
Download



REVIEW ARTICLE
Mechanisms and implications of histamine-induced reactions and complications
Ahmed Al-maamari†, Marwa Sultan†, Shanshan Ding, Duan Yuxin, Meng-Yao Wang, Suwen Su*
The Key Laboratory of Neural and Vascular Biology, Ministry of Education, The Key Laboratory of New Drug Pharmacology and Toxicology, Department of Pharmacology, Hebei Medical University, Shijiazhuang, P.R. China
†Ahmed Al-maamari and Marwa Sultan contributed equally to this work
Abstract
Histamine, classified as a biogenic amine, plays a crucial role in both pro-inflammatory and immune regulatory processes, thereby establishing itself as a key mediator in allergic diseases and immune responses. This review provides an exhaustive analysis of the structure, function, and regulation of histamine, with particular emphasis on its interaction with four receptor subtypes: histamine H1 receptor (H1R), histamine H2 receptor (H2R), histamine H3 receptor (H3R), and histamine H4 receptor (H4R), all of which are instrumental in mediating a variety of physiological processes, including neurotransmitter release, modulation of immune responses, and gastric acid secretion. The review explores intracellular signaling pathways mediated by the activation of these receptors, highlighting the complex cascades involved in immediate- and delayed-type hypersensitivity reactions. It also examines the broad spectrum of histamine-induced complications, focusing on their effects on the gastrointestinal, cardiovascular, respiratory, and central nervous systems, and emphasizes histamine’s potential to cause vascular dysfunction and other pathological changes. Furthermore, the role of histamine in inflammation and immune responses is explored, particularly in the context of allergic diseases such as asthma, allergic rhinitis, and atopic dermatitis. The review also covers pharmacological interventions targeting histamine receptors, including the use of antihistamines and mast cell stabilizers, which are critical for the treatment of symptoms and the inhibition of the progression of histamine-related conditions. Finally, the review addresses emerging research and future directions, identifying potential areas for innovation and improved therapeutic strategies. This comprehensive overview not only deepens understanding of histamine’s multifaceted roles in health and disease, but also underscores the importance of developing advanced diagnostic tools and targeted treatments for histamine-associated disorders.
Key words: allergic diseases, cardiovascular complications, histamine intolerance, histamine receptors, hypersensitivity, immune modulation, inflammation, mast cells
*Corresponding author: Suwen Su, Department of Pharmacology, Hebei Medical University, Shijiazhuang, Hebei 050017, P.R. China. Email address: [email protected]
Received 25 November 2024; Accepted 23 January 2025; Available online 1 May 2025
Copyright: Al-maamari A, et al.
License: This open access article is licensed under Creative Commons Attribution 4.0 International (CC BY 4.0). http://creativecommons.org/licenses/by/4.0/
Introduction
Histamine, an important biogenic amine, plays an essential role in the immune system’s defense mechanisms against external antigens by initiating a range of physiological responses, including vasodilation, increased vascular permeability, and the modulation of immune cell functions.1,2 Histamine is secreted primarily by basophils and mast cells when stimulated by immunoglobulin E (IgE) through high-affinity Fc receptors.3,4 The release of histamine stimulates four distinct receptors, namely H1R, H2R, H3R, and H4R, each fulfilling a unique function in receptor localization, signal transduction, and deactivation processes. Upon binding to a ligand, these receptors engage heterotrimeric guanine nucleotide-binding proteins (G-proteins), which subsequently initiate a cascade of intracellular signaling events, including the activation of protein kinases and second messenger-generating enzymes.5,6 Phosphorylation and dephosphorylation of multiple sites on the plasma membrane effectively modulate the strength, duration, and spatial extent of receptor signaling.7,8 The activity of phosphodiesterase, enzymes that metabolize cyclic AMP (cAMP) and cyclic GMP (cGMP), further shapes the cellular response to histamine.9 Complications arising from histamine release, regardless of whether it is endogenous or exogenous in origin, can fluctuate in severity, ranging from mild cases to conditions that may pose a significant threat to life. Severe manifestations include systemic anaphylaxis and anaphylactic shock, characterized by symptoms such as edema, hypotension, and hypoxemia.10,11 On the milder end of the spectrum, histamine can cause pseudoallergic reactions, commonly referred to as histamine intolerance, often triggered by certain foods. Pharmacological treatment of histamine-induced conditions generally involves inhibition of histamine synthesis or blocking its receptors, with a particular emphasis on the H1R target.12,13 The hypothesis underlying this approach suggests that a singular trigger initiates a signaling cascade. However, recent evidence implies that histamine’s actions on immune cells, especially those involved in mast cell and basophil-mediated responses, are more varied and intricate than previously perceived. This variability in response highlights the complexity of histamine function within the immune system and its ability to elicit a wide spectrum of pro-inflammatory reactions.14,15
Histamine: Structure, Function, and Regulation
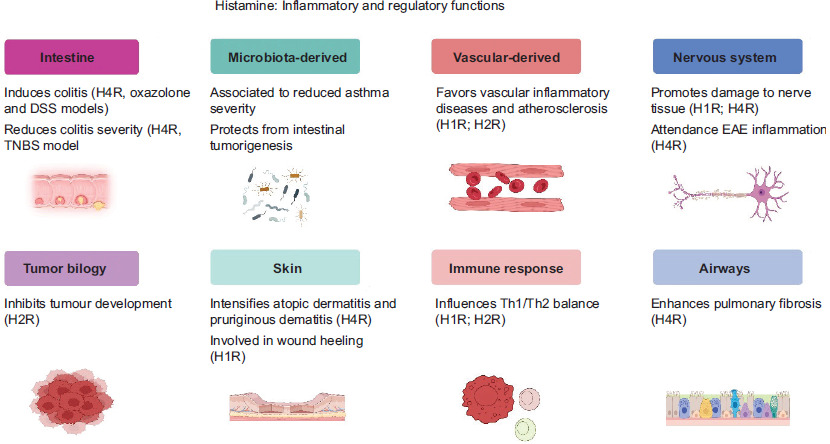
Histamine is a bioactive molecule with an approximate molecular weight of 111 kDa. It is characterized by the presence of an imidazole ring linked to an ethylamine moiety. Histamine biosynthesis involves the decarboxylation of the amino acid histidine, which is facilitated by the enzyme histidine decarboxylase, a mechanistic detail that was elucidated in the 1950s.16 Histamine is stored primarily in granules within mast cells, a discovery made in the 1960s. These cells are predominantly located in connective tissues and mucosal surfaces, where they play a crucial role in immune responses. In addition to mast cells, histamine is also present in nerve cells, lymphocytes, and epithelial cells, where it is sequestered in secretory granules.17 Histamine exerts effects on multiple physiological systems and processes (Figure 1). It influences the nervous system by contributing to the perception of pain and modulating neurotransmitter activity. In the cardiovascular system, histamine facilitates vasodilation and improves vascular permeability, thus playing a critical role in allergic reactions.18 Furthermore, within the gastrointestinal system, histamine stimulates the secretion of gastric acid, induces bronchoconstriction in the respiratory system, and also affects the genitourinary system.19
Figure 1 Histamine plays both inflammatory and regulatory roles across various biological systems.
Furthermore, histamine is crucial in immune responses, being involved in acute allergic reactions and chronic inflammation.1,20 Elevated histamine levels can lead to adverse reactions, such as trauma or necrosis caused by histamine poisoning. Histamine intolerance or hypersensitivity, which affects a small percentage of the population, can lead to severe reactions. Histamine is also known to promote endothelial-mesenchymal transitions, reduce fibrotic scar tissue, and enhance aesthetic skin recovery. Mast cells can secrete sufficient amounts of histamine in the case of fibrotic dermal lesions, where it can combine with other angiogenic molecules to promote adequate neovascularization in the skin, similar to mast cells in the nonfibrotic dermis.21,22
The magnitude of symptoms and adverse outcomes depends on the concentration of histamine, the duration of exposure, and the pathway of entry into the organism. Pathological conditions, such as breast carcinoma and atrophic gastritis, are correlated with modified histamine levels, thereby affecting clinical outcomes.23 In summary, histamine is a vital mediator with a wide range of effects, from regulating immune responses to influencing various organ systems. Its regulation is complex, and when it is dysregulated, it can cause significant health problems.24
Histamine receptor subtypes and their roles
Histamine exerts its effects through four different types of histamine receptors, all of which belong to the G protein-coupled receptor (GPCR) family and are characterized by unique pharmacological properties and specific amino acid sequences.25 These receptors, H1R, H2R, H3R, and H4R, mediate various physiological and pathophysiological processes, including neurotransmitter release, regulation of cell functions, organ perfusion, and modulation of inflammatory responses (Figure 2).5 In addition, histamine influences processes such as pain perception, hormone secretion, cytokine synthesis, and tissue repair.26
Figure 2 Histamine receptors and their roles in various physiological and pathological conditions.
H1 Receptors
H1R plays a crucial role in mediating allergic reactions and inflammatory responses. Upon activation, H1R is associated with G proteins, specifically Gq/11, which subsequently initiate the activation of phospholipase C beta (PLCβ). This enzyme catalyzes the hydrolysis of the membrane phospholipid phosphatidylinositol 4,5-bisphosphate (PIP2), resulting in the production of two important second messengers: inositol trisphosphate (IP3) and diacylglycerol (DAG). Inositol trisphosphate (IP3) then binds to its receptors located in the endoplasmic reticulum, prompting the release of calcium ions into the cytoplasm. The subsequent elevation of intracellular calcium concentration triggers a series of cellular events mediated by calcium-binding proteins such as calmodulin, calcineurin, and calcium/calmodulin-dependent kinases. This pathway highlights the essential role of H1R in orchestrating the immune system’s response to allergens and inflammation.27,28
H2 Receptors
H2R is primarily responsible for regulating gastric acid secretion and plays a significant role in modulating cardiac function. H2R transmits signals primarily through Gs proteins, which stimulate adenylyl cyclase, thereby increasing cAMP levels and leading to the activation of protein kinase A (PKA). Furthermore, H2R is capable of signaling through G protein-independent pathways, such as phosphorylation of its C-terminal tail, thereby enhancing its functional versatility in physiological processes.29,30
H3 Receptors
H3R is predominantly expressed within the central nervous system, where it serves as both an autoreceptor and a heteroreceptor, modulating histamine release along with other neurotransmitters. Upon activation, H3R is associated with Gi/o proteins, resulting in the inhibition of adenylyl cyclase, a decrease in cAMP levels, and subsequent regulation of neurotransmitter release. This inhibitory signaling mechanism underscores the critical role of H3R in maintaining neurotransmitter equilibrium within the central nervous system.6,31
H4 Receptors
H4R is predominantly expressed in hematopoietic cells, where it plays a crucial role in modulating immune responses. Similar to H3R, H4R is associated with Gi/o proteins, which function to inhibit the production of cAMP while simultaneously regulating calcium mobilization and cell migration.32,33 Beyond these widely researched functions, H4R is an essential component in various physiological processes. Histamine is crucial for the protection of the gastrointestinal tract by facilitating mucosal lining renewal, promoting tissue repair, modulating gastric acid secretion, and improving nutrient absorption. Furthermore, histamine plays an active role in the regulation of the wake-sleep cycle, fluid homeostasis, and lipid metabolism.34,35 The various biological effects of histamine are mediated through distinct receptor-driven signaling pathways that can propagate or inhibit cellular responses, depending on the subtype of the receptor and the cellular context.36 A deeper understanding of these pathways is essential for the development of targeted therapies for histamine-related disorders.
Intracellular Signaling Cascades
Complex intracellular signaling pathways initiated by histamine receptors exhibit considerable variability depending on the specific activated subtype (Figure 3), with H1R and H2R being particularly important. H1R is intricately linked to multiple Gq, G11, and G12 proteins, thereby activating the phospholipase C (PLC)/Ca+ mobilization pathway and promoting phosphoinositide hydrolysis upon histamine binding. This robust and multifaceted activation leads to the production of DAG and IP3, which subsequently results in the efficient release of a large amount of calcium ions from the cell and facilitates the activation of protein kinase C (PKC).24,37 In contrast, H2R is coupled synergistically with various proteins to stimulate adenylyl cyclase (AC), leading to a significant increase in cAMP levels. This elevation in cAMP ultimately serves as a powerful activator of protein kinase A (PKA), further accentuating its profound impact.38 Despite their distinct characteristics, the intricate signaling pathways of histamine receptors collectively reveal the multifaceted tissue-specific effects of histamine activation. This complexity underscores the broad influence of histamine on both physiological and pathological processes, highlighting the importance of individual receptor subtypes and their diverse associated signaling pathways in shaping the overall response.39,40
Figure 3 Histamine receptors and their intracellular signaling pathways.
Mechanisms of Action
Histamine exerts effects on target tissues mainly through the modulation of intracellular signaling pathways via receptors that belong to the G-protein-coupled receptor family. To date, four types of histamine receptors have been described: H1R, H2R, H3R, and H4R. All receptors exert pro-inflammatory effects when activated in target cells. However, the net biological action of histamine depends on the activated histamine receptor subtype, the composition of target cells, and the local environment of cytokines and chemokines.41,42 H1R is responsible for mediating an increase in vascular permeability, contraction of smooth muscle tissues, and stimulation of mucous secretions. Upon binding to histamine, activated H1R interacts with Gq/11 proteins, resulting in the activation of phospholipase Cβ, which subsequently generates inositol trisphosphate and diacylglycerol. This sequence leads to the release of intracellular calcium and the activation of calcium-dependent protein kinases, triggering a range of functions, from the contraction of vascular endothelium and smooth muscle cells of the bronchial to the activation of phospholipase A2, culminating in the production of inflammatory mediators such as prostaglandins and leukotrienes.43,44 Upon activation, H1R triggers the activation of small RhoA GTPase, protein kinase C, and mitogen-activated protein kinases, which in turn facilitate the expression of genes that encode endothelial adhesion molecules and cytokines. The increase in these molecules enhances the adherence and transendothelial migration of neutrophils and monocytes to inflammatory sites. The pro-inflammatory characteristic of histamine is demonstrated through its ability to induce the expression of genes for pro-inflammatory cytokines, including IL-6, IL-8, and TNF-α, in human endothelial cells and various other cell types.45,46 The activation of H3R, in addition to an autoinhibitory action in histaminergic cells, leads to anti-inflammatory effects, including down-regulation of the release of cytokines and chemokines by peripheral glial cells and neurons in the central nervous system. H3R inhibits the activation of the Gq/11 protein involved in the transduction of the response by H1R, leading to a decrease in intracellular calcium levels secondary to a reduction in phospholipase C activation, resulting in decreased vascular permeability. Activated H3R also inhibits the expression of adhesion molecules in endothelial cells, limiting the adhesion and transendothelial migration of monocytes and neutrophils to tissues. In other cell types, such as T lymphocytes and macrophages, H3R activation leads to the downregulation of Th2-type cytokine release and/or upregulation of Th1-type cytokines, contributing to the accentuation of immune Th1-type responses. This H3R-mediated Th1 skew effect provides a mechanism through which histamine can negatively control allergic responses.47,48
Histamine-Induced Reactions in the Body
Histamine is a vital mediator in the body and contributes significantly to a variety of histamine-induced reactions, including immediate and delayed-type hypersensitivity reactions. These reactions do not arise from a single trigger factor; instead, they can be triggered by various stimuli that cause the immune system to produce antibodies against histamine or its receptors.49 This can result in cross-reactions between different, structurally unrelated molecules and tissues, complicating the clinical picture and making accurate diagnosis difficult. Clinically, histamine-induced reactions can present as a variety of conditions, such as anaphylactic episodes, vasculitis syndromes, familial Mediterranean fever, inflammatory bowel disease, interstitial cystitis, and various autoinflammatory syndromes.20 The diversity in clinical and biological presentations is due to the complex regulatory role of histamine in the immune system.2 Understanding these mechanisms is essential to improve medical treatments and therapies, as well as to increase awareness of the potential dangers associated with histamine, especially as a food ingredient, and its effects on individuals with allergies, autoimmune diseases, autoinflammatory conditions, and histamine intolerance. This knowledge is crucial for better managing and preventing the wide range of symptoms and complications that can arise from histamine exposure.
Immediate-Hypersensitivity Reactions
Immediate hypersensitivity reactions are rapid and often exaggerated immune responses mediated by the release of significant amounts of histamine and other pro-inflammatory mediators from mast cells. They can lead to a wide variety of localized or systemic symptoms and, when severe, can result in recurrent episodes of anaphylaxis and even death. A solid understanding of the molecular and cellular events that underlie immediate hypersensitivity reactions is essential for our ability to recognize, manage, and prevent severe allergic reactions and anaphylaxis.50,51 Although the immune response is crucial for defending the host against pathogens and promoting tissue repair, it can sometimes result in severe, life-threatening, or even fatal outcomes. Despite its protective role, the release of excessive amounts of histamine and other pro-inflammatory mediators from immune cells can lead to a variety of adverse effects.13,52 When these side effects become severe, they are referred to as severe allergic reactions or chronic allergic diseases. When they are acute and systemic, they are commonly called anaphylaxis. Histamine triggers many of these effects and, as a result, was identified as the most important bactericidal factor in the immune response during the early years of the twentieth century.53
Histamine is most commonly known for its effects on three physiological systems: blood vessels (vascular permeability changes), especially in the post-capillary venule, which allows fluids, macromolecules, and leukocytes to leave the blood and enter the tissues; smooth muscle (smooth muscle contraction); and the heart (increased heart rate, if circulating concentrations are high).43,54 Histamine can also trigger changes in mucus secretions and ion movement, which are responsible for some or all symptoms of rhinorrhea (runny nose), congestion, sneezing, and nasal itching. In the intestine, it can trigger peristalsis or cause diarrhea through secretory changes.55 Additionally, histamine can stimulate and attract eosinophils, neutrophils, and T lymphocytes.30 Histamine and other mediators and cytokines of local inflammation can control the magnitude and duration of an acute allergic reaction or a local response. For example, people who can tolerate foods containing tyramine but take a type of drug that inhibits monoamine oxidase, an enzyme that breaks down histamine, may experience an acute hypersensitivity reaction if they consume foods that the body normally breaks down quickly or while the drugs are in their bloodstream.56 When histamine binds to H1R receptors in the brain, it can trigger sleepiness and other sensations. In rodents, histamine has been identified as a neurotransmitter in the brain, where small amounts of it can be stored, but the effect of histamine as a neurotransmitter in humans is uncertain.57 Histamine can also affect the processing of neurological signals related to the skin and intestine. It can contribute to itching, causing the patient to scratch excessively, which may prolong the inflammatory response and other symptoms.5,58
Delayed-Type Hypersensitivity Reactions
Some hypersensitivity reactions described above, such as the classic Pimms reaction and type I hypersensitivity reactions, may not be due to histamine release in human airways,59 but rather to the release of nonhistaminergic mediators, very high concentrations of mediators, or nonspecific stimuli of bronchial smooth muscle. These reactions and clinical symptoms may contribute to the difficulty in understanding bronchial nonspecific hyperreactivity. Furthermore, salmon proteins, ovalbumin, or toluene diisocyanate may lack the special (non-contractile) stimulus or non-concentration-dependent reactivity necessary to trigger histamine release from human cells.60,61 The onset of antigen-induced activation, proliferation, differentiation, and accumulation of the peptide-hapten conjugate characterizes the ensuing immune response. This involves the activation of helper T cells, the production of cytokines, and tight regulation over cytokine production and cell apoptosis.57 Helper T cells direct the development of peptide-hapten-specific cytotoxic T cells, foreign body macrophages, eosinophils, and neutrophils. Inflammation exhibits the hallmarks of a classic delayed-type hypersensitivity reaction, with limited erythema but severe recruited inflammation, endothelial degranulation, and the expression of adhesion molecules.62,63 Understanding the mechanisms of lymphocyte type IV collagen-mediated injury is crucial, as previously noted, in the general approach to delayed-type hypersensitivity and in the development of antilymphocyte therapies for autoimmune and chronic inflammatory diseases.
Histamine-Induced Complications
Histamine can also cause or be involved in various complications. It induces gastric acid secretion in humans and can lead to a decrease in heart rate in cases of acute histamine poisoning. Additionally, individuals sensitive to histamine may experience adverse reactions. When histamine concentration reaches 20 μg/kg or higher, adverse reactions are more common.64 Acute administration of histamine to humans, either intradermally or intravenously, does not typically cause significant clinical effects. However, a profound hypotensive and anticonvulsant response has been documented in a large clinical series, suggesting the presence of histamine. This phenomenon has mainly been observed in dermatology. Therefore, it is reasonable to predict that most of the clinical activity of histamine stems from the release of endogenous stores, which can provoke potentially serious adverse reactions.13,65 Regulating the release of histamine is essential, and the liberal use of antagonists, particularly H2R antagonists, plays a significant role in managing its effects. Histamine can produce a range of responses across nearly all human organ systems, presenting potential therapeutic challenges. H1R is widely distributed and plays a role in brain edema formation, respiratory bronchoconstriction, and vascular spasm. Both H1R and H2R receptors are found in mucosal mast cells, which can release a variety of active mediators.66,67 A detailed understanding of the different pathological effects of histamine is necessary for those managing histamine-associated disorders, in order to provide accurate laboratory diagnoses and appropriate treatment for histamine-induced adverse effects in various human organ systems.
Allergic Rhinitis Complications
Histamine, a potent biological amine mediator of inflammation, is found in certain cells. It is considered the main pathogenic mediator in the development of the classic symptoms of allergic rhinitis, including nasal congestion, rhinorrhea, and itching, particularly during allergic reactions. Studies have reported increases in nasal sodium, mast cells, basophils, polymorphonuclear leukocytes, platelets, and plasma histamine concentrations in patients with allergic rhinitis. Histamine permeability also increases, and allergic rhinitis patients are likely to encounter significantly higher histamine concentrations during episodic flare-ups.52,68,69 Triggers of the human sneeze reflex include histamine and cold air. Several mechanisms contribute to nasal paralysis and rhinorrhea. Histamine can induce the release of neurotrophic proteins, such as chemokines, cytokines, neurotransmitters, neuropeptides, and growth factors from the nasal mucosa and immune cells. Compound damage or symptomatic drugs can inhibit the release of these chemicals, preventing them from binding to receptors, thereby suppressing the sensitization or stimulation of afferent sensory neurons and the discharge of neurotransmitters. For instance, tachykinins like substance P, bradykinin, or calcitonin gene-related peptide can cause sneezing and rhinorrhea. In allergic rhinitis, sensory neurons in the nasal mucosa are hyperresponsive, further contributing to symptoms.70–72 In vitro studies of human nasal mucosa reveal that V1 receptors in sensory neurons release histamine when stimulated. Additionally, these neurons synthesize intracellular proteins like c-fos kinase and gene product 9.5, which play roles in the neuro-inflammatory process. These findings have significant implications for otolaryngologists, allergists, and patients with allergic rhinitis symptoms.55,73
Complications of Asthma Exacerbation
The role of histamine in the pathophysiology of asthma is prominent. Histamine has been shown to cause the contraction of human airways in vivo, and intravenous injection of histamine can lead to significant increases in airway resistance (Raw) and a reciprocal decrease in forced expiratory volume (FEV1) in normal subjects. Therefore, histamine-induced bronchoconstriction may be associated with the development of asthma symptoms. In addition, markedly increased levels of histamine have been observed during asthma reactions in humans.74,75 Histamine release can enhance the response induced by other bronchoconstricting mediators through both synergistic and non-synergistic mechanisms. However, the exact relationship between histamine and asthma remains incompletely understood. Histamine acts at multiple levels, often triggering similar outcomes due to its release of various other inflammatory mediators. This suggests that, for optimal bronchodilation therapy, histamine must be fully blocked, as its bronchoconstrictor effects cannot be managed or modulated.75,76 Serum histamine levels have been found to significantly increase in patients with mild and moderate asthma compared to healthy controls, with levels being notably higher in moderate asthma patients than in mild asthma patients. However, histamine levels are not particularly elevated in patients with severe asthma. Additionally, histamine increases airway hyperresponsiveness in asthma patients.77,78
Histamine concentration is also elevated in the airways of individuals with asthma. Intranasal administration of histamine in various studies has been shown to mimic the symptoms and characteristic changes of asthma, including bronchial obstruction, bronchoconstriction, and inflammatory responses. Antihistamines are useful in managing conditions like rhinitis and allergic conjunctivitis, and they can help improve asthma outcomes. Asthma pulmonologists, allergists, and clinicians should deepen their understanding of the mechanisms contributing to histamine-related complications and reactions, as well as the adjustments needed when using antihistamines.75,79
Complications of Atopic Dermatitis
The precise relationship between atopic dermatitis (AD) and histamine release remains unclear. Mast cells are found close to nerves in both lesional and perilesional atopic skin, with their degranulation products often adjacent to myelinated A and unmyelinated C nerve fibers. There is a higher prevalence of mast cells in the papillary dermis, and their number correlates with the severity of clinical parameters of AD, such as CD4+ T lymphocyte infiltration into the dermal papillary layer and the intensity of pruritus. Additionally, the number of mast cells is associated with sensory nerves in the context of acute AD exacerbations. The presence of mast cells and neurogenin 1-expressing nerve fibers may be linked to the onset of pruritus in AD. These findings suggest a clear relationship between increased mast cell numbers in lesional skin and pruritus in AD.80 In vitro studies indicate that AD tissue extracts induce more significant histamine release compared to normal skin tissue, suggesting that in AD, sensitized mast cells release more histamine. This mechanism likely involves a mast cell pathway disorder, including actin rearrangement, costimulatory signals passing through the FcεRI scaffolding protein, and increased T cell infiltration and inflammatory cytokines in the lesional skin, ultimately reducing the signal transduced by histamine.81
On the other hand, some studies in which AD patients were sensitized to histamines or challenged with the disease did not demonstrate a clear relationship between the number of mast cells and the condition. Other studies have suggested that the response of AD skin, rather than the mast cells themselves, is hemodynamically linked to a decrease in skin pH following bacterial colonization and exacerbation. This finding suggests that a reduction in skin pH may serve as a universal trigger. In general, neither AD nor histamine-induced pruritus (HIPr) is directly associated with mast cells. Instead, they are linked to a complex histamine-pruritus pathway involving various individual signaling pathways.82 While histamine is a potent and long-lasting mediator of pruritus in skin conditions related to itching, its effects are typically short-term and do not contribute to chronic itching.83 Additionally, the number of mast cells that release histamine during the early stages of repair significantly decreases within three days after mechanical injury. Limited data on the increase in cytokines produced by mast cells has been found in a small number of patient studies, especially those involving anaphylactic skin reactions induced by acupuncture or skin irritation. Histamine-induced itching mediated by the chemokine ligand 27 (CCL27) has been observed, but these findings are not yet widely generalizable.84
Gastrointestinal Complications
Histamine released from gut-derived mast cells, as well as exogenously administered histamine, can induce several luminal effects and complications within the gastrointestinal (GI) tract.85,86 This section explores the release of histamine in the intestinal lumen and its related gastrointestinal complications.
Gastric acid secretion
Mast cell degranulation results in the release of preformed histamine, which rapidly stimulates parietal cells to secrete more gastric acid through H2R-mediated pathways.87 Recent advancements in the understanding of human gastrointestinal afferent nerves have highlighted the role of vagal sensory input in regulating H2R, indicating a central role for histaminergic pathways in both afferent and efferent signaling in the stomach. Additionally, gastrin, through its receptors for cholecystokinin-2 (CCK-2), is present in mast cells in the oxyntic mucosa, contributing to the prostaglandin E2-mediated hyperreactive release of histamine from the oxyntic mucosa in conditions like peptic ulcer disease.88 Histamine also promotes the entry of vagal afferents acting on H3R, which reduces peristalsis through vagal efferents, leading to stasis and a reduction in pH. This process may contribute to luminal damage due to prolonged exposure of the mucosa to acid.89
Abnormal gastrointestinal motility
Intestinal inflammation, bacterial toxins, and gastrointestinal afferents activate gut mast cells, increasing their histamine production. Crosstalk between immune tolerance mechanisms, enteric nerves, and mucosal immune activation enables these effects, particularly under conditions of low mast cell threshold responses in a stressed intestine.52,90 Histamine further activates afferent nerves, which can reduce motility and cause tissue injury if the response is prolonged.91 This progression—from understanding the basic histamine pathways to describing how initial stress leads to immune activation and, eventually, both histaminergic and non-histaminergic effects on the intestine—is crucial for understanding the mechanisms underlying abdominal pain and motility changes driven by gut mast cells.92 This highlights the ongoing relevance of histamine-related research in gastrointestinal studies, offering valuable insights for practicing gastroenterologists.
Cardiovascular Complications
Hemodynamic and immuno-inflammatory aspects
Histamine plays a critical role in cardiovascular function by influencing hemodynamic parameters and immune-inflammatory responses. It induces vasodilation and increases vascular permeability, leading to decreased blood pressure and potential hypotensive episodes. These effects are primarily mediated through H1R and H2R in vascular endothelial cells. In addition, histamine activates immune cells, such as mast cells and basophils, triggering the release of pro-inflammatory mediators that can exacerbate inflammatory responses within the cardiovascular system. This dual role in regulating both hemodynamics and immune responses contributes to the complex cardiovascular manifestations associated with histamine.30,93
Vascular dysfunction and possible pathological effects on the cardiovascular system
Histamine-induced vascular dysfunction can lead to various pathological effects on the cardiovascular system. The resulting vasodilation and increased vascular permeability can cause edema and contribute to the development of atherosclerosis by promoting endothelial cell dysfunction and inflammatory cell infiltration. Additionally, histamine’s effects on smooth muscle cells can trigger vasospasms, potentially leading to conditions such as angina or, in severe cases, myocardial infarction. Furthermore, histamine-induced alterations in vascular tone and permeability can worsen hypertension and contribute to the progression of cardiovascular diseases.30,94,95
Histamine-mediated vascular permeability
Histamine is a key mediator that influences vascular permeability by acting on cell surface receptors. It increases permeability in postcapillary venules by enhancing endothelial contractility and promoting the formation of endothelium-cell gaps. These gaps, ranging from 200 to 1000 nm, contribute to increased vascular permeability.96 Various proteins localized in the endothelial cell junctions, such as actin, are integral to regulating this permeability. The involvement of ERK1/2 and p38 MAPK, both inside and outside these junctions, plays a critical role in the formation of postcapillary venous gaps and the disruption of the vascular barrier. Activation of ERK1/2 has been linked to an increase in the postcapillary venous barrier but a decrease in gap formation under certain conditions. ERK1/2 stimulation in the endothelium correlates with prolonged expression of E-selectin and ICAM-1, as well as the decoration of the endothelial cell by chemotactic proteins, CCL-16 and CCL-23.97 Additionally, histamine-releasing factor (HRF), along with leukosteroids and chemoattractant leukocytes, contributes to the release of corticostron. On the other hand, activation of p38 MAPK promotes gap formation, facilitating the mobilization of HMGB1 into the nucleus and triggering the expression of PBE, which is associated with a decrease in permeability. Furthermore, substance P activates p38 MAPK in HRP, helping to prevent barrier loss in the endothelium.98
Clinical implications
The clinical implications of histamine-induced cardiovascular complications are both broad and significant. For patients with preexisting cardiovascular conditions, histamine release can exacerbate symptoms and trigger acute events such as anaphylaxis or myocardial infarction. Histamine’s role in promoting inflammatory responses also contributes to the progression of chronic cardiovascular diseases, worsening patient outcomes over time. Clinicians must remain vigilant to these potential complications, especially in patients with known histamine sensitivities or those undergoing treatments that could elevate histamine levels. A thorough understanding of how histamine impacts the cardiovascular system is essential for developing effective management strategies and therapeutic interventions aimed at reducing these risks. One of the most notable cardiovascular complications induced by histamine is vasodilation, which can lead to hypotension and, in severe cases, shock.99,100
Respiratory complications
Histamine is widely recognized as both a causative agent and a risk factor for respiratory diseases. The underlying mechanisms of many respiratory complications can be attributed to exaggerated parasympathetic activity, bronchial smooth muscle contraction, exocrine gland secretion, and increased microvascular permeability.101 Histamine enhances microvascular permeability and promotes lymphocyte flow, contributing to inflammation and increasing mucus secretion from goblet cells in the airways, which triggers the pathophysiology of asthma. Bronchoconstriction is commonly evaluated by assessing phasic airway resistance using a forced oscillatory technique. Nonspecific hyperreactivity of the airways is observed in conditions such as asthma, chronic obstructive pulmonary disease, respiratory tract infections, and chronic cough.102,103 While clinicians are generally aware of histamine-induced bronchoconstriction, they may overlook the increased airway responsiveness that often accompanies it. Pulmonologists can add symptoms like chronic cough, rhinitis, chest tightness, and sputum production to their recommendations, advising patients to avoid histamine intake to help manage asthma. Additionally, allergists who treat asthma tend to focus on reducing eosinophilic inflammation but may not address chronic bronchitis. Respiratory therapists are encouraged to use exercise rehabilitation to manage environmental triggers, such as dust and cat allergens, which may affect athletes. However, they often place greater emphasis on managing nasal congestion from vasodilation rather than sneezing due to a reduction in airway resistance.104,105
Central Nervous System Complications
Histamine is present in the mammalian brain and has long been implicated in arousal processes in vitro. It can enhance the release of norepinephrine from brain tissue slices and has been shown to stimulate the release of acetylcholine in the brainstem in vitro. However, not all studies have consistently reported increases in central cholinergic neurotransmission.106,107 The administration of histidine to mice has been found to amplify the toxicity of cholinesterase inhibitors. Some have suggested that the ingestion of large amounts of tyramine can trigger histamine release in humans. Increased histamine levels or cholinergic transmission have significant implications for conditions like stroke or any situation involving exogenous or endogenous cholinesterase blockers.108,109 These histaminergic effects in the central nervous system align with the behavioral effects previously discussed. For example, intraventricular (icv) injection of histamine during the day has been shown to increase sleep, similar to the effects of melatonin or nicotinic acid. Histamine has also been demonstrated to suppress melatonin, and vice versa.110,111 As with its effects on the nose, exogenous histamine lowers body temperature and likely reduces metabolic rate. Both histamine and acetylcholine are considered endothermic neurotransmitters, meaning that increasing either can potentially lower metabolic rates. Mavents, a monoamine oxidase inhibitor, can elevate levels of catecholamines, histamine, or both. Provigil, a weak nonspecific stimulant, works by increasing histamine concentrations in the brain. Histamine is closely linked to serotonin syndromes and plays a significant role in the effects of nicotine and other drug-like substances in the brain. It is also a key factor in withdrawal from alcohol and nicotine, as well as in the obsessive-compulsive behaviors found in individuals with schizophrenia, depression, anxiety, and obsessive-compulsive disorder.112,113
Role of Histamine in Inflammation and Immune Responses
Histamine in inflammation
Histamine is a biogenic amine that plays a critical and multifaceted role in inflammation. Mast cells and basophils are typically the primary sources of histamine during inflammatory processes. In the classical view, mast cells store immune mediators, including histamine, tryptase, and a variety of cytokines, which can be rapidly released when the high-affinity IgE receptoris cross-linked.114 Beyond IgE-mediated allergic responses, mast cells can be activated by a range of other stimuli, such as antigen exposure, pathogens, certain drugs and venoms, cytokines, neuropeptides, complement anaphylatoxins, and physical stimuli like temperature and osmotic pressure. These non-IgE stimuli activate mast cells through pattern recognition receptors, including Toll-like receptors and specific G-protein-coupled receptors (GPCRs). This activation leads to the hydrolysis of phospholipids and the production of various pro-inflammatory mediators, including histamine.115,116 Upon activation, the plasma membrane phospholipase A2 is recruited to release arachidonic acid, which is subsequently metabolized by 5-lipoxygenase (5-LOX) to generate cytokines like IL-6 and leukotriene B4 (LTB4), promoting the recruitment of eosinophils and neutrophils. Additionally, phospholipase C generates diacylglycerol (DAG) and inositol trisphosphate (IP3), leading to Ca2+ influx and the exocytosis of preformed mediators, such as histamine, from granules.117 Histamine plays a central role in modulating various aspects of the inflammatory response. First, it is well established that the pre-existing mediator, histamine, is rapidly released from tissue mast cells to enhance the pro-inflammatory response during acute tissue injury. The pathophysiological roles of mast cell-derived histamine in angioedema and scratch-induced inflammation have been investigated. When histamine signaling was specifically blocked in mast cell-deficient mice, increased vascular permeability, increased neutrophil recruitment, and elevated IL-6 levels appeared in the acute phase of inflammation. During acute inflammation, mast cells release a variety of pre-formed and late-formed mediators, including TNF, IL-6, IL-13, prostaglandins, leukotrienes, and vasoactive amines.118,119 Pro-inflammatory mediators and cytokines induce chemokines to recruit inflammatory leukocytes into tissues. In contrast, late-phase anti-inflammatory mediators, such as IL-10, reprogram the mast cell response and inhibit chemokine production to restore homeostasis. Finally, the intriguing role of histamine in the pathogen-clearing response is explored. Prior to mast cell degranulation, histamine released from myeloid progenitors in tissues acts on neutrophil H1R to up-regulate CXCR2 and enhance neutrophil trafficking to the site of inflammation. Additionally, mast cell-associated granule-associated tryptase promotes the production of IL-6 by epithelial cells, which could further recruit eosinophils and neutrophils. This suggests a paradoxical role for mast cells during the early phase of inflammation.120,121 On the one hand, mast cells serve as the first responders and pathogen-sensing sentinels in tissues that promote inflammation and release multiple mediators to dampen inflammation.122
Histamine in immune responses
The mediatory role of histamine in immune cell regulation has steadily gained attention in recent decades. Although it has long been viewed as a principal pro-inflammatory molecule, and is indeed involved in that capacity, recent literature reveals additional paradoxical functions, including the down-regulation of T cell immunity. In this overarching role of both downregulating and perpetuating immune responses, histamine may also act as a feedback signal to exert a protective role against excessive inflammation.2,123 As a versatile molecule with multiple receptors, histamine is capable of orchestrating the function of a wide variety of immune cells, including adjusting their capacity to respond to antigens. This review explores the various roles of histamine in mediating immune cell function. The activation of H1R, H2R, and H3R histamines mediates the pro-inflammatory functions of histamine. These receptors regulate a variety of pathways within immune cells, leading to altered cytokine production, eosinophil migration, bronchial smooth muscle contraction, and increased endothelial cell permeability. However, beyond its regulatory capacity in eliciting pro-inflammatory responses, numerous studies have also characterized H4R-mediated functions of histamine, including mucus production and chemotaxis of immune cells, as well as potential immune-modulatory roles of histamine.124,125 This more obscure function of histamine is of great interest, as, under certain pathogenic conditions, it may be capable of curtailing excessive and potentially dangerous immune responses. The histamine immunological pathway is activated by allergens when they come into contact with the epithelial surface, resulting in an IgE-mediated hypersensitivity reaction. This sequence of events begins when allergens compromise the epithelial barrier and interact with antigen-presenting dendritic cells (APCs). These APCs process the allergen and subsequently present the allergen-derived peptide to naïve T cells, initiating their differentiation into T-helper 2 (TH2) cells. TH2 cells, through interactions such as CD28-CD80/86 and CD40-CD40L with B cells, facilitate the switching of B cell classes and the generation of allergen-specific IgE antibodies. Additionally, IgE antibodies adhere to Fc receptors located on the surface of mast cells, thereby sensitizing them for future interactions with the specific allergen. Upon reexposure to the allergen, the allergen-specific IgE present on mast cells undergoes crosslinking with the allergen, which triggers the degranulation of mast cells and the subsequent release of histamine as well as other inflammatory mediators. This process culminates in a range of allergic manifestations, which can include allergic conjunctivitis, allergic rhinitis, dyspnea, coughing, abdominal discomfort, and atopic dermatitis (Figure 4).126–128 Histamine’s regulatory roles in the immune system were first suggested more than 60 years ago when it was reported that some bacterial infections, and particularly anaphylactic shock, were associated with a sudden drop in circulating antibodies. This was paralleled by the observation that passive serum transfer from histamine-free rats was insufficient to confer protection. In other words, these early studies suggested a central role for histamine in the regulation of humoral immune responses.129,130
The effects of histamine on the humoral immune responses of rabbits, guinea pigs, and mice were also studied, and it was observed that histamine inhibited the production of antibodies. However, there were no significant differences in antibody titers when preimmunization serum was transferred from rabbits treated with histamine.131,132 More recent studies have similarly demonstrated the role of histamine in down-regulating antibody production in mice, based in part on the activation of histamine H2R. This effect involves inhibiting the IgM secretory responses of naive B cells in treatment. Furthermore, it suppresses the switch of the IgE class in memory B cells through H2R-dependent modulation of the cellular response to IL-4. Alongside its ability to down-regulate humoral immunity, new evidence has emerged of a similar function of histamine in Th immune responses. Specifically, the accumulation of naive CD4+ T cells migrating to CCL3 into Th2-promoting tissues during helminth infection skews the response of T cells toward the Th2 pathway. Since inflammasome activation in macrophages by helminth excretory secretory products greatly enhances CCL3 expression, it is believed that local histamine production in tissue macrophages provides a necessary feedback mechanism that dampens T cell activation.107,133,134
Figure 4 Allergen-induced immune response and histamine release.
Regulation of immune cell function
Histamine, a biogenic amine, exerts its effects through four G protein-coupled receptors: H1R, H2R, H3R, and H4R. H3R is primarily involved in the regulation of histamine release, while H4R is predominantly expressed by cells of the immune system. H4R activation has been shown to mediate the effects of histamine on various types of immune cells, including eosinophils, basophils, mast cells, helper 2 T lymphocytes, dendritic cells, macrophages, and natural killer cells.135,136 Histamine plays a significant role in pro-inflammatory Th2-type immune responses while exerting immunosuppressive effects in Type 1 immune responses. Mast cells and basophils, which are the effector cells of allergy, are involved in Th2-type immune responses. Upon exposure to allergens, these cells are activated by cross-linking of IgE-bound high-affinity IgE receptors, leading to the synthesis and release of various mediators, including histamine. Mast cell- and basophil-derived histamine is believed to enhance Th2-type immune responses via H1R, as well as increase the activity of these cells through H4R. H2R may attenuate the effects of histamine on amplifying Th2-type immune responses.137–139 In contrast, in Type 1 immune responses, histamine acts directly on T cells to inhibit their proliferation and Th1 cytokine production via H2R. The pivotal roles of mast cell- and basophil-derived histamine in promoting Th2-type immune responses have been demonstrated in the models of contact sensitivity and allergic inflammation of the airways. Dendritic cells play an important role in the induction of immune responses. Although normal dendritic cells are poorly responsive to histamine, they become responsive to histamine after maturation. Histamine enhances Langerhans cell migration from the epidermis to the draining lymph nodes via H1R and H4R, promoting Th2 differentiation of naive T cells in the draining lymph nodes. Histamine-activated dendritic cells stimulate Th2-differentiated T cells to produce IL-4, IL-5, and IL-13, thereby enhancing mast cell proliferation through H4R.134,140 Moreover, there is cross-talk between histamine and other mediators, such as prostaglandin D2 and cytokines, produced during Th2-type immune responses. Prostaglandin D2 amplifies Th2-type immune responses by inducing Th2 cell proliferation via CRT H2R and by promoting dendritic cell maturation and migration to lymph nodes. A good example of the involvement of a cascade of mediators is mast cell-derived prostaglandin D2, which upregulates antigen presentation and IL-10 and IL-12 expression by dendritic cells. This upregulation of IL-10 has been shown to promote Th2 differentiation, while further investigation revealed that upregulation of IL-12 production results in Th1 differentiation.141,142
Pharmacological interventions targeting histamine
Treatment of HIT largely depends on the severity of symptoms. In patients with mild to moderate symptoms, the offending agent should be discontinued, and patients treated with antihistamines. Improvement is typically observed within one to two hours. In patients with more severe and life-threatening symptoms, immunosuppressive drugs are necessary. Systemic corticosteroids are often prescribed for these patients, as these drugs reduce inflammation caused by histamine. Monoclonal antibodies, which can specifically block histamine activities, are also available for use. Additionally, desensitization can be considered a treatment option for patients with IgE-mediated or type I hypersensitivity reactions to their therapy. Desensitization is typically performed under the guidance of an allergist. The patient is initially given very low doses of the drug to block histamine-releasing activities. Subsequent doses are administered every 20 to 30 minutes, aiming to increase the amount of drug administered while inducing mast cell desensitization. If an allergic reaction occurs, the protocol is halted, and symptoms are treated before restarting at the last tolerated dose.133,143,144
Antihistamines
Antihistamines are a class of drugs designed to counteract the effects of histamine by blocking its receptors, thereby preventing or mitigating histamine-induced symptoms. They are commonly classified based on the specific histamine receptor they target. H1R antihistamines are widely used to treat allergic reactions, including hay fever, urticaria, and anaphylaxis. These drugs work by inhibiting histamine’s action on H1R, which reduces symptoms such as itching, swelling, and redness.145–147 First-generation H1R antihistamines, such as diphenhydramine, can cross the blood-brain barrier, often causing sedation. In contrast, second-generation H1R antihistamines, such as loratadine and cetirizine, are less likely to cause drowsiness due to their reduced ability to penetrate the central nervous system. H2R antihistamines, such as ranitidine and famotidine, are primarily used to reduce gastric acid secretion in conditions like peptic ulcers and gastroesophageal reflux disease (GERD). These drugs block histamine’s action on H2R in the stomach lining, thereby decreasing acid production.145,148,149 The development of these antihistamines has been crucial in managing a wide range of histamine-related conditions, offering relief from both allergic and gastrointestinal symptoms.
Mast cell stabilizers
Mast cell stabilizers are another category of pharmacological agents that target histamine-related pathways by preventing the release of histamine and other mediators from mast cells. These drugs work by inhibiting mast cell degranulation, a key step in the immune response that leads to histamine release. Cromolyn sodium and nedocromil are examples of mast cell stabilizers used in clinical practice.150,151 They are commonly employed in the management of allergic conditions such as asthma, allergic rhinitis, and conjunctivitis. By stabilizing the mast cell membrane, these drugs help reduce the frequency and severity of allergic reactions, making them particularly useful for chronic conditions in which histamine release plays a significant role.152–154 Mast cell stabilizers are often used as preventive measures rather than acute treatments, as they are more effective at preventing histamine release than at counteracting its effects after release.114,155 Their use represents a crucial strategy for managing diseases characterized by excessive mast cell activation and histamine release, offering a targeted approach to controlling symptoms and improving patient quality of life.
Emerging Research and Future Directions
The ability of histamine to induce various reactions within the body and its involvement in multiple diseases leaves many gaps in the literature, serving as a launching point for future research. Although histamine is a well-established mediator of underlying reactions such as oropharyngeal itch in allergic rhinitis, numerous areas of study focus on characterizing its effects. The fundamental mechanisms by which we sense histamine are still being investigated, and the intricate pathways leading to its activation are also under study. Improved technology has allowed for a greater depth and breadth of data to be collected, advancing the field into the realm of complex cellular signaling, moving away from the strict pharmacokinetics that were historically the focus.145,156 The complex intricacies of study designs and the data available to tailor pharmaceutical therapies to disease and genetic profiles still need to be refined. Knowing when to stimulate and when to inhibit histamine pathways in the treatment of two major chronic dysautonomias (POTS and MCAS) broadens the possibilities for future drug targets. This parallels the field of mast cell research, where we are continuously learning that histamine modulation—rather than merely blocking its receptor—can influence disease processes throughout the body across multiple disciplines. We also have a growing understanding of POTS, which links it to vagal disorders and reduced sympathetic excitation. A separate area gaining traction is the study of histamine and pain in human participants. Although H1R antihistamines are not a standard of care for headaches or fibromyalgia, it is not uncommon for individuals to self-medicate with these agents. Ongoing investigations in this domain focus on determining whether incorporating H1R antihistamines into standard therapeutic protocols is warranted for certain individuals with Central Sensitization Disorders, such as migraines and fibromyalgia, and, if so, identifying which patient populations would benefit the most.40,52,157
Conclusions
Histamine is an essential biogenic amine that plays a multifaceted role in regulating the immune system and inflammatory processes. By interacting with four G protein-coupled receptors (H1R, H2R, H3R, and H4R), histamine influences a wide range of physiological and pathological reactions. Its involvement in Th2-type and Th1-type immune responses highlights its complexity in immune modulation, as it contributes to allergic conditions such as allergic rhinitis, asthma, and atopic dermatitis, while also acting as an immunosuppressant. Histamine’s effects on various immune cells, including mast cells, basophils, eosinophils, dendritic cells, and T cells, play a role in a broad spectrum of allergic, autoimmune, and inflammatory conditions. Pharmacological interventions, such as antihistamines and mast cell stabilizers, are effective in managing symptoms caused by histamine. Both H1R and H2R antihistamines are important for treating allergic reactions and gastric acid-related conditions, while mast cell stabilizers are crucial for preventing histamine release in chronic allergic conditions. Ongoing research into the role of histamine in other medical conditions, such as POTS, MCAS, and pain disorders like migraines, continues to expand the therapeutic applications of histamine-targeted treatments. As current studies further explore the intricate signaling pathways and broader implications of histamine, it is clear that histamine modulation holds great promise for personalized therapies in various immune and inflammatory conditions. Understanding the complex role of histamine in immune regulation and its potential treatment goals is essential for developing more accurate and effective therapies for histamine-related diseases in the future.
Authors Contributions
AAl and MS wrote the text of main manuscript, and SD, DY, and MW collected the data, SS responsible for conception and design of the paper. All authors reviewed the manuscript.
Conflict of Interest
The authors declare no conflict of interest.
Funding
This work was supported by the National Natural Science Foundation of China (NSFC, 81773828), the Natural Science Foundation of Hebei Province (H2022206295), and the Science Fund for Creative Research Groups of the Natural Science Foundation of Hebei Province (No. H2020206474).
REFERENCES
1 O’Mahony L, Akdis M, Akdis CA. Regulation of the immune response and inflammation by histamine and histamine receptors. J Allergy Clin Immunol. 2011;128(6):1153–62. 10.1016/j.jaci.2011.06.051.
2 de la P. Sarasola M, Taquez Delgado MA, Nicoud MB, Medina VA. Histamine in cancer immunology and immunotherapy. Current status and new perspectives, Pharmacol Res Perspect. 2021;9:e00778. 10.1002/prp2.778
3 Colas L, Magnan A, Brouard S. Immunoglobulin E response in health and disease beyond allergic disorders, Allergy. 2022; 77:1700-18. 10.1111/all.15230
4 Kanagaratham C, El Ansari YS, Lewis OL, Oettgen HC. IgE and IgG antibodies as regulators of mast cell and basophil functions in food allergy, Front Immunol. 2020;11: 603050. 10.3389/fimmu.2020.603050
5 Qian H, Shu C, Xiao L, Wang G. Histamine and histamine receptors: Roles in major depressive disorder. Front Psychiatry. 2022;13: 825591. 10.3389/fpsyt.2022.825591
6 Seldeslachts A, Peigneur S, Tytgat J. Histamine receptors: ex vivo functional studies enabling the discovery of hits and pathways. Membranes (Basel). 2023;13: 897. 10.3390/membranes13120897
7 Ackermann F, Stanislas T. The plasma membrane-an integrating compartment for mechano-signaling. Plants. 2020;9: 505. 10.3390/plants9040505
8 Choromańska A, Chwiłkowska A, Kulbacka J, Baczyńska D, Rembiałkowska N, Szewczyk A, Michel O, Gajewska-Naryniecka A, Przystupski D, Saczko J. Modifications of plasma membrane organization in cancer cells for targeted therapy. Molecules. 2021;26:1850. 10.3390/molecules26071850
9 Stangherlin A, Zaccolo M. Phosphodiesterases and subcellular compartmentalized cAMP signaling in the cardiovascular system. Am J Physiol Circ Physiol. 2012; 302 :H379-90. 10.1152/ajpheart.00766.2011
10 Krishnaswamy G. Critical care management of the patient with anaphylaxis: a Concise definitive review. Crit Care Med. 2021;49:838-857. 10.1097/CCM.0000000000004893
11 Takazawa T, Yamaura K, Hara T, Yorozu T, Mitsuhata H, Morimatsu H. Working Group for the Preparation of Practical Guidelines for the Response to Anaphylaxis, Practical Guidelines for the Response to Perioperative Anaphylaxis, Safety Committee of the Japanese Society of Anesthesiologists. Practical Guidelines for the Response to Perioperative Anaphylaxis. J Anesth. 2021;1-16. 10.1007/s00540-021-03005-8
12 Hrubisko M, Danis R, Huorka M, Wawruch M. Histamine intolerance-the more we know the less we know. A review. Nutrients. 2021;13:2228. 10.3390/nu13072228
13 Zhao Y, Zhang X, Jin H, Chen L, Ji J, Zhang Z. Histamine intolerance-a kind of pseudoallergic reaction. Biomolecules. 2022;12:454. 10.3390/biom12030454
14 Akdis CA, Blaser K. Histamine in the immune regulation of allergic inflammation. J Allergy Clin Immunol. 2003;112:15-22. 10.1067/mai.2003.1585
15 Kubo M. Mast cells and basophils in allergic inflammation. Curr Opin Immunol. 2018;54: 74-79. 10.1016/j.coi.2018.06.006
16 Parsons ME, Ganellin CR. Histamine and its receptors. Br J Pharmacol. 2006;147:S127-S135. 10.1038/sj.bjp.0706440
17 Haas HL, Sergeeva OA, Selbach O. Histamine in the nervous system. Physiol Rev. 2008. 10.1016/B978-0-12-375073-0.50018-X
18 Levick SP. Histamine receptors in heart failure. Heart Fail Rev. 2022;27:1355-72. 10.1007/s10741-021-10166-x
19 Fiorani M, Del Vecchio LE, Dargenio P, Kaitsas F, Rozera T, Porcari S, Gasbarrini A, Cammarota G., Ianiro G. Histamine-producing bacteria and their role in gastrointestinal disorders. Expert Rev Gastroenterol. Hepatol. 2023;17:709-718. 10.1080/17474124.2023.2230865
20 Nuñez-Borque E, Fernandez-Bravo S, Yuste-Montalvo A, Esteban V. Pathophysiological, cellular, and molecular events of the vascular system in anaphylaxis. Front Immunol. 2022;13:836222. 10.3389/fimmu.2022.836222
21 Mottais A, Riberi L, Falco A, Soccal S, Gohy S, De Rose V. Epithelial-Mesenchymal Transition Mechanisms in Chronic Airway Diseases: A Common Process to Target? Int J Mol Sci. 2023;24. 10.3390/ijms241512412
22 Macarak EJ, Wermuth PJ, Rosenbloom J, Uitto J. Keloid disorder: Fibroblast differentiation and gene expression profile in fibrotic skin diseases. Exp Dermatol. 2021;30:132-145. 10.1111/exd.14243
23 Tripathi T, Shahid M, Khan HM, Siddiqui M, Khan AA, Khan RA. Histamine: Role in Pathogenesis of Autoimmune, Allergic, Inflammatory and Malignant Diseases. Biomed Asp Histamine Curr Perspect. 2011;201-226. 10.1007/978-90-481-9349-3_9
24 Sudarikova AV, Fomin MV, Yankelevich IA, Ilatovskaya DV. The implications of histamine metabolism and signaling in renal function. Physiol Rep. 2021;9:e14845. 10.14814/phy2.14845
25 Gorain B, Sengupta P, Dutta S, Pandey M, Choudhury H. Pharmacology of Histamine, Its Receptors and Antagonists in the Modulation of Physiological Functions. Front Pharmacol Neurotransmitters. 2020;213-240. 10.1007/978-981-15-3556-7_7
26 O’Mahony L, Akdis M, Akdis CA. Regulation of the immune response and inflammation by histamine and histamine receptors. J Allergy Clin Immunol. 2011;128:1153-62. 10.1016/j.jaci.2011.06.051
27 Jian T, Yang N, Yang Y, Zhu C, Yuan X, Yu G, Wang C, Wang Z, Shi H, Tang M. TRPV1 and PLC participate in histamine H4 receptor-induced itch. Neural Plast. 2016;2016:1682972. 10.1155/2016/1682972
28 Wang L, Gantz I, DelValle J. Histamine H2 receptor activates adenylate cyclase and PLC via separate GTP-dependent pathways. Am J Physiol Liver Physiol. 1996;271:G613-G620. 10.1152/ajpgi.1996.271.4.G613
29 Khawaja M, Thakker J, Kherallah R, Kitakaze M, Jneid H, Angiolillo DJ, Birnbaum Y. Antacid therapy in coronary artery disease and heart failure: proton pump inhibitors vs. H2 receptor blockers. Cardiovasc Drugs Ther. 2024;38:181-189. 10.1007/s10557-022-07358-4
30 Neumann J, Kirchhefer U, Dhein S, Hofmann B, Gergs U. The roles of cardiovascular H2-histamine receptors under normal and pathophysiological conditions. Front Pharmacol. 2021;12:732842. 10.3389/fphar.2021.732842
31 Chen Y-N, Sha H-H, Wang Y-W, Zhou Q, Bhuiyan P, Li N-N, Qian Y-N, Dong H-Q. Histamine 2/3 receptor agonists alleviate perioperative neurocognitive disorders by inhibiting microglia activation through the PI3K/AKT/FoxO1 pathway in aged rats. J Neuroinflamm. 2020;17:1-17. 10.1186/s12974-020-01886-2
32 Moriguchi T, Takai J. Histamine and histidine decarboxylase: Immunomodulatory functions and regulatory mechanisms. Genes Cells. 2020;25:443-449. 10.1111/gtc.12774
33 Nguyen PL, Cho J. Pathophysiological roles of histamine receptors in cancer progression: implications and perspectives as potential molecular targets. Biomolecules. 2021;11:1232. 10.3390/biom11081232
34 Oncel S, Basson MD. Gut homeostasis, injury, and healing: New therapeutic targets. World J Gastroenterol. 2022;28:1725. 10.3748/wjg.v28.i17.1725
35 Kim KH, Park J, Cho Y, Cho SY, Lee B, Jeong H, Lee Y, Yi J-W, Oh Y, Lee J-J. Histamine signaling is essential for tissue macrophage differentiation and suppression of bacterial overgrowth in the stomach. Cell Mol Gastroenterol Hepatol. 2023;15:213-36. 10.1016/j.jcmgh.2022.09.008
36 Seibel-Ehlert U, Plank N, Inoue A, Bernhardt G, Strasser A. Label-free investigations on the G protein dependent signaling pathways of histamine receptors. Int J Mol Sci. 2021;22:9739. 10.3390/ijms22189739
37 Lin C, Yan P, Lou Z, Shi X, Zhao Q, Li E. Effects of histamine on the neuroendocrine-immune regulatory network in the Pacific white shrimp, Litopenaeus vannamei. Aquaculture. 2022;554:738156. 10.1016/j.aquaculture.2022.738156
38 Neumann J, Binter MB, Fehse C, Marušáková M, Büxel ML, Kirchhefer U, Hofmann B, Gergs U. Amitriptyline functionally antagonizes cardiac H 2 histamine receptors in transgenic mice and human atria. Naunyn. Schmiedebergs. Arch Pharmacol. 2021;394:1251-1262. 10.1007/s00210-021-02065-7
39 Zhou Z, An Q, Zhang W, Li Y, Zhang Q, Yan H. Histamine and receptors in neuroinflammation: Their roles on neurodegenerative diseases. Behav Brain Res. 2024;465:114964. 10.1016/j.bbr.2024.114964
40 Dvornikova KA, Platonova ON, Bystrova EY. Inflammatory bowel disease: crosstalk between histamine, immunity, and disease. Int J Mol Sci. 2023;24:9937. 10.3390/ijms24129937
41 Conrad M, Söldner CA, Miao Y, Sticht H. Agonist binding and G protein coupling in histamine H2 receptor: A molecular dynamics study. Int J Mol Sci. 2020;21:6693. 10.3390/ijms21186693
42 Fukasawa T, Yoshizaki-Ogawa A, Enomoto A, Miyagawa K, Sato S, Yoshizaki A. Pharmacotherapy of itch-antihistamines and histamine receptors as G protein-coupled receptors. Int J Mol Sci. 2022;23:6579. 10.3390/ijms23126579
43 Jo MH, Kim BC, Sung K, Panettieri Jr RA, An SS, Liu J, Ha T. Molecular nanomechanical mapping of histamine-induced smooth muscle cell contraction and shortening. ACS Nano. 2021;15:11585-96. 10.1021/acsnano.1c01782
44 Si H, Wang J, Meininger CJ, Peng X, Zawieja DC, Zhang SL. Ca2+ release-activated Ca2+ channels are responsible for histamine-induced Ca2+ entry, permeability increase, and interleukin synthesis in lymphatic endothelial cells. Am J Physiol Circ Physiol. 2020;318:H1283-H1295. 10.1152/ajpheart.00544.2019
45 Gao S, Liu K, Ku W, Wang D, Wake H, Qiao H, Teshigawara K, Nishibori M. Histamine induced high mobility group box-1 release from vascular endothelial cells through H1 receptor. Front Immunol. 2022;13:930683. 10.3389/fimmu.2022.930683
46 Yang X, Shi Z, Wang X, Yang Y, Sun D, Zhu B, Song F, Zhu X, Ding S, Zou Y. Disruption of Histamine-H1R signaling exacerbates cardiac microthrombosis after periodontal disease via TLR4/NFκB-p65 pathway. Int Immunopharmacol. 2023;123:110774. 10.1016/j.intimp.2023.110774
47 Wang J, Liu B, Sun F, Xu Y, Luan H, Yang M, Wang C, Zhang T, Zhou Z, Yan H. Histamine H3R antagonist counteracts the impaired hippocampal neurogenesis in lipopolysaccharide-induced neuroinflammation. Int Immunopharmacol. 2022;110:109045. 10.1016/j.intimp.2022.109045
48 Honkisz-Orzechowska E, Popiołek-Barczyk K, Linart Z, Filipek-Gorzała J, Rudnicka A, Siwek A, Werner T, Stark H, Chwastek J, Starowicz K. Anti-inflammatory effects of new human histamine H3 receptor ligands with flavonoid structure on BV-2 neuroinflammation. Inflamm Res. 2023;72:181-194. 10.1007/s00011-022-01658-z
49 Baldo BA, Pham NH. Mechanisms of hypersensitivity. In: Drug Allergy: Clinical Aspects, Diagnosis, Mechanisms, Structure-Activity Relationships. Springer Nature; 2021. p. 59-137. 10.1007/978-3-030-51740-3_3
50 Yalew ST. Hypersensitivity reaction: Review. Int J Vet Sci Technol. 2020;4:28-32.
51 Mir MA, Mehraj U, Nisar S, Sheikh BA, Suhail S, Qayoom H. Hypersensitivity Reaction. In The Fundamentals of Hypersensitivities and Allergies. NOVA Bio-Medical Publishers New York USA. 2020. p. 1-34. 10.52305/KFXT3254
52 Smolinska S, Winiarska E, Globinska A, Jutel M. Histamine: a mediator of intestinal disorders-a review. Metabolites. 2022;12:895. 10.3390/metabo12100895
53 Dib K, Banna AEl, Radulescu C, Campos GLopez, Sheehan G, Kavanagh K. Histamine produced by Gram-negative bacteria impairs neutrophil’s antimicrobial response by engaging the histamine 2 receptor. J Innate Immun. 2023;15:153-73. 10.1159/000525536
54 Neumann J, Grobe JM, Weisgut J, Schwelberger HG, Fogel WA, Marušáková M, Wache H, Bähre H, Buchwalow IB, Dhein S. Histamine can be Formed and Degraded in the Human and Mouse Heart. Front Pharmacol. 2021;12:582916. 10.3389/fphar.2021.582916
55 Velasco E, Delicado-Miralles M, Hellings PW, Gallar J, Van Gerven L, Talavera K. Epithelial and sensory mechanisms of nasal hyperreactivity. Allergy. 2022;77:1450-63. 10.1111/all.15259
56 Tedner SG, Asarnoj A, Thulin H, Westman M, Konradsen JR, Nilsson C. Food allergy and hypersensitivity reactions in children and adults-A review. J Intern Med. 2022;291:283-302. 10.1111/joim.13422
57 Li H, Xiao Y, Li Q, Yao J, Yuan X, Zhang Y, Yin X, Saito Y, Fan H, Li P. The allergy mediator histamine confers resistance to immunotherapy in cancer patients via activation of the macrophage histamine receptor H1. Cancer Cell. 2022;40:36-52. 10.1016/j.ccell.2021.11.002
58 Provensi G, Costa A, Izquierdo I, Blandina P, Passani MB. Brain histamine modulates recognition memory: possible implications in major cognitive disorders. Br J Pharmacol. 2020;177:539-556. 10.1111/bph.14478
59 Hsu U-H, Chiang B-L. γδ T Cells and Allergic Diseases. Clin Rev Allergy Immunol. 2023;65:172-182. 10.1007/s12016-023-08966-0
60 Tsagareli MG, Nozadze I, Tsiklauri N, Carstens MI, Gurtskaia G, Carstens E. Thermal hyperalgesia and mechanical allodynia elicited by histamine and non-histaminergic itch mediators: respective involvement of TRPV1 and TRPA1. Neuroscience. 2020;449:35-45. 10.1016/j.neuroscience.2020.09.048
61 Aliotta GE. Characterization and modulation of histaminergic and non-histaminergic itch. PhD dissertation, Aalborg University; 2022.
62 Pankhurst TE, Montgomerie I, Marshall A, Draper SL, Bilbrough T, Button KR, Palmer OR, Hermans IF, Painter GF, Connor LM. A Glycolipid-Peptide-Hapten Tricomponent Conjugate Vaccine Generates Durable Antihapten Antibody Responses in Mice. ACS Chem Biol. 2024. 10.1021/acschembio.4c00214
63 Li C, Mao S, Montemayor Z, Traore MDM, Duran A, Ke F, Djibo M, Wen H, Gao W, Sun D. SARS-CoV-2 B Epitope-Guided Neoantigen NanoVaccines Enhance Tumor-Specific CD4/CD8 T Cell Immunity Through B Cell Antigen Presentation. BioRxiv. 2024:2007-24. 10.1101/2024.07.18.604163
64 Zheng L-F, Ji T, Guo Z-H, Wang T, Xiu X-L, Liu X-Y, Li S-C, Sun L, Xue H, Zhang Y. Na+-K+-2Cl-cotransporter 2 located in the human and murine gastric mucosa is involved in secretagogue-induced gastric acid secretion and is downregulated in lipopolysaccharide-treated mice. Eur J Pharmacol. 2020;880:173162. 10.1016/j.ejphar.2020.173162
65 Nazar W, Plata-Nazar K, Sznurkowska K, Szlagatys-Sidorkiewicz A. Histamine intolerance in children: a narrative review. Nutrients. 2021;13:1486. 10.3390/nu13051486
66 Shulpekova YO, Nechaev VM, Popova IR, Deeva TA, Kopylov AT, Malsagova KA, Kaysheva AL, Ivashkin VT. Food intolerance: the role of histamine. Nutrients. 2021;13:3207. 10.3390/nu13093207
67 Comas-Basté O, Sánchez-Pérez S, Veciana-Nogués MT, Latorre-Moratalla M, M del Vidal-Carou C. Histamine intolerance: The current state of the art. Biomolecules. 2020;10:1181. 10.3390/biom10081181
68 Nur Husna SM, Tan H-TT, Md Shukri N, Mohd Ashari NS, Wong KK. Allergic rhinitis: a clinical and pathophysiological overview. Front Med. 2022;9:874114. 10.3389/fmed.2022.874114
69 Drazdauskaitė G, Layhadi JA, Shamji MH. Mechanisms of allergen immunotherapy in allergic rhinitis. Curr Allergy Asthma Rep. 2021;21:1-17. 10.1007/s11882-020-00977-7
70 Klimov V, Cherevko N, Klimov A, Novikov P. Neuronal-immune cell units in allergic inflammation in the nose. Int J Mol Sci. 2022;23:6938. 10.3390/ijms23136938
71 Backaert W, Steelant B, Wils T, Qian Z, Dilissen E, Jonckheere AC, Boonen B, Jorissen M, Schrijvers R, Bullens MA. Nasal hyperreactivity in allergic rhinitis and chronic rhinosinusitis with polyps: a role for neuronal pathways. Rhinology. 2024;62:299-309. 10.4193/Rhin23.287
72 Jean EE, Good O, Rico JMI, Rossi HL, Herbert DR. Neuroimmune regulatory networks of the airway mucosa in allergic inflammatory disease. J Leukoc Biol. 2022;111:209-221. 10.1002/JLB.3RU0121-023R
73 Meng Y, Wang C, Zhang L. Diagnosis and treatment of non-allergic rhinitis: focus on immunologic mechanisms. Expert Rev Clin Immunol. 2021;17:51-62. 10.1080/1744666X.2020.1858804
74 van den Berg MPM, Nijboer-Brinksma S, Bos IST, van den Berge M, Lamb D, van Faassen M, Kema IP, Gosens R, Kistemaker LEM. The novel TRPA1 antagonist BI01305834 inhibits ovalbumin-induced bronchoconstriction in guinea pigs. Respir Res. 2021;22:1-15. 10.1186/s12931-021-01638-7
75 Meurs H, Zaagsma J. Pharmacological and biochemical changes in airway smooth muscle in relation to bronchial hyperresponsiveness. In: Inflamm. Cells Mediat. Bronchial Asthma, CRC Press; 2020. p. 1-38. 10.1201/9781003068679-1
76 Anderson SD, Kippelen P. A proposal to account for the stimulus, the mechanism, and the mediators released in exercise-induced bronchoconstriction. Front Allergy. 2023;4:1004170. 10.3389/falgy.2023.1004170
77 Cucca V, Ramirez GA, Pignatti P, Asperti C, Russo M, Della-Torre E, Breda D, Burastero SE, Dagna L, Yacoub M-R. Basal serum diamine oxidase levels as a biomarker of histamine intolerance: a retrospective cohort study. Nutrients. 2022;14:1513. 10.3390/nu14071513
78 Baioumy SA, Elgendy A, Ibrahim SM, Taha SI, Fouad SH. Association between serum zonulin level and severity of house dust mite allergic asthma. Allergy, Asthma Clin Immunol. 2021;17:86. 10.1186/s13223-021-00586-7
79 Vassilopoulou E, Konstantinou GN, Dimitriou A, Manios Y, Koumbi L, Papadopoulos NG. The impact of food histamine intake on asthma activity: a pilot study. Nutrients. 2020;12:3402. 10.3390/nu12113402
80 Ohsawa Y, Hirasawa N. The role of histamine H1 and H4 receptors in atopic dermatitis: from basic research to clinical study. Allergol Int. 2014;63:533-542. 10.2332/allergolint.13-RA-0675
81 De Vuyst E, Salmon M, Evrard C, de Rouvroit C Lambert, Poumay Y. Atopic dermatitis studies through in vitro models. Front Med. 2017;4:119. 10.3389/fmed.2017.00119
82 DeBenedetto A, Yoshida T, Fridy S, Park J-ES, Kuo I-H, Beck LA. Histamine and skin barrier: are histamine antagonists useful for the prevention or treatment of atopic dermatitis? J. Clin. Med. 2015;4:741-755. 10.3390/jcm4040741
83 Rossbach K, Nassenstein C, Gschwandtner M, Schnell D, Sander K, Seifert R, Stark H, Kietzmann M, Bäumer W, pruritus Histamine H1. H3 and H4 receptors are involved in pruritus. 2011;190:89-102. 10.1016/j.neuroscience.2011.06.002
84 Saeki H, Tamaki K. Role of TARC/CCL17 and CTACK/CCL27 in the Pathogenesis of Atopic Dermatitis. Curr Immunol Rev. 2008;4:221-229. 10.2174/157339508786447878
85 Chen M, Ruan G, Chen L, Ying S, Li G, Xu F, Xiao Z, Tian Y, Lv L, Ping Y. Neurotransmitter and intestinal interactions: focus on the microbiota-gut-brain axis in irritable bowel syndrome. Front Endocrinol (Lausanne). 2022;13:817100. 10.3389/fendo.2022.817100
86 Ma T, Xue X, Tian H, Zhou X, Wang J, Zhao Z, Wang M, Song J, Feng R, Li L. Effect of the gut microbiota and their metabolites on postoperative intestinal motility and its underlying mechanisms. J Transl Med. 2023;21:349. 10.1186/s12967-023-04215-2
87 Kovacheva E, Gevezova M, Maes M, Sarafian V. The mast cells-Cytokines axis in Autism Spectrum Disorder. Neuropharmacology. 2024;249:109890. 10.1016/j.neuropharm.2024.109890
88 Li Y, Feng Y, Liu L, Li X, Li X, Sun X, Li K, Zha R, Wang H, Zhang M. The baroreflex afferent pathway plays a critical role in H2S-mediated autonomic control of blood pressure regulation under physiological and hypertensive conditions. Acta Pharmacol Sin. 2021;42:898-908. 10.1038/s41401-020-00549-5
89 Tighilet B, Trico J, Marouane E, Zwergal A, Chabbert C. Histaminergic System and Vestibular Function in Normal and Pathological Conditions. Curr Neuropharmacol. 2024;22:1826-45. 10.2174/1570159X22666240319123151
90 Hasler WL, Grabauskas G, Singh P, Owyang C. Mast cell mediation of visceral sensation and permeability in irritable bowel syndrome. Neurogastroenterol. Motil. 2022;34:e14339. 10.1111/nmo.14339
91 Carthy E, Ellender T. Histamine, neuroinflammation and neurodevelopment: a review. Front Neurosci. 2021;15:680214. 10.3389/fnins.2021.680214
92 Guo Y, Wang B, Gao H, He C, Xin S, Hua R, Liu X, Zhang S, Xu J. Insights into the Characteristics and Functions of Mast Cells in the Gut. Gastroenterol Insights. 2023;14:637-652. 10.3390/gastroent14040043
93 Saheera S, Potnuri AG, Guha A, Palaniyandi SS, Thandavarayan RA. Histamine 2 receptors in cardiovascular biology: A friend for the heart. Drug Discov Today. 2022;27:234-45. 10.1016/j.drudis.2021.08.008
94 Guimbal S, Cornuault L, Rouault P, Hollier P-L, Chapouly C, Bats M-L, Imbault J, Gadeau A-P, Couffinhal T, Renault M-A. Mast cells are the trigger of small vessel disease and diastolic dysfunction in diabetic obese mice. Arterioscler Thromb Vasc Biol. 2021;41:e193-e207. 10.1161/ATVBAHA.121.315900
95 Noguchi K, Ishida J, Kim J-D, Muromachi N, Kako K, Mizukami H, Lu W, Ishimaru T, Kawasaki S, Kaneko S. Histamine receptor agonist alleviates severe cardiorenal damages by eliciting anti-inflammatory programming. Proc Natl Acad Sci. 2020;117:3150-6. 10.1073/pnas.1909124117
96 Majno G, Palade GE. Studies on inflammation: I. The effect of histamine and serotonin on vascular permeability: An electron microscopic study. J Cell Biol. 1961;11:571-605. 10.1083/jcb.11.3.571
97 Ferreira LKDP, Ferreira LAMP, Barros GCB, Monteiro TM, de Araújo Silva LA, de A Pereira R, Figueiredo PTR, Alves AF, Rodrigues LC, Piuvezam MR. MHTP, a synthetic alkaloid, attenuates combined allergic rhinitis and asthma syndrome through downregulation of the p38/ERK1/2 MAPK signaling pathway in mice. Int Immunopharmacol. 2021;96:107590. 10.1016/j.intimp.2021.107590
98 Zhao J-W, Ping J-D, Wang Y-F, Liu X-N, Li N, Hu Z-L, Ming L. Vitamin D suppress the production of vascular endothelial growth factor in mast cell by inhibiting PI3K/Akt/p38 MAPK/HIF-1α pathway in chronic spontaneous urticaria. Clin Immunol. 2020;215:108444. 10.1016/j.clim.2020.108444
99 Nicoud MB, Formoso K, Medina VA. Pathophysiological role of histamine H4 receptor in cancer: therapeutic implications. Front Pharmacol. 2019;10:556. 10.3389/fphar.2019.00556
100 Cataldi M, Borriello F, Granata F, Annunziato L, Marone G. Histamine receptors and antihistamines: from discovery to clinical applications. Hist Allergy. 2014;100:214-226. 10.1159/000358740
101 Baldo BA. Toxicities of opioid analgesics: respiratory depression, histamine release, hemodynamic changes, hypersensitivity, serotonin toxicity. Arc. Toxicol. 2021;95:2627-2642. 10.1007/s00204-021-03068-2
102 Ichinose M, Belvisi MG, Barnes PJ. Histamine H3-receptors inhibit neurogenic microvascular leakage in airways. J Appl Physiol. 1990;68:21-25. 10.1152/jappl.1990.68.1.21
103 White MV, Slater JE, Kaliner MA. Histamine and asthma. Am Rev Respir Dis. 1987;135:1165-76.
104 Manning PJ, O’Byrne PM. Histamine bronchoconstriction reduces airway responsiveness in asthmatic subjects. Am Rev Respir Dis. 1988;137:1323-5. 10.1164/ajrccm/137.6.1323
105 Tanaka S, Hirata K, Kurihara N, Yoshikawa J, Takeda T. Effect of loratadine, an H1 antihistamine, on induced cough in non-asthmatic patients with chronic cough. Thorax. 1996;51:810-4. 10.1136/thx.51.8.810
106 Mochizuki T. Histamine as an Alert Signal in the Brain. Funct Roles Histamine Recept. 2021;413-425. 10.1007/7854_2021_249
107 Obara I, Telezhkin V, Alrashdi I, Chazot PL. Histamine, histamine receptors, and neuropathic pain relief. Br J Pharmacol. 2020;177:580-599. 10.1111/bph.14696
108 Holeček M. Histidine in health and disease: metabolism, physiological importance, and use as a supplement. Nutrients. 2020;12:848. 10.3390/nu12030848
109 Virdi JK, Bhanot A, Jaggi AS, Agarwal N. Investigation on beneficial role of l-carnosine in neuroprotective mechanism of ischemic postconditioning in mice: possible role of histidine histamine pathway. Int J Neurosci. 2020;130:983-998. 10.1080/00207454.2020.1715393
110 Thakkar MM. Histamine in the regulation of wakefulness. Sleep Med Rev. 2011 Feb;15(1):65–74. 10.1016/j.smrv.2010.06.004.
111 Khan S, Khurana M, Vyas P, Vohora D. The role of melatonin and its analogues in epilepsy. Rev Neurosci. 2021;32:49-67. 10.1515/revneuro-2019-0088
112 Schellenberg RR, Ohtaka H, Paddon HB, Bramble SE, Rangno RE. Catecholamine responses to histamine infusion in man. J Allergy Clin Immunol. 1991;87:499-504. 10.1016/0091-6749(91)90008-C
113 Baumer W. Histamine, Serotonin, and their Antagonists. Vet Pharmacol Ther. 2017;267-81.
114 Theoharides TC, Perlman AI, Twahir A, Kempuraj D. Mast cell activation: beyond histamine and tryptase. Expert Rev Clin Immunol. 2023;19:639-654. 10.1080/1744666X.2023.2200936
115 Tontini C, Bulfone-Paus S. Novel approaches in the inhibition of IgE-induced mast cell reactivity in food allergy. Front Immunol. 2021;12:613461. 10.3389/fimmu.2021.613461
116 Katsoulis-Dimitriou K, Kotrba J, Voss M, Dudeck J, Dudeck A. Mast cell functions linking innate sensing to adaptive immunity. Cells. 2020;9:2538. 10.3390/cells9122538
117 Dutta BJ, Singh S, Seksaria S, Gupta GDas, Bodakhe SH, Singh A. Potential role of IP3/Ca2+ signaling and phosphodiesterases: Relevance to neurodegeneration in Alzheimer’s disease and possible therapeutic strategies. Biochem Pharmacol. 2022;201:115071. 10.1016/j.bcp.2022.115071
118 Komi DEA, Khomtchouk K, Maria PLSanta. A review of the contribution of mast cells in wound healing: involved molecular and cellular mechanisms. Clin Rev Allergy Immunol. 2020;58:298-312. 10.1007/s12016-019-08729-w
119 Dahlin JS, Maurer M, Metcalfe DD, Pejler G, Sagi-Eisenberg R, Nilsson G. The ingenious mast cell: Contemporary insights into mast cell behavior and function. Allergy. 2022;77:83-99. 10.1111/all.14881
120 Mihlan M, Wissmann S, Gavrilov A, Kaltenbach L, Britz M, Franke K, Hummel B, Imle A, Suzuki R, Stecher M. Neutrophil trapping and nexocytosis, mast cell-mediated processes for inflammatory signal relay. Cell. 2024;187:5316-35. 10.1016/j.cell.2024.07.014
121 AhYoung AP, Eckard SC, Gogineni A, Xi H, Lin SJ, Gerhardy S, Cox C, Phung QT, Hackney JA, Katakam AK. Neutrophil serine protease 4 is required for mast cell-dependent vascular leakage. Commun Biol. 2020;3:687. 10.1038/s42003-020-01407-0
122 Bansal SS. Immune Cells, Inflammation, and Cardiovascular Diseases. CRC Press. 2022. 10.1201/b22824
123 Kakavas S, Karayiannis D, Mastora Z. The complex interplay between immunonutrition, mast cells, and histamine signaling in COVID-19. Nutrients. 2021:13:3458. 10.3390/nu13103458
124 Branco ACCC, Yoshikawa FSY, Pietrobon AJ, Sato MN. Role of histamine in modulating the immune response and inflammation. Mediators Inflamm. 2018;2018:9524075. 10.1155/2018/9524075
125 Jutel M, Akdis M, Akdis CA. Histamine, histamine receptors and their role in immune pathology. Clin Exp Allergy. 2009;39:1786-800. 10.1111/j.1365-2222.2009.03374.x
126 Hong GU, Lim JY, Kim NG, Shin J-H, Ro JY. IgE and IgA produced by OX40-OX40L or CD40-CD40L interaction in B cells-mast cells re-activate FcεRI or FcαRI on mast cells in mouse allergic asthma. Eur J Pharmacol. 2015;754:199-210. 10.1016/j.ejphar.2015.02.023
127 Wang WW, Pan YL, Yu HW, Zhang B, Shao SW. Histamine H4 receptor regulates Th2-cytokine profile through thymic stromal lymphopoietin in allergic rhinitis. Eur Arch Otorhinolaryngol. 2019;276:1655-61. 10.1007/s00405-019-05369-w
128 Ahmad SF, Zoheir KMA, Ansari MA, Nadeem A, Bakheet SA, Al-Hoshani AR, Al-Shabanah OA, Al-Harbi MM, Attia SM. Histamine 4 receptor promotes expression of costimulatory B7. 1/B7. 2 molecules, CD28 signaling and cytokine production in stress-induced immune responses. J Neuroimmunol. 2015;289:30-42. 10.1016/j.jneuroim.2015.10.008
129 Benvenuto M, Focaccetti C, Ciuffa S, Fazi S, Bei A, Miele MT, Albonici L, Cifaldi L, Masuelli L, Bei R. Polyphenols affect the humoral response in cancer, infectious and allergic diseases and autoimmunity by modulating the activity of TH1 and TH2 cells. Curr Opin Pharmacol. 2021:60:315-330. 10.1016/j.coph.2021.08.005
130 Shawky H. Immunomodulatory Role of Histamine in Oncoimmunology and Immunotherapy: an update. In: Critical Developments in Cancer Immunotherapy; IGI Global. 2024; p. 351-390 10.4018/979-8-3693-3976-3.ch010
131 Jin C, Ye K, Luan H, Liu L, Zhang R, Yang S, Wang Y. Tussilagone inhibits allergic responses in OVA-induced allergic rhinitis guinea pigs and IgE-stimulated RBL-2H3 cells. Fitoterapia. 2020;144:104496. 10.1016/j.fitote.2020.104496
132 Palma GDe, Shimbori C, Reed DE, Yu Y, Rabbia V, Lu J, Jimenez-Vargas N, Sessenwein J, Lopez-Lopez C, Pigrau M. Histamine production by the gut microbiota induces visceral hyperalgesia through histamine 4 receptor signaling in mice. Sci Transl Med. 2022;14:eabj1895. 10.1126/scitranslmed.abj1895
133 Burghi V, Echeverría EB, Zappia CD, Nebreda AD, Ripoll S, Gómez N, Shayo C, Davio CA, Monczor F, Fernández NC. Biased agonism at histamine H1 receptor: Desensitization, internalization and MAPK activation triggered by antihistamines. Eur J Pharmacol. 2021;896:173913. 10.1016/j.ejphar.2021.173913
134 Alcain J, Cruz AdelPInfante, Barrientos G, Vanzulli S, Salamone G, Vermeulen M. Mechanisms of unconventional CD8 Tc2 lymphocyte induction in allergic contact dermatitis: Role of H3/H4 histamine receptors. Front Immunol. 2022;13:999852. 10.3389/fimmu.2022.999852
135 Mommert S, Jahn M, Schaper-Gerhardt K, Gutzmer R, Werfel T. Expression of histamine receptors H2R and H4R are predominantly regulated via the IL-4/IL-13 receptor type II on human M2 macrophages. Allergy. 2021;76:2886-90. 10.1111/all.14979
136 Schaper-Gerhardt K, Rossbach K, Nikolouli E, Werfel T, Gutzmer R, Mommert S. The role of the histamine H4 receptor in atopic dermatitis and psoriasis. Br J Pharmacol. 2020;177:490-502. 10.1111/bph.14550
137 Heo M-J, Choi SY, Lee C, Choi YM, An I, Bae S, An S, Jung JH. Perphenazine attenuates the pro-inflammatory responses in mouse models of Th2-type allergic dermatitis. Int J Mol Sci. 2020;21:3241. 10.3390/ijms21093241
138 Bao Y, Zhu X. Role of chemokines and inflammatory cells in respiratory allergy. J Asthma Allergy. 2022;1805-22. 10.2147/JAA.S395490
139 Shin S-H, Ye M-K, Lee D-W, Chae M-H, Han B-D. Nasal epithelial cells activated with alternaria and house dust mite induce not only Th2 but also Th1 immune responses. Int J Mol Sci. 2020;21:2693. 10.3390/ijms21082693
140 Steele MM, Lund AW. Afferent lymphatic transport and peripheral tissue immunity. J Immunol. 2021;206:264-72. 10.4049/jimmunol.2001060
141 Honda K, Arima M, Cheng G, Taki S, Hirata H, Eda F, Fukushima F, Yamaguchi B, Hatano M, Tokuhisa T. Prostaglandin D2 reinforces Th2 type inflammatory responses of airways to low-dose antigen through bronchial expression of macrophage-derived chemokine. J Exp Med. 2003;198:533-43. 10.1084/jem.20022218
142 Raible DG, Schulman ES, DiMuzio J, Cardillo R, Post TJ. Mast cell mediators prostaglandin-D2 and histamine activate human eosinophils. J Immunol (Baltimore, Md. 1950). 1992;148:3536-42. 10.4049/jimmunol.148.11.3536
143 Paradis L, Lavoie A, Brunet C, Bedard PM, Hebert J. Effects of systemic corticosteroids on cutaneous histamine secretion and histamine-releasing factor in patients with chronic idiopathic urticaria. Clin Exp Allergy. 1996;26:815-20. 10.1046/j.1365-2222.1996.d01-384.x
144 Matysiak J. Histamine intolerance (HIT). J Med Sci. 2022;91:e727. 10.20883/medical.e727
145 Ince M, Ruether P. Histamine and antihistamines. Anaesth Intensive Care Med. 2021;22:749-755. 10.1016/j.mpaic.2021.07.025
146 Parisi GF, Licari A, Papale M, Manti S, Salpietro C, Marseglia GL, Leonardi S. Antihistamines: ABC for the pediatricians. Pediatr Allergy Immunol. 2020;31:34-6. 10.1111/pai.13152
147 Du K, Qing H, Zheng M, Wang X, Zhang L. Intranasal antihistamine is superior to oral H1 antihistamine as an add-on therapy to intranasal corticosteroid for treating allergic rhinitis. Ann Allergy Asthma Immunol. 2020;125:589-96. 10.1016/j.anai.2020.06.038
148 Yanai K, Yoshikawa T, Church MK. Efficacy and safety of non-brain penetrating h1-antihistamines for the treatment of allergic diseases. Funct Roles Histamine Recept. 2021:193-214. 10.1007/7854_2021_265
149 Kobayashi T, Tonai S, Ishihara Y, Koga R, Okabe S, Watanabe T. Abnormal functional and morphological regulation of the gastric mucosa in histamine H2 receptor-deficient mice. J Clin Invest. 2000 Jun;105(12):1741–9. 10.1172/JCI9441.
150 Kılınç E, Baranoğlu Y. Mast cell stabilizers as a supportive therapy can contribute to alleviate fatal inflammatory responses and severity of pulmonary complications in COVID-19 infection. Anatol Clin J Med Sci. 2020;25:111-8. 10.21673/anadoluklin.720116
151 Marchitto MC, Chien AL. Mast cell stabilizers in the treatment of rosacea: a review of existing and emerging therapies. Dermatol Ther (Heidelb). 2021;11:1541-9. 10.1007/s13555-021-00597-7
152 Blaiss MS, Bernstein JA, Kessler A, Pines JM, Camargo CA, Fulgham P, Haumschild R, Rupp K, Tyler T, Moellman J. The role of cetirizine in the changing landscape of iv antihistamines: a narrative review. Adv Ther. 2022:1-15. 10.1007/s12325-021-01999-x
153 Wang L, Wang C, Lou H, Zhang L. Antihistamine premedication improves safety and efficacy of allergen immunotherapy. Ann Allergy Asthma Immunol. 2021;127:363-71. 10.1016/j.anai.2021.05.023
154 Parisi GF, Leonardi S, Ciprandi G, Corsico A, Licari A, Del Giudice MM, Peroni D, Salpietro C, Marseglia GL. Antihistamines in children and adolescents: A practical update. Allergol Immunopathol (Madr). 2020;48:753-62. 10.1016/j.aller.2020.02.005
155 Baran J, Sobiepanek A, Mazurkiewicz-Pisarek A, Rogalska M, Gryciuk A, Kuryk L, Abraham SN, Staniszewska M. Mast cells as a target-A comprehensive review of recent therapeutic approaches. Cells. 2023;12:1187. 10.3390/cells12081187
156 Nam JH, Kim K. The role of TRP channels in allergic inflammation and its clinical relevance. Curr Med Chem. 10.2174/0929867326666181126113015
157 Moya-García AA, Pino-Ángeles A, Sánchez-Jiménez F, Urdiales JL, Medina MÁ. Histamine, metabolic remodelling and angiogenesis: a systems level approach. Biomolecules. 2021;11:415. 10.3390/biom11030415