food spoilage, indoor air quality, and human pathogens.46 Emericella nidulans is the second most common mould found in chronic granulomatous disease (CGD) patients, causing nearly invasive infections in this particular host and distinguished by its aggressive behavior.47 Emericella species are commonly detected in soil and decaying plant materials and have a larger morphological diversity than the asexual stages of the Aspergillus genus.48 Emericella is the teleomorph of several Aspergillus species, including Emericella nidulans. It was first reported in China by Matsuzawa et al. in 2006.49
According to the NCBI database, another isolated nucleotide sequence, Aspergillus sydowii isolate ASH-01, is connected with Aspergillus sp. voucher (MH655008.1). Aspergillus sp. voucher was first isolated from Vagator Beach of Goa in India. Both strains are found in the marine environment. The absence of identification of species makes the pathogenic potential uncertain. Some Aspergillus species are opportunistic, especially among immunocompromised patients. Others, on the other hand, are merely environmental and non-pathogenic.50 The third highlighted strain in the current study is Arthroderma multifidum isolate 1, which is directly related to Arthroderma multifidum (AB861736.1). The latter strain was isolated from domestic fowl25 and was published in Mycopathologia in 2014.
The fourth isolated strain from the collected samples is an unidentified fungal species with a taxonomic identification No. OP351722. Its nucleotide sequence resembles another unidentified fungal sp. strain OTU55 (KT923221.1). It was isolated and reported by Clay in 2015 by the Great Lakes Science Center, Cleveland, OH, US from the host Phragmites australis var. australis.
In the Pakistani folk medicine, essential oil of N. sativa is commonly applied topically to treat fungal infections, eczema, acne, and other skin conditions. It is believed that its oil is a staple remedy for skin problems, reflecting deeply rooted cultural practices and a preference for natural treatments. Along with widespread traditional use, scientific research is in progress to fully understand and validate the efficacy and safety of essential oil of Nigella sativa. The efficacy of antifungals was detected initially by molecular docking and later by in-vitro testing. A molecular docking study was conducted to identify the possible mechanism through which each drug (itraconazole, fluconazole, nystatin, amphotericin B, and Nigella sativa) showed potent inhibitory potential of antifungal agent against various fungal infections. Docking of macromolecular proteins (5eqb and 5v5z) indicated significant differences in binding affinities, interacting residues, and interaction patterns, such as hydrophobic and hydrogen bonding as presented in tables 2 and 3 Itraconazole showed the highest score (-7.93 kcal/mol) in all compounds in the case of target protein 5eqb (Figure 1), which is supported by a hydrogen bond with Leu383 and a significant number of hydrophobic residues, such as Phe80, Leu95, Leu96, Met100, Val102, Phe113, Pro238, Phe241, Val242, Leu380, Leu383, Phe384, His405, Val406, Leu407, Val408, Pro410, and Tyr412. With a moderate score of -6.56 kcal/mol, Nystatin acquired the second-highest score in the group because of its hydrophobic associations with Leu58, Val59, Tyr72, Pro76, Phe79, Phe80, Phe93, Val 102, Phe113, Leu407, Val408, Pro410, Tyr412, Thr413, His414, and Leu461. In addition, it formed hydrogen bonds with Pro76, Ser409, and Tyr412. The identical interaction profile of amphotericin B, which had a score of -6.49 kcal/mol, varied in the hydrogen bonds of Trp62, Ile63, Val71, Lys75, Phe79, and Ser409. Among the substances, fluconazole and Nigella sativa had the lowest energy scores and interaction profiles, having respective values of -4.40 kcal/mol and -4.66 kcal/mol. Itraconazole exhibited the highest docking energy of -10.89 kcal/mol against the target protein (5v5z) because of hydrophobic bonding (Leu88, Leu87, Tyr118, Leu121, Phe126, Ile131, Leu139, Phe228, Pro230, Phe233, Leu300, Leu376, His377, Phe380, Ile471, Tyr505, Met508, Val509, and Val510). The effects of fluconazole with Leu88, Leu87, Pro230, Ile231, Phe233, Phe380, and Tyr505 (hydrophobic), and Lys90 (hydrogen) were limited, with a score of -5.35 kcal/mol. In a similar manner, Nigella sativa and amphotericin B showed multiple hydrogen and hydrophobic interactions with lower docking scores (-5.09 kcal/mol and -5.12 kcal/mol). In addition to hydrogen bonding (Thr311 and Ser378) and hydrophobically engaged Tyr64, Phe380, and other residues, nystatin revealed a much less favorable score (-6.40 kcal/mol) than itraconazole and a higher score than other drugs (Figure 2). The knowledge of docking score and interaction profile concluded that itraconazole illustrated the strongest affinity toward both PDB’s (5eqb and 5v5z), indicating the possible mechanism of anti-fungal activity against all the sources by targeting or inhibiting the biosynthesis of lanosterol 14-α-demethylase. This suggests strong antifungal potential of itraconazole against other testing compounds.
Antifungal susceptibility testing has evolved rapidly during the last decade and has now became a relevant tool. To date, studies with molds have concentrated on the selection of optimal in vitro conditions and little attention is paid to correlation with clinical outcome. According to Rex et al. (2001)51, understanding antifungal resistance mechanisms helped to find resistant isolates and validate in vitro measuring techniques. In order to be clinically useful, in vitro methods should predict in vivo response to therapy. There is limited data on in vitro drug resistance in dermatophytes.52 Antifungal resistance, such as antibacterial resistance, is caused by frequent usage or misusage. However, due to limited knowledge, the misuse or overuse of antifungals is not common globally. Ibrahim et al. (2023)53 conducted research on the basic knowledge of antifungals in 2023. The study revealed a substantial gap in physician knowledge and practice when it comes to prescribing antifungal medication. It is one of the reasons that almost all antifungals in the current study are sensitive when compared with the CLSI and EUCAST standards for dermatophyte resistance. Comparison of ZOI in the current study was done with other dermatophytes according to both CLSI and EUCAST guidelines. The species are less reported as non-dermatophyte and dermatophyte fungi from clinical samples, so itraconazole and nystatin could be a better option to treat dermatomycoses caused by isolated fungi. The results of in vitro testing correlate with molecular docking, in which itraconazole and nystatin have the highest binding energy against target molecules. Therefore, both drugs are an effective treatment option for dermatophyte and non-dermatophyte skin infections, as their susceptibility patterns are close to EUCAST and CLSI standards.
Essential oil and various extracts of N. sativa, particularly thymoquinone, have potent antifungal effects on T. mentagrophytes as pathogenic dermatophyte strains.54 Medicinal plants and their extracts have been in use for hundreds of years for treating infectious diseases. Thymoquinone lowered the MIC of standard antimicrobials if used in combination. Thymoquinone also possessed antifungal activity against some very important fungal pathogens, including Candida albicans and certain dermatophytes55. A recent study has demonstrated 100% inhibition for 30 pathogens with a concentration of 1 mg/mL. The MIC test of each compound was studied on eight dermatophytes, five yeast and five mold isolates. Thymoquinone was the best antifungal compound against all of the tested dermatophytes and yeasts, followed by thymol56. Similar to the previous study, the antifungal activity of the ether extract of N. sativa seed and its active principle thymoquinone was tested against dermatophytes species Trichophyton rubrum and one each of T. mentagrophytes. The potentiality of N. sativa as a source for anti-dermatophyte drug support its use in medicine for the treatment of fungal skin infections.57 Above mentioned studies correlates with current findings that N. sativa has antifungal activity and showed active results. Isolates A. multifidum, A. sydowii, and A. sublatus were susceptible to the essential oil of N. sativa and showed inhibitory effects against dermatophyte and non-dermatophyte fungi. Hence, N. sativa is a natural treatment option against cutaneous fungal infections. The current study indicates prominent variations in the distribution of dermatophytes and non-dermatophytes in different samples, while antifungal susceptibility of these fungi remains a favorable option to control cutaneous infections. However, natural remedy, N. sativa emerges as promising therapeutic option.
Conclusions
Cutaneous infections are common in Pakistan. However, lack of awareness and diagnostic facilities remained underdiagnosed. Some fungal diseases even have zoonotic potential. Isolation of non-dermatophytes, A. sydowii and A. sublatus, in our study demonstrates that NDMs are major pathogens in cutaneous infections. In case of animals, zoonotic fungi, A. multifidum, was recovered. This study demonstrated the higher efficacy of itraconazole against dermatophyte and non-dermatophyte fungi in in-silico and in-vitro analyses. In addition, essential oil of N. sativa showed promising results for antifungal activity, particularly against non-dermatophyte A. sydowii. This indicates a cost-effective and safe treatment of cutaneous infections. Although findings of the study are encouraging, large scale studies with increased sample size and geographical area are required to validate results. Overall, this study contributed to the growing understanding of cutaneous fungal infection and emerging fungal pathogens in Pakistan.
Ethical Approval
All experiments were carried out in accordance with relevant guidelines and regulations and all experimental protocols were approved by the Departmental Ethical Review Committee, Institute of Molecular Biology and Biotechnology, University of Lahore, Pakistan.
Consent to Participate
A written consent was signed by all the participants.
Data Availability Statement
All the data generated in this research are included in the manuscript.
Acknowledgments
The authors acknowledged Pet Centre University of Veterinary and Animal Sciences (UVAS), Lahore, for facilitating sample collection from animals and Medical Centre UVAS Lahore, Ravi Campus, for sampling of human hair and nail. The authors extended their appreciation to Princess Nourah bint Abdulrahman University Researchers Supporting Project Number: (PNURSP2026R437), Princess Nourah bint Abdulrahman University, Riyadh, Saudi Arabia.
Author Contributions
Conceptualization: Saba Sana; methodology: Ahsaan Ahmed; software: Najeeb Ullah; validation: Nawal Al-Hoshani; formal analysis: Usman Shehzad and Shaza N. Alkhatib; investigation: Layyaba Nazir; resources: Fakhria A. Al-Joufi; data curation: Hanan Abdulrahman Sagini; writing—original draft preparation: Rukhma; writing—review and editing: Muhammad Nauman Aftab; visualization: Maher S. Alwethaynani; supervision: Saba Sana.; project administration: Saba Sana.
Conflicts of Interests
The authors declared no conflict of interest.
REFERENCES
1 Tahir C, Garbati M, Nggada A, Yawe T, Abubakar M. Primary cutaneous aspergillosis in an immunocompetent patient. J Surg Tech Case Rep. 2011;3(2):94–6.
2 Chanyachailert P, Leeyaphan C, Bunyaratavej S. Cutaneous fungal infections caused by dermatophytes and non-dermatophytes: An updated comprehensive review of epidemiology, clinical presentations, and diagnostic testing. J Fungi. 2023 Jun 20;9(6):669–88.
3 Shah A, Mirza R, Sattar A, Khan Y, Khan SA. Unveiling onychomycosis: Pathogenesis, diagnosis, and innovative treatment strategies. Microb Pathog. 2025 Jan;198(1):107111.
4 Gupta AK, Summerbell RC, Venkataraman M, Quinlan EM. Nondermatophyte mould onychomycosis. J Eur Acad Dermatol Venereol. 2021 Sep;35(9):1628–41.
5 Farwa U, Abbasi SA, Mirza IA, Amjad A, Ikram A, et al. Non-dermatophyte moulds as pathogens of onychomycosis. J Coll Physicians Surg Pak. 2011 Sep;21(9):597–600.
6 Menu E, Filori Q, Dufour JC, Ranque S, L’ollivier C. A repertoire of clinical non-dermatophyte moulds. J Fungi. 2023 Apr 10;9(4):433–502.
7 Smith J. Cutaneous fungal infections. US Pharm. 2015; 40(12):35–9.
8 Soomra H, Shar H, Soomra M. Fungal biota of the domestic animals in a city in Pakistan. Pak J Med Sci. 2010;26(4):964–7.
9 Chang C, Wechtaisong W, Chen Y, Cheng C, Chung S, et al. Prevalence and risk factors of zoonotic dermatophyte infection in pet rabbits in northern Taiwan. J Fungi. 2022 Jun 18;8(6):627–38.
10 Chen Q, Yang Q, Chen H, Yao Y, Shen Y, et al. Zoonotic fungus Arthroderma multifidum causing chronic pulmonary infection. Int J Infect Dis. 2023 Mar 1;130(3):17–9.
11 Puumala E, Fallah S, Robbins N, Cowen LE. Advancements and challenges in antifungal therapeutic development. Clin Microbiol Rev. 2024 Jan 5;37(1):e00142–23.
12 Agu C, Afiukwa A, Orji OU, Ezeh EM, Ofoke IH, Ogbu CO, et al. Molecular docking as a tool for the discovery of molecular targets of nutraceuticals in disease management. Sci Rep. 2023;13(1):13398.
13 Shalaby M, Din N, Hamd A. Isolation, identification, and in vitro antifungal susceptibility testing of dermatophytes from clinical samples at Sohag University Hospital in Egypt. Electron Physician. 2016;8(3):2557–67.
14 Gupta C, Tripathi K, Tiwari S, Rathore Y, Nema S, et al. Current trends of clinicomycological profile of dermatophytosis in Central India. J Am Podiatr Med Assoc. 2014;13(10):23–6.
15 Poluri V, Indugula P, Kondapaneni L. Clinicomycological study of dermatophytosis in South India. J Lab Physicians. 2015;7(2):84–9.
16 Gnat S, Lagowski D, Nowakiewicz A, Dyląg M, Osińska M, et al. Detection and identification of dermatophytes based on currently available methods—A comparative study. J Appl Microbiol. 2021;130(1):278–91.
17 Dalis J, Kazeem H, Kwaga J, Kwanashie C, Yakubu B, et al. Molecular characterization of dermatophytes isolated from cattle in Plateau State, Nigeria. Vet Microbiol. 2018;219(1):212–8.
18 Halgren TA. MMFF VI. MMFF94s option for energy minimization studies. J Comput Chem. 1999;20(7):720–9.
19 Monk BC, Tomasiak M, Keniya V, Huschmann FU, Tyndall JD, O’Connell JD, et al. Architecture of a single membrane spanning cytochrome P450 suggests constraints that orient the catalytic domain relative to a bilayer. Proc Natl Acad Sci U S A. 2014;111(10):3865–70.
20 Phienluphon A, Kondo K, Mikami B, Nagata T, Katahira M. Structural insights into the molecular mechanisms of substrate recognition and hydrolysis by feruloyl esterase from Aspergillus sydowii. Int J Biol Macromol. 2023;253(1):127188.
21 Aala F, Yusuf UM, Khodavandi A, Jamal F. In vitro antifungal activity of allicin alone and in combination with two medications against six dermatophytic fungi. Afr J Microbiol Res. 2010;4(4):380–5.
22 Prajapati S, Sharma M, Kumar A, Gupta P, Dwivedi B, et al. Antimicrobial activity of different homoeopathic drugs and their potencies against Aspergillus niger in vitro. Indian J Res Homoeopathy. 2019;13(3):150–8.
23 Pfaller A, Castanheira M, Diekema J, Messer A, Jones N. Triazole and echinocandin MIC distributions with epidemiological cutoff values for differentiation of wild-type strains from non-wild-type strains of six uncommon species of Candida. J Clin Microbiol. 2011;49(10):3800–4.
24 Taxonomy. Taxonomy (Arthrodermataceae) [Internet]. National Center for Biotechnology Information. [cited n.d.]. Available from: https://www.ncbi.nlm.nih.gov/Taxonomy/Browser/wwwtax.cgi?id=34384
25 Yamaguchi S, Sano A, Hiruma M, Murata M, Kaneshima T, Murata Y, et al. Isolation of dermatophytes and related species from domestic fowl (Gallus gallus domesticus). Mycopathologia. 2014;178(1–2):135–43.
26 Saitou N, Nei M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol Biol Evol. 1987;4(4):406–25.
27 Felsenstein J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution. 1985;39(4):783–91.
28 Kumar S, Tamura K, Nei M. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc Natl Acad Sci U S A. 2004;101(30):11030–5.
29 Tamura K, Stecher G, Kumar S. MEGA 11: Molecular evolutionary genetics analysis version 11. Mol Biol Evol. 2021 Jun 25;38(7):3022–7.
30 Martinez-Rossi NM, Peres NTA, Bitencourt TA, Martins MP, Rossi A. State-of-the-art dermatophyte infections: Epidemiological aspects, pathophysiology, and resistance mechanisms. J Fungi. 2021;7(8):629–46.
31 Araya S, Tesfaye B, Fente D. Epidemiology of dermatophyte and non-dermatophyte fungi infection in Ethiopia. Clin Cosmet Investig Dermatol. 2020;13(1):291–7.
32 Teklebirhan G, Bitew A. Prevalence of dermatophytic infection and the spectrum of dermatophytes in patients attending a tertiary hospital in Addis Ababa, Ethiopia. Int J Microbiol. 2015;2015(1):1–5.
33 Woldeamanuel Y, Leekassa R, Chryssanthou E. Clinico-mycological profile of dermatophytosis in a reference centre for leprosy and dermatological diseases in Addis Ababa. Mycopathologia. 2006;161(3):167–72.
34 Fentaw S, Gentachew T, Assefa M. A five-year retrospective study of dermatophytosis and dermatomycoses at the Mycology Referral Laboratory of EHNRI, Addis Ababa, Ethiopia. Cienc Biol. 2010;41(1):1–7.
35 Bitew A. Dermatophytosis: Prevalence of dermatophytes and non-dermatophyte fungi from patients attending Arsho Advanced Medical Laboratory, Addis Ababa, Ethiopia. Dermatol Res Pract. 2018;2018:1–6.
36 Ridzuan M, Ruth M, Arffah K. Isolation of dermatophytes from infected stray cats in Selangor. J Berkala Epidemiol. 2021;9:231–8.
37 Smagulova AM, Kukhar YV, Glotova TI, Glotov AG, Kim AS. First record of Trichophyton benhamiae isolated from domestic cats in Russia. Med Mycol Case Rep. 2023;40:16–21.
38 Long S, Carveth H, Chang YM, O’Neill D, Bond R. Isolation of dermatophytes from dogs and cats in the south of England between 1991 and 2017. Vet Rec. 2020;187:e87.
39 Moskaluk A, Woude SV. Two novel species of Arthroderma isolated from domestic cats with dermatophytosis in the United States. Med Mycol. 2022;60:1–10.
40 Dawson CO. Two new species of Arthroderma isolated from soil from rabbit burrows. Sabouraudia. 1963;3:185–91.
41 Ali-Shtayeh MS, Salameh AA, Abu-Ghdeib SI, Jamous RM, Khraim H. Prevalence of tinea capitis as well as of asymptomatic carriers in school children in Nablus area (Palestine). Mycoses. 2002;45:188–94.
42 Shakir S, Saleem S, Rizvi W, Waheed A, Iqbal J. Isolation, identification and antifungal susceptibility of dermatophytes isolated from clinically suspected cases of tinea infections in Pakistan. Microbiol Res J Int. 2019;29:1–11.
43 Arora T, Oberoi L, Malhotra A, Kauri R. Mycological pattern of dermatophytes and non-dermatophytes in a tertiary care hospital. Int J Health Sci Res. 2020;10:37–41.
44 Nouripour-Sisakht S, Mirhendi H, Shidfar MR, Ahmadi B, Rezaei-Matehkolaei A, et al. Aspergillus species as emerging causative agents of onychomycosis. J Mycol Med. 2015;25:101–7.
45 Van Straten MR, Balkis MM, Ghannoun MA. The role of non-dermatophyte molds in onychomycosis: diagnosis and treatment. Dermatol Ther. 2002;15:89–95.
46 Hubka V, Nováková A, Peterson SW, Frisvad JC, Sklenář F, Matsuzawa T, et al. A reappraisal of Aspergillus section Nidulantes with descriptions of two new sterigmatocystin-producing species. Plant Syst Evol. 2016;302(9):1267–99.
47 Henriet SS, Verweij PE, Warris A. Aspergillus nidulans and chronic granulomatous disease: a unique host–pathogen interaction. J Infect Dis. 2012;206(7):1128–37.
48 Abdel-Azeem A, Salem F, Abdel-Azeem M, Nafady N, Mohesien M, Soliman E. Biodiversity of the genus Aspergillus in different habitats. In: Gupta VK, editor. New and Future Developments in Microbial Biotechnology and Bioengineering. Amsterdam: Elsevier; 2016. p. 3–28.
49 Schoch CL, Ciufo S, Domrachev M, Hotton CL, Kannan S, Khovanskaya R, et al. NCBI taxonomy: a comprehensive update on curation, resources and tools. Database (Oxford). 2020;2020:baaa062.
50 Mousavi B, Hedayati MT, Hedayati N, Ilkit M, Syedmousavi S. Aspergillus species in indoor environments and their possible occupational and public health hazards. Curr Med Mycol. 2016;2(1):36–42.
51 Rex JH, Pfaller MA, Walsh TJ, Chaturvedi V, Espinel-Ingroff A, Ghannoum MA, et al. Antifungal susceptibility testing: practical aspects and current challenges. Clin Microbiol Rev. 2001;14:643–58.
52 Rudramurthy S, Dogra S, Shaw D. Antifungal drug susceptibility testing of dermatophytes: laboratory findings to clinical implications. Indian Dermatol Online J. 2019;10:225.
53 Ibrahim SM, Adlan N, Alomair SM, Butaiban I, Alsalman A, Bawazeer A, et al. Evaluation of systemic antifungal prescribing knowledge and practice in the critical care setting among ICU physicians and clinical pharmacists: a cross-sectional study. Antibiotics. 2023;12:238.
54 Mahmoudvand H, Sepahvand A, Jahanbakhsh S, Ezatpour B, Mousavi SA. Evaluation of antifungal activities of the essential oil and various extracts of Nigella sativa and its main component, thymoquinone, against pathogenic dermatophyte strains. J Mycol Med. 2014;24:155–61.
55 Nouri N, Mohammadi SR, Beardsley J, Aslani P, Ghaffarifar F, et al. Thymoquinone antifungal activity against Candida glabrata oral isolates from patients in intensive care units—an in vitro study. Metabolites. 2023;13:580–92.
56 Taha M, Azeiz AZ, Saudi W. Antifungal effect of thymol, thymoquinone and thymohydroquinone against yeasts, dermatophytes and non-dermatophyte molds isolated from skin and nail fungal infections. Egypt J Biochem Mol Biol. 2010;28:109–26.
57 Aljabre SH, Randhawa MA, Akhtar N, Alakloby OM, Alqurashi AM, et al. Antidermatophyte activity of ether extract of Nigella sativa and its active principle, thymoquinone. J Ethnopharmacol. 2005;101:116–19.