
Download
ORIGINAL ARTICLE
Acanthoside B attenuates NLRP3-mediated pyroptosis and ulcerative colitis through inhibition of tAGE/RAGE pathway
Xiaobo Hea, Chunfang Zhoua, Rui Shanga, Xiaoyan Wangb*
aDepartment of Gastroenterology, Renmin Hospital, Hubei University of Medicine, Shiyan, Hubei Province, China
bDepartment of Neurofunction, Renmin Hospital, Hubei University of Medicine, Shiyan, Hubei Province, China
Abstract
Acanthoside B (Aca.B), a principal bioactive compound extracted from Pogostemon cablin, exhibits superior anti-inflammatory capacity. Ulcerative colitis is a nonspecific inflammatory bowel disease with unknown etiology. The potential of Aca.B as a therapeutic agent for ulcerative colitis is also unknown and remains an area for future investigation. In this study, we established both in vitro and in vivo models to investigate ulcerative colitis, utilizing Llipopolysaccharide (LPS)-stimulated MODE-K cells and dextran sulfate sodium (DSS)-induced colitis in mice, respectively. The progression of ulcerative colitis was evaluated through histologic analysis, body weight monitoring, and assessment of disease activity index assessment. Furthermore, the effects on pyroptosis were detected through immunoblot analysis. We found that Aca.B treatment significantly ameliorated LPS-induced injury in MODE-K cells, as evidenced by increased cell viability and inhibition of inflammatory response. Moreover, the Aca.B treatment attenuated pyroptosis-specific protein expression, caspase-1 activation, and inflammatory cytokine secretion. In the animal study, Aca.B administration improved bowel symptoms in DSS-induced colitis mice model. This was accompanied by reductionsreduced inweight, colon shortening, inflammatory cell infiltration, and cell pyroptosis in vivo. Furthermore, Aca.B diminished the accumulation of advanced glycation end-products (AGE), resulting in a decrease in the expression of the receptor of AGE (RAGE) and downstream phosphorylated P65 expression. e.The inhibition of the inflammatory response and pyroptosis by Aca.B depends on suppressing the AGE/RAGE pathway. This study confirms the effects of Aca.B on pyroptosis and ulcerative colitis, providing a fundamental evidence for translating Aca.B into clinical applications as an anti-inflammatory medicine.
Key words: Acanthoside B, NOD-like receptor protein 3, pyroptosis, receptor for advanced glycation end-products, ulcerative colitis
*Corresponding author: Xiaoyan Wang, Department of Neurofunction, Renmin Hospital, Hubei University of Medicine, No. 39 Chaoyang Middle Road, Shiyan, Hubei Province 442000, China. Email address: [email protected]
Received 26 September 2024; Accepted 28 November 2024; Available online 1 January 2025
Copyright: He X, et al.
This open access article is licensed under Creative Commons Attribution 4.0 International (CC BY 4.0). http://creativecommons.org/licenses/by/4.0/
Introduction
Ulcerative colitis is a chronic, nonspecific inflammatory disease that primarily affects the colon and rectum. It is characterized by persistent and recurring mucosal inflammation.1 Common symptoms include abdominal pain and bloody mucopurulent diarrhea. The diagnosis of ulcerative colitis depends on colonoscopy and biopsy.2,3 The global prevalence of ulcerative colitis has been steadily increasing steadily in recent years, yet its underlying cause remains unknown. Previous studies have suggested that the activation of the NOD-like receptor protein 3 (NLRP3) inflammasome and pyroptosis play a significant role in the progression of ulcerative colitis. 4 However, the potential of drugs targeting pyroptosis to alleviate the severity of ulcerative colitis is still a topic of debate.
Emerging evidence reports the therapeutic potential of traditional Chinese medicine (TCM) for treating ulcerative colitis. For example, sophora flavescens (Kushen)–based TCM has shown superior efficacy in treating ulcerative colitis, as confirmed by clinical evidence.5 Furthermore, compound sophorae decoction has been provenis known to alleviate the symptoms of ulcerative colitis through rebalancing T-helper 17–T-regulatory (Th17/Treg) ratio in dextran sulfate sodium (DSS)-induced colitis mice model.6 Pogostemon cablin has been extensively applied in the treatment of gastroenteric diseases, particularly ulcerative colitis. Its extract has shown the ability to inhibit DSS-induced inflammatory response and cell apoptosis.7
Acanthoside B (Aca.B) is the primary bioactive compound isolated from Pogostemon cablin. Previous studies demonstrated that Aca.B could alleviate cognitive impairment in mice by inhibiting oxidative stress and activating the brain-derived neurotrophic factor (BDNF) signaling pathway.8 Additionally, Aca.B has been found to relieve lipopolysaccharide (LPS)-induced inflammatory cytokine secretion phenotype and pneumonia symptoms.9 However, further research is needed to explore the protective effect of Aca.B on DSS-induced colitis in mice.
Advanced glycation end-products (AGEs) are a group of heterogenous proteins associated with the pathology of various inflammatory diseases. The specific binding of AGEs with its receptor (RAGE) on the cell membranes leads to the overproduction of inflammatory factors and chemokines, leading to persistent inflammatory response.10 Notably, the AGE/RAGE pathway is documented to be a primary activator of NLRP3 inflammasome and pyroptosis.11 The aAccumulation of AGEs promotes the expression of NLRP3 and apoptosis-associated speck-like protein containing a caspase recruitment domain (CARD), the activation of caspase-1 and gasdermin D (GSDMD), and the production of various inflammatory cytokines, such as interleukin-1β (IL-1β), IL-10, IL-18, and tumor necrosis factor-α (TNF-α). Therefore, further study is essentialrequired to explore its pharmaceutical influence on the AGE/RAGE and pyroptosis pathways.
In the present study, we highlighted Aca.B as a promising therapeutic agent against ulcerative colitis. Our results demonstrated that, Aca.B ameliorated the severity of DSS-induced colitis and epithelial cell pyroptosis in mice models through attenuating the AGE/RAGE pathway activation. This study provided fundamental evidence supporting the translation of Aca.B into clinical application as an anti-inflammatory therapy.
Materials and Methods
Cell culture
MODE-K cell line (intestinal epithelial cell from mice) was purchased from Youze Biotech Co. (Changsha, China). MODE-K cells were cultured in RPMI-1640 medium (Gibco, Miami, FL, USA) with 10% fetal bovine serum (Gibco) and 1% penicillin–streptomycin cocktail (New Cell & Molecular Biotech, Shanghai, China) at 37°C under 5% CO2 atmosphere. To establish inflammatory cell injury models, the cells were treated with 1-μg/mL LPS for 4 h as described previously.12 Aca.B of various concentrations and 200 μg/mL AGE (Topscience Biotech, Shanghai, China) were administered to treat the LPS-stimulated MODE-K cells.
Establishing animals and ulcerative colitis model
All experiments with animals followed the guidelines approved by the Institutional Animal Care and Use Committee of Renmin Hospital, Hubei University of Medicine (Approval No. SYSRMYY-100), ensuring compliance with ethical standards. Male C57BL/6J mice (4-week old; n = 6 per group) were obtained from the Shanghai SLAC Animal Co. (Shanghai, China). Mice were housed under specific pathogen-free (SPF) conditions.13 Mice were anesthetized with isoflurane (1.5–2.0% v/v).
DSS-induced ulcerative colitis mice models were constructed as described previously.14 The manimals were administered 5% DSS orally or regular drinking for 7 days to induce intestinal inflammation. Aca.B was dissolved in 0.9% saline with 1% tween-80. For animal experiments, 30 male mice were divided into the following five groups: (1) mice with regular drinking (Sham group); (2) mice with DSS drinking (DSS group); (3) mice with DSS + Aca.B (5 mg/kg) drinking group; (4) mice with DSS + Aca.B (10 mg/kg) drinking group; and (5) mice with DSS + Aca.B (20 mg/kg) drinking group. During the experiments, the body weight and disease activity index (DAI) of each animal were assessed daily. After sacrifice, the colon length was measured, and colon samples and serum were harvested for subsequent experiments. Hematoxylin and eosin (H&E) staining was performed to evaluate inflammatory response and colon injury.
Cell counting kit-8 (CCK-8) assay
A CCK-8 assay kit (New Cell & Molecular Biotech, Shanghai, China) was used to evaluate the effect of Aca.B on cell viability. MODE-K cells were plated into a 96-well plate 24 h prior to treatment. After complete adherence, cells were treated with different concentrations of Aca.B with/without LPS stimulation. Then, 10 μL of CCK-8 solution was added into each well, and the absorbance at 450 nm was read using a microplate reader (Tecan, USA) 2 h after the reaction.
Quantitative real time-polymerase chain reaction (qRT-PCR) assay
After indicated treatments, cells were harvested for RNA isolation. Total RNA was purified using a RNA isolation kit (Qiagen, China), and the qRT-PCR was performed using a complementary DNA (cDNA) synthesis kit (Takara, Tokyo, Japan). A SYBR PCR SuperMix Plus (Bimake, Shanghai, China) was used in qRT-PCR; all primer sequences are listed in Table 1. The experimental data was analyzed using the 2(-Delta Delta threshold cycle [CT]) (2−ΔΔCT) method.
Table 1 The primer sequences used in quantitative real time-polymerase chain reaction (qRT-PCR).
| Gene | Sequence | |
|---|---|---|
| IL-10 | Forward | 5′-GCTCTTACTGACTGGCATGAG-3′ |
| Reverse | 5′-CGCAGCTCTAGGAGCATGTG-3′ | |
| TNF-α | Forward | 5′-CCCTCACACTCAGATCATCTTCT-3′ |
| Reverse | 5′-GCTACGACGTGGGCTACAG-3′ | |
| IL-6 | Forward | 5′-TAGTCCTTCCTACCCCAATTTCC-3′ |
| Reverse | 5′-TTGGTCCTTAGCCACTCCTTC-3′ | |
| RAGE | Forward | 5′-CTTGCTCTATGGGGAGCTGTA-3′ |
| Reverse | 5′-GGAGGATTTGAGCCACGCT-3′ | |
Enzyme-linked immunosorbent serological assay (ELISA)
Commercial ELISA kits for TNF-α (EK282HS; Multi Science Biotech, Shanghai, China), IL-6 (EK206; Multi Science Biotech), IL-10 (EK210; Multi Science Biotech), IL-1β (EK201B; Multi Science Biotech), IL-18 (EK218S; Multi Science Biotech), and AGE (0-008073; Jianglai Biotech, China) were used in this study. Cell culture supernatants or mice serum were harvested for ELISA assay, and all the experimental procedures were performed by following the provided protocols.
Western blot (WB) assay
Western blot A assay was performed based on a previously published study.15 After indicated treatments, cells or tissue samples were harvested for total protein isolation using 1% sodium dodecyl sulfate (SDS) lysis buffer (Beyotime, Shanghai, China) with 1% protease and phosphatase inhibitor cocktails (Pierce, Rockford, USA). Then, total protein samples were separated using sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) gel, transferred to polyvinylidene fluoride (PVDF) membranes, blocked with 5% bovine serum albumin (BSA), and incubated with primary antibodies for 24 h at 4°C. The following antibodies were used in this study: β-actin (1:5000; ab178787; Abcam, USA); RAGE (1:2000; ab216329; Abcam); p-P65 (1:500; ab176302; Abcam); P65 (1:1000; ab76311; Abcam); NLRP3 (1:3000; ab26399; Abcam); ACS (1:2500; ab175449; Abcam); GSDMD-N (1:1000; ab215203; Abcam); Caspase-1 (1:2000; ab179515; Abcam); IL-1β (1:1000; ab283818; Abcam); and IL-18 (1:1000; ab207323; Abcam). Finally, protein signals were detected with an enhanced chemiluminescence (ECL; Bio-Rad Hercules, CA, USA). The ImageJ software was used to quantify band density.
Flow cytometry (FCM) assay
A fluorescent caspase-1 activity detection kit (Beyotime) was used to evaluate pyroptosis in MODE-K cells by labeling 5-carboxyfluorescein-Tyr-Val-Ala-Asp-fluoromethylketone (FAM-YVAD-FMK) to the activated caspase-1 by following the instructions. After the indicated treatment, cells were incubated with FAM-YVAD-FMK working solution for 30 min in the dark, followed by centrifugation and washing. Then, the cells were stained with propidium iodide (PI) working solution for 10 min in the dark on ice. A flow cytometer (BD Biosciences, NJ, USA) was used to detect fluorescent signals, and only the Caspase-1+/PI+ cell was defined as a pyroptotic cell.
Drug target prediction
To predict the potential target of Aca.B, we analyzed Aca.B chemical structure on Swiss target prediction website (http://www.swisstargetprediction.ch/). Moreover, the pathway enrichment and network analysis were performed on the Metascope website (https://metascape.org/gp/index.html#/main/step1).
Statistical analyses
All data were expressed as mean ± standard deviation (SD). Statistical significance was determined using Student’s t-test or one-way ANOVA, followed by Tukey’s post hoc test; P < 0.05 was considered statistically significant. The SPSS 22.0 software (SPSS Inc, Chicago, IL) was used to analyze all data. Each assay was repeated independently for at least three times.
Results
Aca.B ameliorates LPS-induced MODE-K cell injury and inflammatory factor secretion
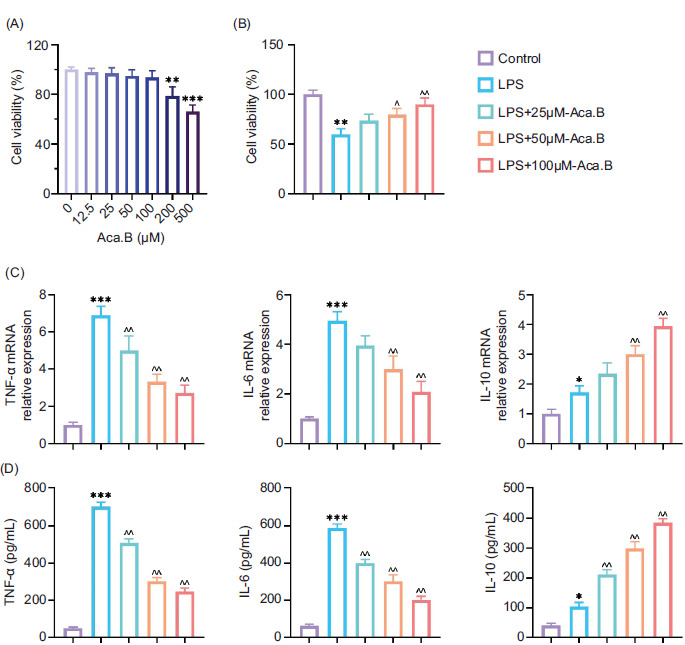
In order to explore the pharmaceutical effect of Aca.B on MODE-K cells, we first assessed its influence on cell viability at different concentrations. As demonstrated in Figure 1A, no toxicity was detected until the concentration reached 200 μM. Thus, in the study, the concentration of Aca.B was kept below 200 µM. LPS was utilized to mimic inflammatory microenvironment in vitro.16 Upon LPS stimulation, MODE-K cell viability decreased significantly decreased, and there was an increase was observed in the expression of inflammatory factors (TNF-α and IL-6) along with a reduction in anti-inflammatory IL-10 expression at both messenger RNA (the mRNA) and protein levels. eaHowever, Aca.B successfully ameliorated LPS-induced MODE-K cell injury and inflammatory factor secretion in a dose-dependent manner (Figures 1B–1D). This result indicated that Aca.B could alleviate inflammatory injury of intestinal epithelial cells in vitro.
Figure 1 Aca.B ameliorates LPS-induced MODE-K cell injury and inflammatory factor secretion. (A) Effect of Aca.B at various concentrations on MODE-K cell viability. (B) Aca.B at various concentrations promoted MODE-K cell survival under LPS treatment. (C) Aca.B at various concentrations affected inflammatory factor (TNF-α, IL-6 and IL-10) expression at the mRNA level, as detected by RT-PCR. (D) Aca.B at various concentrations affected.
Aca.B ameliorates NLRP3-mediated proptosis in MODE-K cell
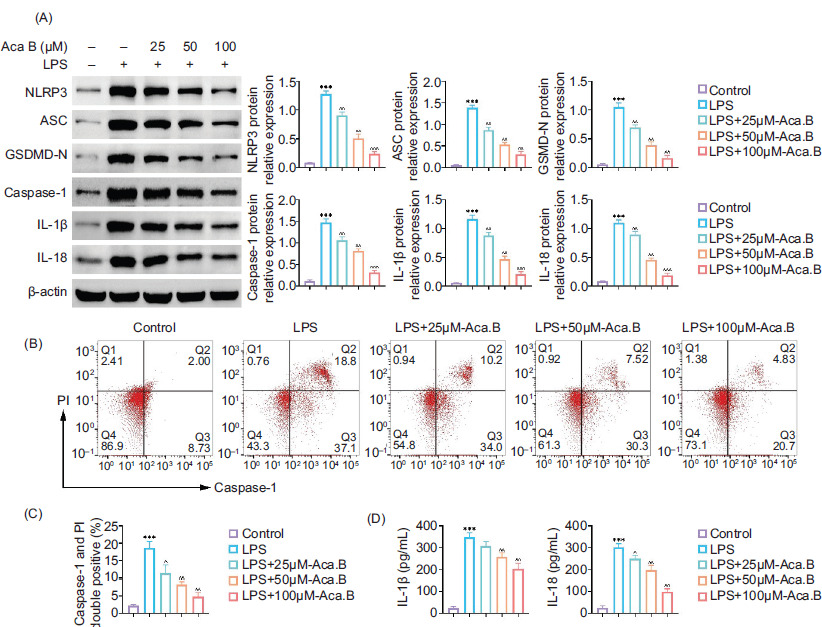
NLRP3-mediated pyroptosis is documented as a major inflammatory cell death.17 Therefore, we investigated whether Aca.B could attenuate MODE-K cell pyroptosis. As shown in Figure 2A, the upregulation of pyroptosis-specific biomarkers (NLRP3, ASC, GSDMD-N, Caspase-1, IL-1β, and IL-18) induced by LPS was dose-dependently reversed by Aca.B treatment. Caspase-1 activation is a central step of pyroptosis.18 Caspase-1 activity in MODE-K cells treated with LPS or Aca.B was evaluated using FCM. As expected, Aca.B significantly inhibited LPS-induced caspase-1 activation, decreasing the Caspase-1+–PI+ cell ratio from about 20% in the LPS group to 5% in the LPS+100 μM Aca.B group (Figures 2B and 2C). Notably, the LPS-induced expression of pyroptosis effectors in MODE-K cell supernatants was also reduced by Aca.B treatment (Figure 2D). These data suggested that Aca.B could alleviate NLRP3-mediated proptosis in MODE-K cells.
Figure 2 Aca.B ameliorates NLRP3-mediated proptosis in MODE-K cells. (A) Expressions of NLRP3, ASC, GSDMD-N, Caspase-1, IL-1β, and IL-18 were determined in MODE-K cells treated with LPS or Aca.B using immunoblotting. The grayscale value of the corresponding band was quantitated with ImageJ. (B) Caspase-1 activity in MODE-K cells treated with LPS or Aca.B was assessed using flow cytometry (FCM) analysis. (C) Quantitative statistics of double positive for caspase-1 and Propidium iodide (PI) staining. (D) Secretion of IL-1β and IL-18 was evaluated in MODE-K cells treated with LPS or Aca.B using ELISA. ***P < 0.001, compared with the control group. ^P < 0.05, ^^P < 0.01, and ^^^P < 0.001, compared with the LPS group. Aca.B: Acanthoside B.
Aca.B alleviates DSS-induced ulcerative colitis and NLRP3-mediated pyroptosis in mice
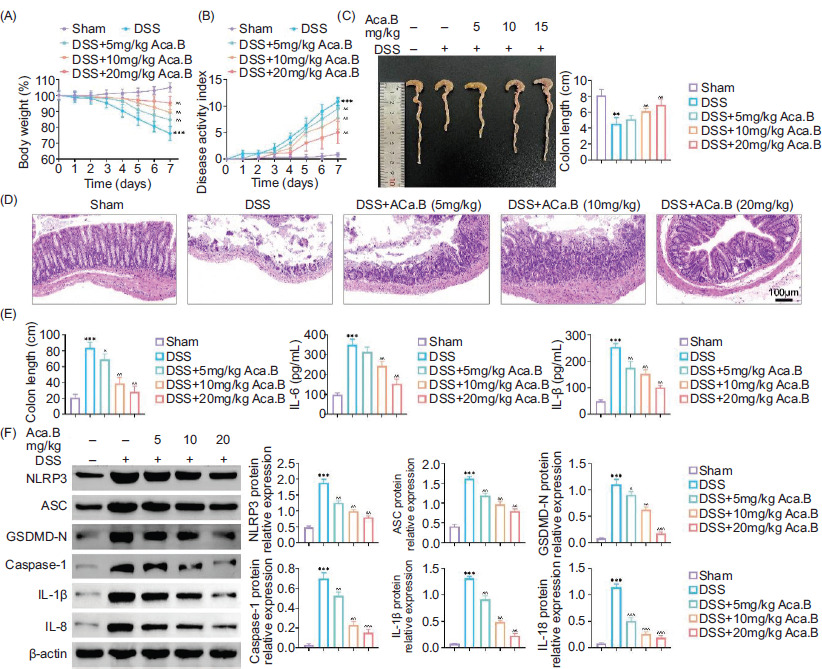
In order to confirm our findings in animal models, we established DSS-induced ulcerative colitis mice models, and Aca.B was administrated orally. Mice in the DSS group exhibited substantial weight loss, rectal bleeding, and diarrhea, resulting in significant decrease in body weight and an increase in DAI (Figures 3A and 3B). However, oral treatment with Aca.B effectively alleviated the severity of colitis in a dose-dependent manner. Additionally, Aca.B reversed the symptom of colon shortening (Figure 3C). Ulcerative colitis is characterized by intestinal epithelial injury, particularly damage of crypt structure, inflammatory cell infiltration, and loss of goblet cells.19 H&E staining revealed that Aca.B significantly ameliorated DSS-induced epithelial injury. Mice treated with Aca.B exhibited relative intact crypt architecture without ulceration or obvious inflammatory cell infiltration (Figure 3D). Notably, the levels of inflammatory cytokines (TNF-α, IL-6, and IL-1β) in the serum of mice treated with Aca.B were reduced (Figure 3E). Consistent with in vitro results, Aca.B also decreased the expression of pyroptosis-specific biomarkers (NLRP3, ASC, GSDMD-N, Caspase-1, IL-1β, and IL-18) in colon samples from DSS-induced ulcerative colitis mice in vivo. These results demonstrated that Aca.B alleviated DSS-induced ulcerative colitis and NLRP3-mediated proptosis in mice.
Figure 3 Aca.B alleviates dextran sulfate sodium (DSS)-induced ulcerative colitis and NLRP3-mediated proptosis in mice. The C57BL/6 mice were subjected to the DSS-induced ulcerative colitis. Aca.B in various concentrations was orally administrated twice a day for consecutive 7 days. (A) Change in body weight, (B) disease activity index (DAI), and (C) colon length were measured. (D) Pathological changes in colons were evaluated using H&E staining. (E) Inflammatory factor levels (TNF-α, IL-6, and IL-1β) in serum were assessed using ELISA. (F) Expressions of NLRP3, ASC, GSDMD-N, Caspase-1, IL-1β, and IL-18 were determined in tissue samples using immunoblotting analysis. The grayscale value of the corresponding band was quantitated with ImageJ. **P < 0.01 and ***P < 0.001, compared with the Sham group. ^P < 0.05, ^^P < 0.01, and ^^^P < 0.001, compared with the DSS group. Aca.B: Acanthoside B.
Aca.B inhibits the AGE/RAGE pathway in LPS-stimulated MODE-K cells and DSS-induced ulcerative colitis mice
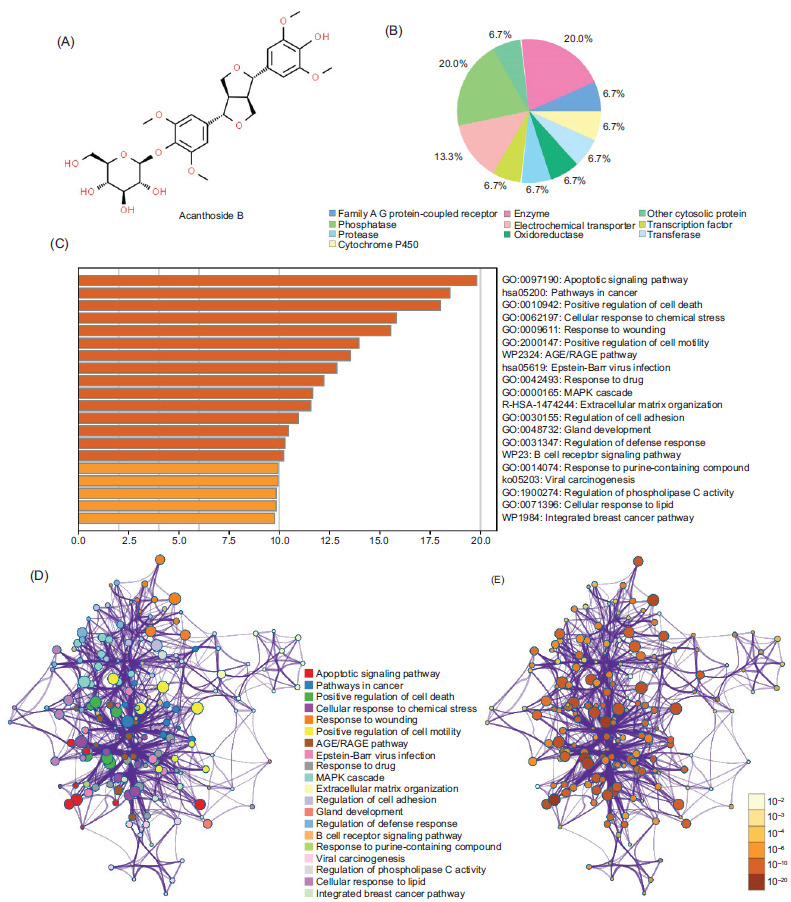
In order to understand the pharmaceutical effect of Aca.B, Swiss Target Prediction was employed to identify potential target pathways of Aca.B based on its chemical structure analysis (Figures 4A and 4B). By following the screening process, numerous binding targets predominantly associated with programmed cell death, response to stress, and cell homeostasis pathways were identified (Figure 3C). Moreover, network analysis illustrated the interconnections among the predicted pathways (Figures 3D and 3E). Notably, our attention was drawn to the AGE/RAGE pathway, known to be a key regulator of inflammatory response.20 Therefore, we postulated that the inhibition of inflammatory response by Aca.B might depend on the AGE/RAGE signaling pathway.
Figure 4 The Aca.B target pathways prediction. (A) The chemical structure of Aca.B. (B) The predicted binding targets of Aca.B using Swiss Target Prediction website. (C) The predicted downstream signaling pathways of Aca.B using Swiss Target Prediction website. (D and E) Network analysis of the predicted downstream signaling pathways. The node color reflects P value. Aca.B: Acanthoside B.
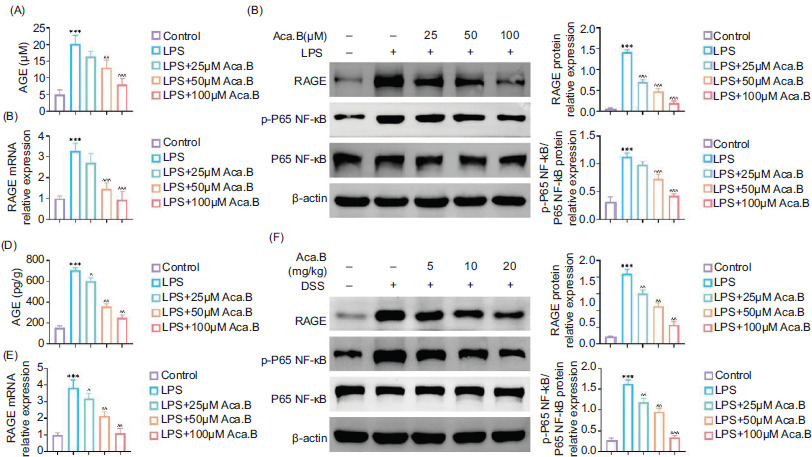
We first explored whether Aca.B affected the AGE/RAGE signaling expression under the inflammatory microenvironment to verify our hypothesis. As anticipated, LPS stimulation resulted in increased AGE accumulation and RAGE mRNA expression in MODE-K cells. However, Aca.B attenuated their upregulation in a dose-dependent manner (Figures 5A and 5B). In addition to RAGE, Aca.B further decreased the LPS-induced phosphorylated P65 NF-κB protein expression in MODE-K cells (Figure 5C). These in vitro results were consistent with the findings in DSS-induced ulcerative colitis mice. Aca.B treatment significantly attenuated AGE, RAGE, and downstream p-P65 NF-κB expression in colon samples from ulcerative colitis mice (Figures 5D and 5F). These results indicate that Aca.B inhibits the AGE/RAGE pathway in LPS-stimulated MODE-K cells and DSS-induced ulcerative colitis mice. Therefore, the inhibition of ulcerative colitis progression by Aca.B could be mediated by the AGE/RAGE pathway.
Figure 5 Aca.B inhibits the AGE/RAGE pathway in LPS-stimulated MODE-K cells and dextran sulfate sodium (DSS)-induced ulcerative colitis mice. (A) The expression of AGE in MODE-K cells treated with LPS or Aca.B. (B) The mRNA expression of RAGE in MODE-K cells treated with LPS or Aca.B as detected by qRT-PCR. (C) Expressions of RAGE, p-P65, and P65 were determined in MODE-K cells treated with LPS or Aca.B using immunoblotting analysis. The grayscale value of the corresponding band was quantitated with ImageJ. ***P < 0.001, compared with the control group. ^^P < 0.01 and ^^^P < 0.001, compared with the LPS group. (D) Expression of AGE in tissue samples from DSS-induced ulcerative colitis. (E) The mRNA expression of RAGE in tissue samples from DSS-induced ulcerative colitis as detected by qRT-PCR. (F) Expressions of RAGE, p-P65, and P65 were determined in tissue samples from DSS-induced ulcerative colitis using immunoblotting analysis. The grayscale value of the corresponding band was quantitated with ImageJ. ***P < 0.001, compared with the Sham group. ^P < 0.05, ^^P < 0.01, and ^^^P < 0.001, compared with the DSS group. Aca.B: Acanthoside B.
Aca.B Ameliorates NLRP3-mediated proptosis in MODE-K cell through inhibition of tAGE/RAGE pathway
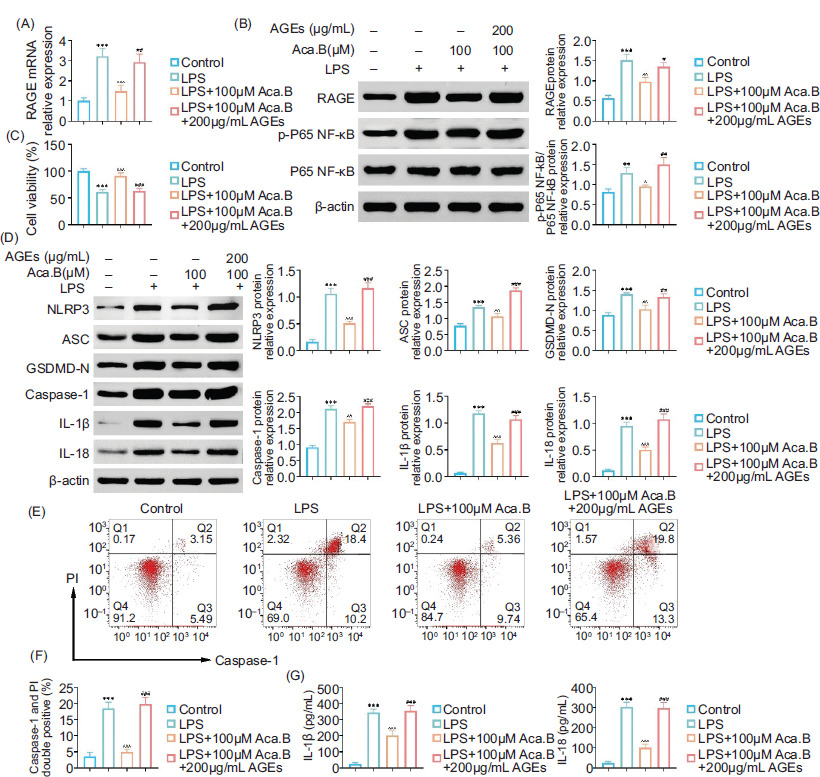
In order to verify whether the downregulation of AGE was essential for the therapeutic effect of Aca.B on inflammatory injury of epithelial cells, we added exogenous AGE (200 μg/mL) to MODE-K cells treated with LPS and Aca.B. Notably, exogenous AGE counteracted the protective effects of Aca.B on LPS-induced cell injury, evidenced by a decrease in cell viability (Figure 6B). Moreover, cells treated with both Aca.B and AGE showed higher RAGE and p-P65 NF-κB expression than those treated with Aca.B alone, indicating that AGE decreaseds inflammatory response (Figures 6A and 6C). Considering the pyroptosis pathway, AGE increased the expression of pyroptosis-specific biomarkers (NLRP3, ASC, GSDMD-N, Caspase-1, IL-1β, and IL-18) in MODE-K cells treated with Aca.B (Figure 6D). AGE further reactivated caspase-1, which was inhibited by Aca.B, increasing the caspase-1+–PI+ cell ratio from about 5% in LPS+100-μM Aca.B group to 20% in LPS+100 μM Aca.B+200 μg/mL AGE group (Figures 6E and 6F). The inhibition of IL-1β and IL-18 secretion by Aca.B was also counteracted by exogenous AGE (Figure 6G). In summary, these findings indicated that Aca.B ameliorated NLRP3-mediated proptosis in MODE-K cells by inhibiting the AGE/RAGE pathway.
Figure 6 Aca.B ameliorates NLRP3-mediated proptosis in MODE-K cell through inhibition of the AGE/RAGE pathway. (A) qRT-PCR assay showed the mRNA levels of AGE in MODE-K cells upon the indicated treatment. (B) Immunoblot analysis showed the expression of RAGE, p-P65 NF-kappaB, and P65 NF-kappaB in MODE-K cells upon the indicated treatment. (C) CCK-8 assay showed the viability of MODE-K cells upon the indicated treatment. (D) Immunoblot analysis showed the expression of NLRP3, ASC, GSDMD-N, and Caspase-1 in MODE-K cells upon the indicated treatment. (E) Flow cytometry (FCM) assay showed the apoptosis of MODE-K cells upon the indicated treatment. (F) Quantification of panel E. (G) ELISA showed the levels of IL-1β and IL-18 in MODE-K cells upon the indicated treatment. **P < 0.01 and ***P < 0.001 LPS, compared with the control group. ^P < 0.05, ^^P < 0.01, and ^^^P < 0.001, Aca.B (100 uM), compared with the LPS group. ##P < 0.01 and ###P < 0.001, Aca.B (100 uM)+AGEs (200 ug/mL), compared with the Aca.B (100 uM) group. Aca.B: acanthoside B.
Discussion
Ulcerative colitis is a chronic autoimmune disease characterized by persistent inflammatory reactions in the colon.21 The etiology of aberrant inflammatory injury remains unknown, but recent studies have suggested that pyroptosis maycan play a role.22 Our current study investigated how Aca.B could alleviate inflammatory damage and NLRP3-mediated pyroptosis of intestinal epithelial cells caused by ulcerative colitis. Additionally, we found that the protective effect of Aca.B depended on inhibiting the downstream AGE/RAGE signaling pathway. Our findings could expand the potential use of Aca.B in treating ulcerative colitis and other inflammatory conditions with distinct underlying mechanisms.
The DSS-induced colitis mice model is a well-established animal model for studying inflammatory bowel disease.23 The symptoms in the DSS model were similar to manifestations in clinical ulcerative colitis patients. In terms of pathogenesis, DSS causes acute inflammatory response independent of the acquired immune system, causing the recruitment and activation of neutrophils and macrophages as well as the overproduction of reactive oxygen species (ROS) and inflammatory mediators. The persistent inflammatory reaction results in the destruction of the colonic epithelial barrier and crypt structure, leading to bloody diarrhea.24 Herein, oral administration of Aca.B effectively reduced the severity of colitis symptoms, as evidenced by the body weight curve, DAI, and colon length. Notably, Aca.B further protected the integrity of intestinal villi with less inflammatory cell infiltration and inflammatory mediator expression. These results imply that Aca.B can inhibit inflammation and cell damage in acute stage. Considering that acute inflammatory response is a common feature in various diseases, we in the future intend to explore continuously the therapeutic effects of Aca.B on other disease models.
Aca.B is identified as the main bioactive constituent derived from the TCM herb Pogostemon cablin. Recent studies have demonstrated that Aca.B predominantly exerts anti-inflammatory effects by inhibiting the NF-κB signaling pathway in LPS-stimulated cell models.25 In the present study, our findings suggested that Aca.B could offer protection against pyroptosis independently of the inhibition of NF-κB signaling. Pyroptosis, a recently characterized form of inflammatory programmed cell death, is marked by the excessive production of a broad spectrum of inflammatory cytokines.26,27 NLRP3 inflammasome formation, cleavage of GSDMD, and activation of caspase-1 are the three hallmarks of pyroptosis initiation.28 We observed that Aca.B is capable of suppressing the expression of NLRP3, GSDMD, and caspase-1 induced by LPS, as evidenced by Western Bblot analysis.
Furthermore, the anti-inflammatory effect of Aca.B was corroborated in models of DSS-induced colitis, suggesting that the mechanism behind the inhibition of inflammation by Aca.B may be dependent on pyroptosis. To substantiate this hypothesis, further investigation into the upstream expression of AGE and RAGE was conducted both in vitro and in vivo. Previous studies demonstrated that scavenging AGE or deficiency in RAGE cancould prevent cell death and pyroptosis dependent on caspase-1 and caspase-11 in bacterial sepsis.29,30 Our results consistently showed that exogenous AGE treatment significantly reversed the pharmaceutical inhibition of pyroptosis by Aca.B in the LPS-stimulated MODE-K cell model. This suggests that the AGE/RAGE complex mightcould be the direct drug target of Aca.B. Our next step would be to further confirm the potential interaction between Aca.B and the AGE/RAGE complex.
Despite the promising results, it’s essential to acknowledge that this study has certain limitations. While animal models provide valuable insights, it may only partially reflect the complexities of human ulcerative colitis. Further studies are necessary to confirm these findings in clinical settings and to delve into the specific molecular mechanisms driving AGE/RAGE-mediated pyroptosis. Additionally, future research should explore the potential of Aca.B therapies and their safety profiles in human subjects.
Conclusions
In summary, Aca.B could ameliorate the LPS-induced cell injury and the progression of DSS-induced ulcerative colitis. The underlying mechanism is attributed to the inhibition of AGE/RAGE-mediated pyroptosis.
Competing Interests
The authors stated that they had no conflict of interest to disclose.
Data Availability
The authors declared that all data supporting the findings of this study are available in the paper, and any raw data could be obtained from the corresponding author upon request.
Author Contributions
Xiaobo He and Xiaoyan Wang designed the study and carried out the same. Xiaobo He, Chunfang Zhou, and Rui Shang supervised data collection. Xiaobo He, Chunfang Zhou, and Rui Shang analyzed the data. Xiaobo He, Chunfang Zhou, Rui Shang interpreted the data. Xiaobo He and Xiaoyan Wang prepared the manuscript for publication and reviewed its draft. All authors read and approved the final manuscript.
Conflicts of Interest
None.
Funding
None.
REFERENCES
1 Abdullah, S, Taj I, Muhammad U, Shah S, Imran A, Khalid S. A hybrid approach of vision transformers and CNNs for detection of ulcerative colitis. Sci Rep. 2024:14(1):24771. 10.1038/s41598-024-75901-4
2 Kucharzik T, Koletzko S, Kannengiesser K, Dignass A. Ulcerative colitis-diagnostic and therapeutic algorithms. Dtsch Arztebl Int. 2020;117(33–34):564–74. 10.3238/arztebl.2020.0564
3 Feuerstein JD, Isaacs KL, Schneider Y, Siddique SM, Falck-Ytter Y, Singh S. AGA clinical practice guidelines on the management of moderate to severe ulcerative colitis. Gastroenterology. 2020;158(5):1450–61. 10.1053/j.gastro.2020.01.006; 10.1053/j.gastro.2020.03.012
4 Wu X, Pan S, Luo W, Shen Z, Meng X, Xiao M, et al. Roseburia intestinalis-derived flagellin ameliorates colitis by targeting miR-223-3p-mediated activation of NLRP3 inflammasome and pyroptosis. Mol Med Rep. 2020;22(4):2695–704. 10.3892/mmr.2020.11351
5 Chen M, Ding Y, Tong Z. Efficacy and safety of Sophora flavescens (Kushen)-based traditional Chinese medicine in the treatment of ulcerative colitis: Clinical evidence and potential mechanisms. Front Pharmacol. 2020;11:603476. 10.3389/fphar.2020.603476
6 Xu M, Duan XY, Chen QY, Fan H, Hong ZC, Deng SJ, et al. Effect of compound sophorae decoction on dextran sodium sulfate (DSS)-induced colitis in mice by regulating Th17/Treg cell balance. Biomed Pharmacother. 2019;109:2396–408. 10.1016/j.biopha.2018.11.087
7 Li Y, Han Y, Wang X, Yang X, Ren D. Kiwifruit polysaccharides alleviate ulcerative colitis via regulating gut microbiota-dependent tryptophan metabolism and promoting colon fucosylation. J Agric Food Chem. 2024;72(43):23859-23874. 10.1021/acs.jafc.4c06435.
8 Karthivashan G, Kweon MH, Park SY, Kim JS, Kim DH, Ganesan P, et al. Cognitive-enhancing and ameliorative effects of acanthoside B in a scopolamine-induced amnesic mouse model through regulation of oxidative/inflammatory/cholinergic systems and activation of the TrkB/CREB/BDNF pathway. Food Chem Toxicol. 2019;129:444–57. 10.1016/j.fct.2019.04.062
9 Lee JH, Sun YN, Kim YH, Lee SK, Kim HP. Inhibition of lung inflammation by Acanthopanax divaricatus var. albeofructus and its constituents. Biomol Ther (Seoul). 2016;24(1):67–74. 10.4062/biomolther.2015.070
10 Mollace A, Coluccio ML, Donato G, Mollace V, Malara N. Cross-talks in colon cancer between RAGE/AGEs axis and inflammation/immunotherapy. Oncotarget. 2021;12(13):1281–95. 10.18632/oncotarget.27990
11 Jia C, Zhang J, Chen H, Zhuge Y, Chen H, Qian F, et al. Endothelial cell pyroptosis plays an important role in Kawasaki disease via HMGB1/RAGE/cathespin B signaling pathway and NLRP3 inflammasome activation. Cell Death Dis. 2019;10(10):778. 10.1038/s41419-019-2021-3
12 Wu X, Jilin Y, Bao X, Wang Y. Toll-like receptor 4 damages the intestinal epithelial cells by activating endoplasmic reticulum stress in septic rats. PeerJ. 2024;1212(0):e18185. 10.7717/peerj.18185
13 National Research Council Committee for the Update of the Guide for the Care and Use of Laboratory Animals. Guide for the Care and Use of Laboratory Animals, 8th ed. Washington, DC: National Academies of Sciences Press (US); 2011. Reports funded by National Institutes of Health. PMid: 21595115.
14 Chen X, Liu G, Yuan Y, Wu G, Wang S, Yuan L. NEK7 interacts with NLRP3 to modulate the pyroptosis in inflammatory bowel disease via NF-κB signaling. Cell Death Dis. 2019;10(12):906. 10.1038/s41419-019-2157-1
15 Huang MH, Han Y, Xi QH, Jin YF, Liu YL, Han YW, et al. Exploration on correlation of high DLX2 expression with poor prognosis and cellular proliferation in epithelial ovarian cancers. Eur J Gynaecol Oncol. 2021;42(3):521–9. 10.31083/j.ejgo.2021.03.2093
16 Zhang SR, Shao QM, Jia LH, Zhou F. ANXA3 regulates HIF1 alpha-induced NLRP3 inflammasome activity and promotes LPS-induced inflammatory response in bronchial epithelial cells. Signa Vitae. 2021;17(3):206–13.
17 Hou J, Hsu JM, Hung MC. Molecular mechanisms and functions of pyroptosis in inflammation and antitumor immunity. Mol Cell. 2021;81(22):4579–90. 10.1016/j.molcel.2021.09.003
18 Sun L, Ma W, Gao W, Xing Y, Chen L, Xia Z, et al. Propofol directly induces caspase-1-dependent macrophage pyroptosis through the NLRP3-ASC inflammasome. Cell Death Dis. 2019;10(8):542. 10.1038/s41419-019-1761-4
19 Du L, Ha C. Epidemiology and pathogenesis of ulcerative colitis. Gastroenterol Clin North Am. 2020;49(4):643–54. 10.1016/j.gtc.2020.07.005
20 Zeng M, Liang G, Yuan F, Yan S, Liu J, He Z. Macrophages-derived high-mobility group box-1 protein induces endothelial progenitor cells pyroptosis. iScience. 2024;27(10):110996. 10.1016/j.isci.2024.110996
21 Burri E, Maillard MH, Schoepfer AM, Seibold F, Van Assche G, Rivière P, et al. Treatment algorithm for mild and moderate-to-severe ulcerative colitis: An update. Digestion. 2020;101(Suppl 1):2–15. 10.1159/000504092
22 Zhen Y, Zhang H. NLRP3 inflammasome and inflammatory bowel disease. Front Immunol. 2019;10:276. 10.3389/fimmu.2019.00276
23 Bauer C, Duewell P, Mayer C, Lehr HA, Fitzgerald KA, Dauer M, et al. Colitis induced in mice with dextran sulfate sodium (DSS) is mediated by the NLRP3 inflammasome. Gut. 2010;59(9):1192–9. 10.1136/gut.2009.197822
24 Mei Y, Fang C, Ding S, Liu X, Hu J, Xu J, et al. PAP-1 ameliorates DSS-induced colitis with involvement of NLRP3 inflammasome pathway. Int Immunopharmacol. 2019;75:105776. 10.1016/j.intimp.2019.105776
25 Kim MJ, Wang HS, Lee MW. Anti-inflammatory effects of fermented bark of Acanthopanax sessiliflorus and its isolated compounds on lipopolysaccharide-treated RAW 264.7 macrophage cells. Evid Based Complement Alternat Med. 2020;2020:6749425. 10.1155/2020/6749425
26 Fang Y, Tian S, Pan Y, Li W, Wang Q, Tang Y, et al. Pyroptosis: A new frontier in cancer. Biomed Pharmacother. 2020;121:109595. 10.1016/j.biopha.2019.109595
27 Yu P, Zhang X, Liu N, Tang L, Peng C, Chen X. Pyroptosis: Mechanisms and diseases. Signal Transduct Target Ther. 2021;6(1):128. 10.1038/s41392-021-00507-5
28 Kovacs SB, Miao EA. Gasdermins: Effectors of pyroptosis. Trends Cell Biol. 2017;27(9):673–84. 10.1016/j.tcb.2017.05.005
29 Chen L, Zhao Y, Lai D, Zhang P, Yang Y, Li Y, et al. Neutrophil extracellular traps promote macrophage pyroptosis in sepsis. Cell Death Dis. 2018;9(6):597. 10.1038/s41419-018-0538-5
30 Deng M, Tang Y, Li W, Wang X, Zhang R, Zhang X, et al. The endotoxin delivery protein HMGB1 mediates caspase-11-dependent lethality in sepsis. Immunity. 2018;49(4):740–53.e7. 10.1016/j.immuni.2018.08.016