
Download
ORIGINAL ARTICLE
Analysis of B cell proliferation in response to in vitro stimulation in patients with CVID
Velásquez-Ortiz Ma. Guadalupea, Saldaña-Solano Abigail Jb, O’Farrill-Romanillos Patriciac, Herrera-Sánchez Diana Andreac, Scheffer-Mendoza Selmad, Yamazaki-Nakashimada Marco Ad, Zarate-Hernández Ma. Carmene, Espinosa-Padilla Sara Ef, Laura Berrón-Ruizf*
aPosgrado en Ciencias Biológicas UNAM, Ciudad Universitaria, México
bDepartamento de Atención de la Salud, Universidad Autónoma Metropolitana, Unidad Xochimilco, Ciudad de México, México
cServicio de Alergia e Inmunología Clínica, Hospital de Especialidades del Centro Médico Siglo XXI, IMSS
dServicio de Inmunología y Alergia, Instituto Nacional de Pediatría SSA, Ciudad de México, México
eServicio de Alergia, Hospital Universitario, Monterrey, Nuevo León
fLaboratorio en Inmunodeficiencias, Instituto Nacional de Pediatría SSA, Ciudad de México, México
Abstract
Background: Common variable immunodeficiency (CVID) is a heterogeneous disorder characterized by defective antibody production and impaired differentiation of B cells. B cell proliferation is an essential step for antibody synthesis. Depending on the nature of the stimulus, their response may be either T-cell-dependent or T-cell-independent.
Methods: We studied 23 CVID patients and 14 healthy donors (HD). The patients were categorized based on their percentage of memory B cells. In addition to standard immunophenotyping of circulating human B and T cell subsets, an in vitro CFSE dilution assay was used to assess the proliferative capacity of B cells and to compare the activation of the T cell-dependent and T cell-independent response among the patients.
Results: Patients with a reduction in memory B cells exhibited an increase in follicular T cells (Tfh) and showed low proliferation in response to PKW, CpG, and SAC stimuli (Condition II) (p= 0.0073). In contrast, patients with a normal percentage of memory B cells showed a high expression of IL-21R and low proliferation in response to CPG (Condition III); IL-21, CD40L, and anti-IgM (Condition IV) stimuli (p= 0.0163 and p = 0.0475, respectively).
Conclusion: Defective proliferation in patients depends on the type of stimulus used and the phenotypic characteristics of the patients. Further studies are necessary to understand the disease mechanisms, which may guide us toward identifying genetic defects associated with CVID.
Key words: B cells proliferation, CVID, T-dependent response, T-independent response
*Corresponding author: Laura Berron-Ruiz, Laboratorio en Inmunodeficiencias, Instituto Nacional de Pediatría SSA, Ciudad de México, México. Email address: [email protected]
Received 12 April 2024; Accepted 29 July 2024; Available online 1 September 2024
Copyright: Velásquez-Ortiz G, et al.
License: This open access article is licensed under Creative Commons Attribution 4.0 International (CC BY 4.0). http://creativecommons.org/licenses/by/4.0/
Introduction
Common variable immunodeficiency (CVID) is the most prevalent primary immunodeficiency, now referred as inborn errors of immunity, and is characterized by antibody deficiency due to B cell dysfunction. CVID is defined by decreased serum immunoglobulin levels and impaired humoral immune responses, leading to increased susceptibility to respiratory infections. Patients also present with various clinical manifestations, such as gastrointestinal, autoimmune, lymphoproliferative, and malignancies.1 Over the past years, numerous studies have reported abnormalities in both the innate and adaptive immune systems in CVID patients. Disturbances in T and Natural killer (NK) cell homeostasis have been reported,2 with decreased circulating NK cell numbers associated with severe bacterial infections and granulomas.3 In addition, CD4+ T cells lymphopenia has been observed, along with reduced counts of naïve CD4+ T cells, regulatory T cells (Treg), and an increase in circulating memory CXCR5+ CD4+ T cells.4,5 T cells in CVID patients also exhibit lower proliferative capacities and abnormal cytokine production.6
CVID is a primary antibody defect, hence attention is naturally focused on B cells analysis. Immunophenotyping of B cell compartments is a routine evaluation for patients with CVID. A low percentage of switched memory B cells is an abnormality present in 75% of these patients.7 Classifying the subpopulations of B cells in CVID patients help define subcategories, as patients can present with varying B cell numbers and reduced frequencies of isotype-switched memory B cells and plasma cells (PCs), which correlate with clinical manifestations.9,10 The association between mutations in specific genes and the development of CVID is well established in only 2–10% of cases. Whole-genome or exome sequencing efforts have led to the discovery of novel mutations in genes associated with CVID. Although a few genetic mutations may contribute to the condition, the etiology remains unknown in most cases.11
Many defects in B cells have yet to be fully characterized through functional analysis. B cell proliferation is an essential step in the chain of events leading to antibody synthesis. For B cell responses to progress, proliferation must occur to generate a clonal population of daughter cells and facilitate differentiation. This proliferation can proceed primarily through two distinct pathways, depending on the nature of the stimulus, that is, T cell-dependent antigens, typically protein structures, or T cell--independent antigens, typically carbohydrates.12
After encountering an antigen, B cells proliferate in response to signals derived from helper T cells in T cell--dependent (TD) responses. T cell-dependent differentiation leads to the development of long-lived, highly specific, Ig-producing PCs. Activation results in the formation of germinal centers in lymphoid organs such as the lymph nodes, spleen, and Peyer’s patches, where activated CD4+ T cells interact with activated B cells through direct CD40L–CD40 interactions and the release of soluble factors such as IL-21.13 These T–B cell interactions are essential for somatic hypermutation and CSR in B cells, both of which are necessary to induce differentiation into high-affinity IgG-producing PCs.14 In some cases, strong signaling through co-receptor (such as TLRs) can overcome the need for T cell help, resulting in a T cell-independent (TI) response. T cell-independent activation most often leads to the generation of short-lived, moderate-affinity Ig-producing PCs. When activated in a T cell-independent manner, activated B cells mainly differentiate into IgM-producing PCs, as observed with polyclonal activation either through BCR-independent TLR activation or BCR-dependent carbohydrate antigens.15,16 For this reason, we have developed a cell culture system to determine the capacity of mature peripheral blood B cells to proliferate in CVID patients.
Using this system, we classified platelets by their percentage of memory B cells in peripheral blood and demonstrated differences between two groups of patients. The results help target molecular defects associated with the clinical CVID phenotype. This functional assay is a valuable tool for evaluating B cell function and provides a more precise characterization of CVID patients.
Methods
Patients
The patients were diagnosed at medical units with greater resolution capacity within the health system, and by specialized doctors following criteria suggested by the European Society for Immunodeficiencies (ESID) and the Pan-American Group for Immunodeficiencies (PAGID).17 These criteria include patients who have a marked decrease (at least 2 SD less than the mean for age) in at least one of the major isotypes (IgM, IgG, and IgA) and meet all of the following conditions: onset of immunodeficiency at an age older than 2 years, absence of isohemagglutinins and/or poor response to vaccines, and exclusion of other defined causes of hypogammaglobulinemia. Twenty-three patients diagnosed with CVID were selected for the study. Patients with hypogammaglobulinemia and a variable number of B cells (>2%) secondary to medications or other immunodeficiencies, and those with well-known single-gene defects, were excluded. All selected patients received IVIG replacement therapy at doses 400-600 mg/kg/dose. Peripheral blood samples were collected before IVIG infusion from patients who did not have severe infection. Participants were provided with an informed consent letter, which they read, accepted, and signed. This study was approved the local ethics number 27/2026 (INP) and was conducted in accordance with the ethical guidelines of the 1975 Declaration of Helsinki.
Data collection and patient classification
Demographic, clinical, and immunological data were collected for each patient. Clinical severity was assessed using a CVID severity score proposed by Mokhtari et al.8 which provides numerical input on clinical manifestations. Patients were classified into two groups based on the percentage of isotype-switched memory B cells in the peripheral blood: Group 1, with a reduced percentage of isotype-switched memory B cells (<0.4% of the total lymphocyte population), and Group 2, with a normal percentage of isotype-switched memory B cells (>0.4% of the total lymphocyte population), as described by Warnatz et al.9
Lymphocyte population and subpopulation phenotyping
Flow cytometric analysis of CVID patients was performed before IVIG infusion to evaluate different populations and subpopulations of lymphocytes. Cell phenotyping was conducted using whole-blood samples with an anticoagulant (Acid Citrate/Dextrose BD Vacutainer®). Lymphocyte populations were stained with the following mixtures of monoclonal antibodies (mAbs): anti-CD45-FITC/anti-CD14-PE, anti-CD3-FITC/anti-CD19-PE/anti-CD45-PerCP, anti-CD4-FITC/anti-CD8-PE/anti-CD3-PerCP, anti-CD3-FITC/anti-CD16+56/anti-CD45-PerCP. To detect naïve (CD45RA+) and memory (CD45RO+) T cells, the following antibodies were used: anti-CD45RO-PE/anti-CD45RA-FITC/anti-CD3-PerCP/anti-CD4-APC. Circulating memory T cells were analyzed with the following antibody mixture: anti-CD45RA-FITC/anti-CD4-APCCy7/anti-CXCR5-APC/anti-CD3-PC5. To identify B cell, populations were stained with a mixture of anti-CD19-APC/anti-IgD-FITC/anti-CD27-PE, anti-CD19-APC/anti-CD38-FITC/anti-CD24-PE, and anti-CD19-APC/anti-CD38-FITC/anti-CD21-PE. The regulatory T-cell population was assessed indirectly using the combination of antibodies anti-CD4-APC/anti-CD25-PE/anti-CD127-FITC. Finally, the IL-21 receptor on B cells was determined with anti-CD19-APC/anti-CD27-FITC/anti-IL21R-PE. All antibodies were purchased from BD Biosciences, San Diego, CA, USA. Samples were incubated for 30 min at room temperature in the dark. After incubation, erythrocytes were lysed by adding FACS lysing solution (BD Biosciences) for 10 min. The cells were then washed with PBS and fixed in PBS containing 1% formalin.
Proliferation assay
PBMCs were isolated from blood with anticoagulant using a density gradient centrifugation with Histopaque®-1077 (Sigma-Aldrich, St Louis, MO, USA). To track cell divisions, cells were labeled before culture with carboxyfluorescein diacetate and succinimidyl ester (CellTraceTM CFSE, Life Technologies) according to the manufacturer's protocol. PBMCs were adjusted to match the number of B cells in each culture from patients and controls. Cells were seeded at a density of 1×106 B cells/mL in RPMI supplemented in 96-well flat-bottom culture plates under five conditions for 5 days. The condition I non-stimulated. Condition II contained three stimuli: 3.85 µL/mL CpG (ODN 2006, Invivogen), 1:5,000 protein A from Staphylococcus aureus Cowan SAC (Merck) and 1 µL/mL pokeweed mitogen (PWM, Sigma Aldrich). Condition III contained 3.85 µL/mL CpG. Condition IV contained 80 ng/mL recombinant human IL-21 (R&D System), 1 µL/mL soluble CD40 ligand (sCD40L, Cell Signaling Technology®), and 1 µL/mL unconjugated goat anti-human IgM F(ab’)2 fragments (Invitrogen). Condition V contained 1 µL/mL soluble CD40 ligand, 1 µL/mL unconjugated goat anti-human IgM F(ab’)2 fragments. After culturing the cells, they were washed with PBS and labeled with anti-CD19-APC (BD Biosciences). Samples were incubated for 30 min at room temperature in the dark. After incubation, cells were washed with PBS and fixed in PBS containing 1% formalin.
Statistical analysis
Fisher’s exact test was used to evaluate classified variables and assess their independence from clinical parameters, as well as to find the association (contingency) between the clinical manifestations of patients in Group 1 (with memory B cells) and patients in Group 2 (without memory B cells). This test determined whether the proportions of a variable differed depending on the value of another variable. Differences between groups were analyzed using the Mann–Whitney U test (GraphPad Prism v8). Statistical significance is indicated as follows * for p ≤ 0.05, **p ≤ 0.01, ***p ≤ 0.001, and ****p-value ≤ 0.0001.
Results
Clinical, demographic, and immunologic characteristics of CVID patients
The study included 23 patients diagnosed with CVID, consisting of 6 male and 17 female patients. The mean age of the patients was 36±17 years old. Regarding the clinical characteristics, the most prevalent were sinusitis (78.3%), followed by lymphadenopathy and autoimmune diseases (56.5%), pneumonia and enteropathies (47.8%), and bronchiectasis (39.1%). Group 1, which had the highest frequency of clinical manifestations, showed significant differences compared to Group 2 in infectious processes such as sinusitis and pneumonia (p = <0.0001) and bronchiectasis (p=0.0286). In addition, Group 1 experienced inflammatory complications such as granulomas and enteropathies (p= <0.0001), as well as solid tumors (p = 0.0068). Among the 10 patients with autoimmune manifestations, 30 % were diagnosed with Evans syndrome, 50 % with immune thrombocytopenic purpura (ITP), and 20 % had two autoimmune diseases, one patient with Evans syndrome and cutaneous discoid lupus erythematosus, and another with ITP and mixed connective tissue disease (MCTD).
The mean clinical severity score of the cohort was 6.8. The severity was higher in Group 1 (7.7) compared to Group 2 (5.6), but the difference was not statistically significant. In all patients, serum levels of immunoglobulins IgG, IgA, and IgM were below the reference values. A significant difference was observed in IgG values (p = 0.0344), with the mean IgG level being higher in Group 2. No significant differences were found for IgM and IgA levels; however, patients in Group 2 had higher mean values for IgA (35.64) and IgM (63.9) compared to Group 1 (4.9 and 61.5, respectively).
After performing phenotyping of cell populations and subpopulations and comparing these results between patient groups, significant differences were observed. CD19+ B cells (p=0.0015) were found at lower levels in Group 1. Similarly, there were differences found in the percentages of isotype-switched memory B cells (p=0.123), which were decreased in Group 1. CD21 low B cells were found to be increased in Group 1(p = 0.0400). In addition, CD4+CD45RO+ T cells were found to be increased in Group 1 patients (p = 0.0474), as detailed in Table 1.
Table 1 Clinical and immunologic features of total CVID patients, grouped by their memory B cells frequency.
| Total CVID | CVID group 1 <0.4%** | CVID group 2 >0.4%* | CVID group 1 vs. CVID group 2 p-value*** | |
|---|---|---|---|---|
| Number | 23 | 13 | 10 | |
| Sex (female/male) | 17/6 | 8/5 | 9/1 | |
| Age (mean±SD) | 36.8±17.1 | 33.5±17.1 | 38.7±18.3 | |
| Clinical manifestations (number of cases (%)) | ||||
| Pneumonia | 11 (47.8) | 8 (61.5) | 3 (30.0) | <0.0001 |
| Sinusitis | 18 (78.3) | 12 (92.3) | 6 (60.0) | <0.0001 |
| Bronchiectasis | 9 (39.1) | 6 (46.2) | 3 (30.0) | 0.0286 |
| Splenomegaly | 7 (30.4) | 4 (30.8) | 3 (30.0) | >0.9999 |
| Lymphadenopathy | 13 (56.5) | 7 (53.8) | 6 (60.0) | 0.4752 |
| Granulomas | 3 (13.0) | 3 (23.1) | 0 (0.0) | <0.0001 |
| Enteropathies | 11 (47.8) | 8 (61.5) | 3 (30.0) | <0.0001 |
| Autoimmune diseases | 10 (43.5) | 6 (46.2) | 4 (40.0) | 0.4752 |
| Cancer (solid tumors) | 1 (4.3) | 1 (7.7) | 0 (0.0) | 0.0068 |
| Severity score (mean±SD) | 6.8±4.7 | 7.7±5.7 | 5.6±2.9 | 0.4144 |
| Immunoglobulin levels mg/dL (means±SD) | ||||
| IgG | 285.0±187.4 | 210.1±148.2 | 378.5±187.0 | 0.0344 |
| IgA | 19.0±31.5 | 4.9±7.0 | 35.64±39.7 | 0.0627 |
| IgM | 62.5±120.2 | 61.5±148.2 | 63.9±66.4 | 0.1521 |
| Leukocytes populations (absolute cells count means/mm3±SD) | ||||
| Total leukocytes | 6563.1±2582.2 | 6576.9±2538.7 | 6545.0±2775.6 | 0.9758 |
| Lymphocytes populations (absolute cells count means/mm3±SD) | ||||
| Total lymphocytes | 1770.2 ± 950.1 | 1714.4±1023.6 | 1842.7±893.9 | 0.7381 |
| T lymphocytes CD3+ | 1294.2±766.1 | 1391.3±941.7 | 1167.7±469.6 | 0.7381 |
| T lymphocytes CD4+ | 597.8±388.5 | 605.2±464.1 | 588.1±285.4 | 0.5629 |
| T lymphocytes CD8+ | 641.2±610.3 | 740.3±765.1 | 512.3±311.8 | 0.3758 |
| NK lymphocytes CD16+56+ | 200.3±182.4 | 199.4±181.7 | 201.3±193.0 | 0.9758 |
| B lymphocytes CD19+ | 261.3±324.1 | 108.1±89.0 | 460.4±410.2 | 0.0015 |
| T CD4+ cell subpopulations (percentage means) | ||||
| T lymphocytes CD4+CD45RA+ | 24.04 | 18.69 | 31.00 | 0.0794 |
| T lymphocytes CD4+CD45RO+ | 72.09 | 79.00 | 63.10 | 0.0474 |
| B cells subpopulations (percentage means) | ||||
| Naive B cells IgD+ CD27- CD19+ | 76.60 | 80.92 | 71.00 | 0.2130 |
| Not-switched B cells IgD+ CD27+ CD19+ | 7.24 | 5.75 | 9.20 | 0.2745 |
| Switched B cells IgD- CD27+ CD19+ | 6.55 | 4.37 | 9.40 | 0.0123 |
| Transitional B cells CD38+CD24+CD19+ | 6.08 | 6.43 | 5.60 | 0.3467 |
| CD21low B cells CD38-CD21lowCD19+ | 18.54 | 24.15 | 11.20 | 0.0400 |
| Plasmablast CD38+CD19+ | 0.55 | 0.51 | 0.60 | 0.5395 |
*Represented patients contain an isotype-switched memory B cell fraction of above 0.4% of total lymphocytes.
**Represented patients containing an isotype-switched memory B cell fraction of below 0.4% of total lymphocytes.
Fisher exact test were used to analyzed clinical manifestations and U Mann-Whitney to immunological phenotypes.
***P-value significant at the level <0.05. Significant values are represented in bold.
CVID patients grouped by their percentage of memory B cells show differences in the expression of T cell subpopulations, such as T CXCR5+ cells and Treg, as well as in the percentage of IL-21R expression in B cells
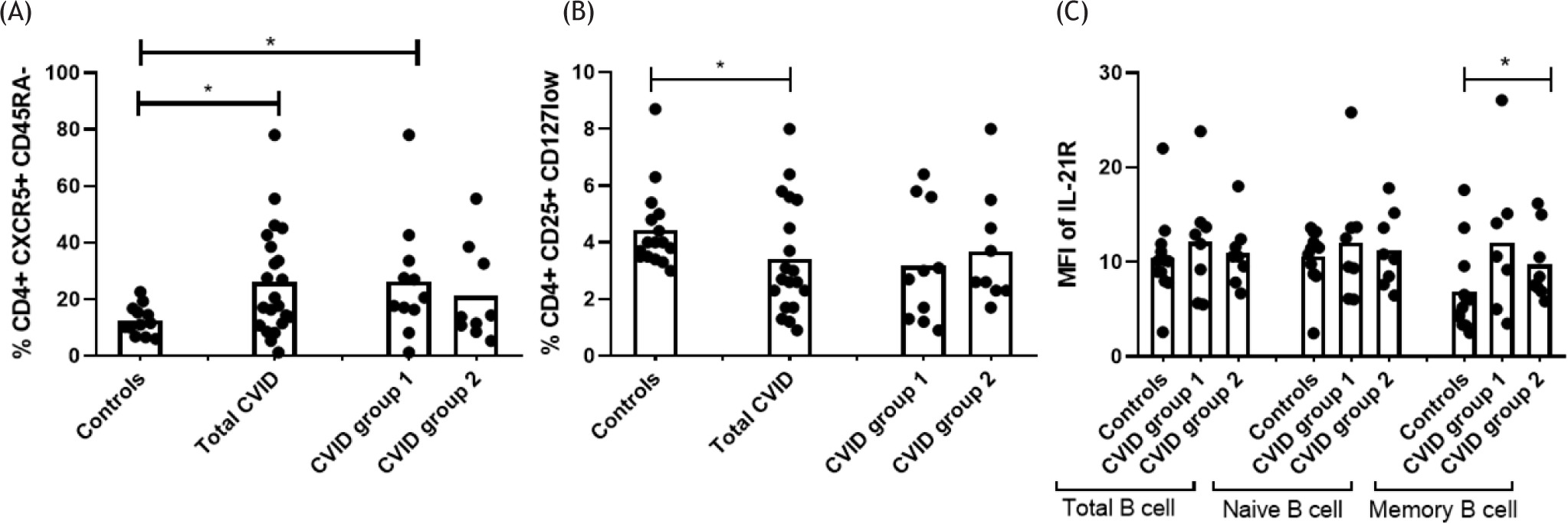
After analyzing the CD4+ CXCR5+ CD45RA- cell population, it was observed that patients with CVID had higher percentages compared to controls (p=0.0336). Similarly, when classified by group, Group 1 showed increased percentages of CD4+ CXCR5+ CD45RA- population (p = 0.0225) (Figure 1A). In addition , patients with CVID showed lower percentages of CD4+ CD25+ CD127low T cells (p = 0.0396) (Figure 1B), although this difference was not significant when patients were further divided into groups. IL-21 receptor expression in B cells was analyzed to determine if there were differences between control and patient groups. While total B cells, naive B cells, and memory B cells were analyzed, significant differences were found only in the latter. It was observed that patients with CVID in Group 2 have a higher expression of IL-21R compared to controls (p = 0.0473) (Figure 1C).
Figure 1 Comparison of specific subpopulations and IL-21R expression, grouping patients by Freiburg classification. (A) Percentages of CD4+ CXCR5+ CD45RA- population are compared between controls (n =12), patients with CVID (n =20), and patients with CVID classified into Groups 1 (n = 11) and 2 (n = 9). (B) Percentages of CD4+ CD25+ CD127low populations are compared between controls (n =16), CVID patients (n = 19), and CVID patients classified in Groups 1 (n =10) and 2 (n = 9). (C) IL-21R expression in total, naïve, and memory B cells are compared between controls (n = 12) and CVID patients classified into Groups 1 (n = 8) and 2 (n = 8). Data are presented as means. Statistical differences between patients and controls were compared using the Mann-Whitney U test. (*) Significant, p = 0.05; (**) very significant, p = 0.01; and (***) highly significant, p = 0.001. CVID, common variable immunodeficiency; MFI, mean fluorescent intensity.
CVID patients show differences in vitro activation with TD and TI stimulation
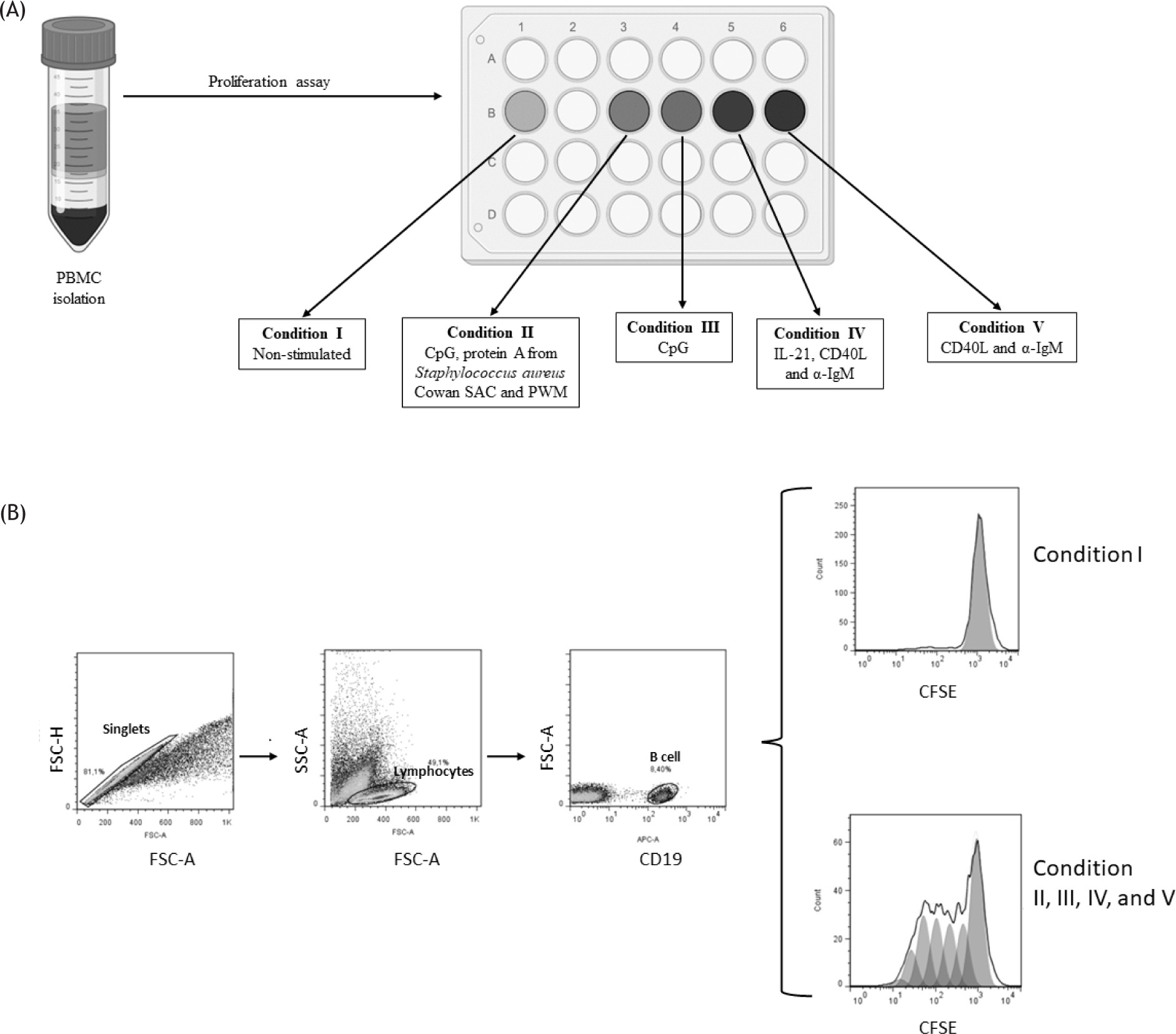
To study the functions of the B-cell compartment in vitro, we activated B cells with either TI and/or TD signals. Anti-IgM, CpG ODN-2006, and pokeweed mitogen (PWM) provide both TI and TD signals. CPG, a synthetic oligonucleotide that binds to TLR9, was chosen as a representative TI stimulus, whereas the TD signals were mimicked by engaging CD40, IL-21, and IgM on B cells. PBMCs were cultured under different conditions, as shown in Figure 2A. Conditions II to V involved activation with different mitogens, most of which were specific for B cell proliferation, while Condition I served as the non-stimulated control. After obtaining the proliferation results, the analysis was performed as follows: first, the singlets region was selected, then the lymphocyte population expressing CD19 (indicating B cells) were identified, and finally, peak 0 of proliferation, corresponding to the condition I, was established as a reference point. This reference point was used to determine the percentage of division in the stimulated conditions (II to V) as shown in Figure 2B.
Figure 2 The proliferation of B cells under various conditions of activation and analysis. (A) Schematic representation of the experimental approach. PBMCs were isolated from the blood of controls and CVID patients, stimulated under the indicated conditions and analyzed at day 5 by flow cytometry. (B) Analysis of B cells proliferation by flow cytometry. Cells were pre-gated on FSC-A/FSC-H to identify singlets and on FSC-A/SSC-A to identify lymphocytes. The CD19+ population was then selected to identify B cells, and the split percentage was determined from the MFI of CFSE, using the FlowJo v7.6 proliferation tool.
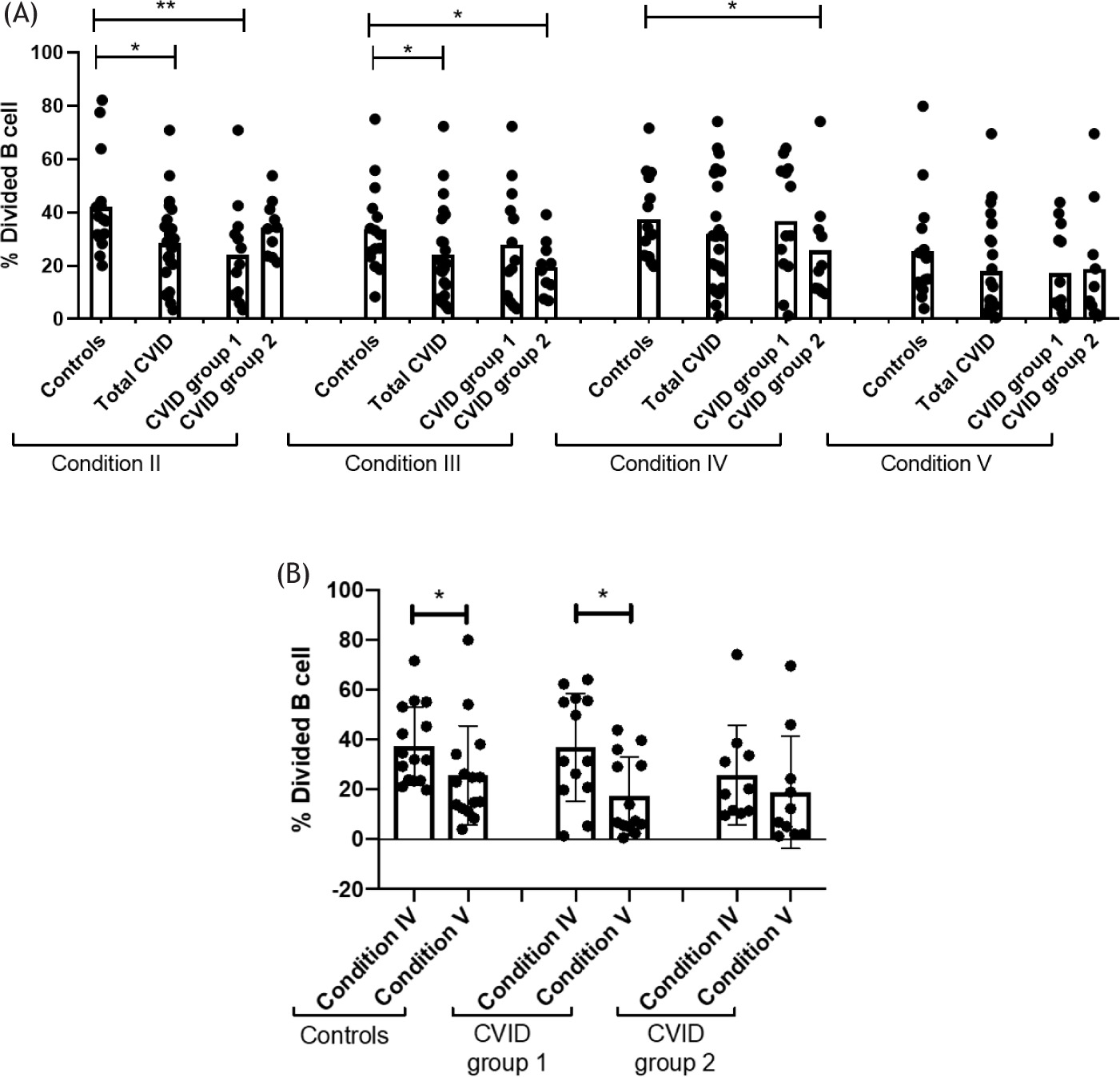
From the results obtained from B cell proliferation, decreased proliferation was observed in total CVID patients in Condition II (p = 0.0263) and Condition III (p = 0.0073). When patients were divided into different groups, it was observed that patients in Group 1 exhibited decreased B cell proliferation in Condition II (p = 0.0073). In contrast, patients in Group 2 presented decreased proliferation in Condition III (p = 0.0163) and Condition IV (p = 0.0475). No differences in proliferation were observed between patients and controls in Condition V (Figure 3A).
Figure 3 (A) Comparison of proliferation between controls (n = 15), CVID patients (n = 23), and CVID patients classified in Groups 1 (n = 13) and 2 (n = 10) under different activation conditions. Data are presented as means. (B) Comparison of proliferation between conditions IV and V in the groups of controls, CVID patients in Group 1 and Group 2. Statistical differences between patients and controls were compared using the U Mann-Whitney test. (*) Significant, p = 0.05; (**) very significant, p = 0.01; and (***) highly significant, p = 0.001. CVID = Common Variable Immunodeficiency; MFI = mean fluorescent intensity.
An analysis was performed between Conditions IV and V to determine the effect of adding IL-21 to the anti-CD40 and anti-IgM stimuli on B cell proliferation. This addition resulted in increased proliferation in B cells for both Group 1 patients and controls (p = 0.0329; p = 0.0338, respectively). However, in Group 2 patients, there was no significant increase in proliferation (Figure 3B).
Discussion
CVID is a complex and heterogeneous disease characterized by a wide spectrum of infectious and non-infectious clinical manifestations. The results of our study confirm the diverse clinical courses of CVID. The clinical phenotype demonstrated in our patients was similar to that reported in other analyzed CVID populations.18 Recurrent upper and lower respiratory tract infections were predominant in 80% of patients, with pneumonia and sinusitis diagnosed in 47.8% and 78.3% of patients, respectively. At the time of diagnosis, immunoglobulin levels in the main classes were very low in most of the presented patients. In addition, many of our CVID patients also experienced inflammatory complications, emphasizing the complex interplay between infection and immune dysregulation. The patient cohort was divided based on the percentage of memory B cells: Group 1 included patients with a decreased percentage of memory B cells, While Group 2 consisted of those with a normal percentage of memory B cells.
Group 1 patients have a higher clinical severity score compared to Group 2. This is because they present a greater number of infectious manifestations in the respiratory tract, such as pneumonia and sinusitis, as well as a higher incidence of enteropathies and inflammatory conditions, such as bronchiectasis and granulomas. Although other cohorts of CVID patients grouped by memory B cells percentage have shown that Group 1 typically has more patients with autoimmune manifestations, our cohort did not reveal significant differences in autoimmune processes between the two groups.19,20 Notably, our cohort consists mostly of older adults, with a mean age of 37 years, and a high percentage of patients with autoimmune diseases. The advanced age of our patients could contribute to the increased prevalence of non-infectious complications, such as autoimmune diseases. On the other hand, the IgG immunoglobulin concentration of Group 1 had a significant difference compared to Group 2. As for the other immunoglobulin isotypes, there was no significant difference in their concentration. This difference may be due to the decreased number of memory B cells in Group 1. In addition, extremely low levels of immunoglobulins, of all or some major classes, are associated with a higher risk of complications, e.g., pneumonia and meningitis were more often associated with low levels of immunoglobulin G.21
In the analysis of the immunological phenotype of lymphocyte populations and subpopulations, a significant difference was observed between the groups. Group 1 had a decrease in both total B cells and switched memory B cells compared to Group 2 patients. Additionally, Group 1 showed an increase in CD21low B cell populations, with a significant difference from Group 2. Various studies have reported that cohorts of CVID patients with a decreased percentage of total and memory B cells also show an increase in populations of CD21lowB cells. The expansion of CD21low B cells is a manifestation of chronic activation and is significantly associated with splenomegaly and granulomatous diseases.10,22 In our cohort, this population was clinically associated with granulomatous disease only in Group 1. Another notable difference in this cohort was the increased number of memory CD4+ CD45RO+ T cells in Group 1 compared to Group 2. Interestingly, both our Group1 patients and those observed in other studies show a severe reduction in naïve CD4+ T cells and an increase in memory CD4+ T cells, alongside a reduction in class-switched memory B cells and an expansion of CD21low B cells. Additionally, splenomegaly and granulomatous disease have been noted.23
Although B cell subpopulations are notably affected in most CVID patients, abnormalities in T cell subpopulations have also been reported. T cells in CVID patients often show lower proliferative capacities and abnormal cytokine production.6,24 Regulatory T cells (Tregs) and follicular T helper (Tfh) cells are also reported to be abnormal in CVID patients with non-infectious complications.25 In CVID, it is known that many patients experience a decrease in memory B cell populations. Recent studies have shown that follicular helper (Tfh) cells, which are major CD4+ T helper subsets, are essential for B cell differentiation into plasma cells and for the generation of memory B cells in germinal center (GC).26 Tfh cells are characterized by the expression of CXCR5 and the production of cytokine IL-21, which supports B cell differentiation via the IL-21 receptor (IL-21R). The role of Tfh and Treg cells in antibody-mediated autoimmune diseases is well-established in humans.27 An increase in THF cells and a reduction in Treg have been observed in CVID patients with autoimmune diseases and a deficiency of switched memory B cells.19,26,28 In our study, we examine these T cell population in CVID patients. We found that patients had a higher percentage of Tfh cells compared to controls, with Group 1 showing a significant difference from the controls. This may be due to the high percentage of inflammatory manifestations in this cohort. Although the percentage of Tregs were significantly low in the overall CVID patient population compared to controls, there was no significant difference between Groups 1 and 2. This lack of difference could be due to both groups having a similar percentage of patients with autoimmune diseases. We also measured IL-21R expression to evaluate its role in the IL-21/STAT3 signaling pathway, which is crucial for B cell proliferation and subsequent differentiation. Interestingly, we observed a significant increase in IL21R expression in memory B cells in Group 2 compared to controls. The IL-21R is highly expressed on naïve B cells and its expression is lower on memory B cells.29 This behavior has not been previously evaluated in CVID patients. The observation that memory B cells from Group II patients express more IL-21R compared to naïve B cells from the same group could indicate defects in the IL-21/STAT3 pathway activation. Therefore, further research is necessary, as the high expression of this receptor could impair the terminal differentiation to plasma cells and lead to deficiencies in serum immunoglobulin secretion.
We showed that the functional differentiation of mature human B cells compartment in vivo can be effectively studied using a standard in vitro culture system through the assessment of B cell proliferation. Our results reveal distinct differences between CVID patients groups when stimulated under T-dependent and T-independent conditions. Specifically, we observed reduced proliferation in Groups 1 and 2 patients under condition II (CpG, PWM, Cowan), III (CpG only) and IV (IL-21, CD40, α-IgM) compared to control subjects. The stimuli used in the different conditions activate the B cells through several pathways. Regardless of the specific condition used, B cells of CVID patients have a lower percentage of proliferation rates compared to controls, with significant differences observed. One of the critical issues in CVID patients is that the B cells do not become completely activated. The B cells not only fail to proliferate normally but also exhibit defective differentiation into memory B cells or plasma cells.
In our study, we found of that patients in Group 1 exhibited reduced proliferation in condition II compared to both Group 2 and controls. This suggests that Group 1 patients may have defective BCR triggering, leading to impaired T cell-dependent germinal center response. Conversely, Group 2 patients showed decreased proliferation in the stimuli of conditions III and IV. TLR-9 triggering by CpG-DNA activates B cells with or without ligation of the BCR, mediates T-cell independent. Recent studies have highlighted that B cells of CVID patients exhibit defects in TLR-9 signaling, irrespective of a lack of memory B cells. These defects would lead to reduce B cell proliferation, and reduced IgG class switching.30,31 Conditions IV and V were designed to show the ability of IL-21 to increase proliferation. We observed that there was an increase in proliferation and was evidenced in Group 1 and control group. However, Group 2 patients did not show an increase in proliferation with IL-21 supplementation. This indicates that Group 2 may have defects in signaling pathways important for terminal differentiation in the germinal center; this would partly explain that even if they have a normal percentage of memory B cells, they fail to optimally secrete serum immunoglobulins.
Our study had the limitation that the sample size of each patient group was relatively small. Given that CVID is a rare disease, our sample size is similar to that of other single-center studies. Future research with larger cohorts will be necessary to confirm these findings. We demonstrated defects in the expression percentages of T cells subpopulations, such as Tfh, and Treg cells, as well as in B cells the expression of IL-21R, which together with the findings of proliferate assay functional, may contribute to understand the defect in B cells in CVID patients. CVID patients showed functional differences in T-dependent response activation and T-independent. These results are important for further defining and characterizing this heterogeneous disease.
Acknowledgments
M.G. Velásquez-Ortiz, MSc, is a student who received a graduate scholarship from CONACHyT, México. This work was partially supported by the Consejo Nacional de Ciencia y Tecnología (CONACHYT, México) grant CB # 256471. This paper is part of the MSc degree at the Posgrado en Ciencias Biológicas, UNAM.
Conflict of Interest
The authors declare no conflicts of interest. The authors alone are responsible for the content and writing of the paper.
REFERENCES
1. Ameratunga R, Allan C, Woon ST. Defining common variable immunodeficiency disorders in 2020. Immunol Allergy Clin North Am. 2020; 40(3):403–20. 10.1016/j.iac.2020.03.001
2. Wiesik-Szewczyk E, Jahnz-Rózyk K. From infections to autoimmunity: Diagnostic challenges in common variable immunodeficiency. World J Clin Cases. 2020; 8(18):3942–55. 10.12998/wjcc.v8.i18.3942
3. Ebbo M, Gérard L, Carpentier S, Vély F, Cypowyj S, Farnarier C, et al. Low circulating natural killer cell counts are associated with severe disease in patients with common variable immunodeficiency. EBioMedicine. 2016; 6:222–30. 10.1016/j.ebiom.2016.02.025
4. Giovannetti A, Pierdominici M, Mazzetta F, Marziali M, Renzi C, Mileo AM, et al. Unravelling the complexity of T cell abnormalities in common variable immunodeficiency. J Immunol. 2007; 178(6):3932–43. 10.4049/jimmunol.178.6.3932
5. Turpin D, Furudoi A, Parrens M, Blanco P, Viallard JF, Duluc D. Increase of follicular helper T cells skewed toward a Th1 profile in CVID patients with non-infectious clinical complications. Clin Immunol. 2018; 197:130–8. 10.1016/j.clim.2018.09.006
6. Unger S, Seidl M, van Schouwenburg P, Rakhmanov M, Bulashevska A, Frede N, et al. The TH1 phenotype of follicular helper T cells indicates an IFN-γ–associated immune dysregulation in patients with CD21low common variable immunodeficiency. J Allergy Clin Immunol. 2018; 141(2):730–40. 10.1016/j.jaci.2017.04.041
7. Bonilla FA, Barlan I, Chapel H, Costa-Carvalho BT, Cunningham-Rundles C, de la Morena MT, et al. International Consensus Document (ICON): common variable immunodeficiency disorders. J Allergy and Clin Immunol Pract. 2016; 4(1):38–59. 10.1016/j.jaip.2015.07.025
8. Mokhtari M, Shakeri A, Mirminachi B, Abolhassani H, Yazdani R, Grimbacher B, Aghamohammadi A. Important Factors Influencing Severity of Common Variable Immunodeficiency. Arch Iran Med. 2016;19(8):544–50.
9. Warnatz K, Denz A, Dräger R, Braun M, Groth C, Wolff-Vorbeck G, et al. Severe deficiency of switched memory B cells (CD27+IgM-IgD-) in subgroups of patients with common variable immunodeficiency: A new approach to classify a heterogeneous disease. Blood. 2002; 99(5):1544–51. 10.1182/blood.v99.5.1544
10. Wehr C, Kivioja T, Schmitt C, Ferry B, Witte T, Eren E, et al. The EUROclass trial: Defining subgroups in common variable immunodeficiency. Blood. 2008; 111(1):77–85. 10.1182/blood-2007-06-091744
11. Maffucci P, Filion CA, Boisson B, Itan Y, Shang L, Casanova JL, et al. Genetic diagnosis using whole exomesequencing in common variable immunodeficiency. Front Immunol. 2016; 7(Jun):1–9. 10.3389/fimmu.2016.00220
12. McHeyzer-Williams M, Okitsu S, Wang N, McHeyzer-Williams L. Molecular programming of B cell memory. Nat Rev Immunol. 2012; 12(1):24–34. 10.1038/nri3128
13. Ettinger R, Sims GP, Fairhurst AM, Robbins R, da Silva YS, Spolski R, et al. IL-21 Induces differentiation of human naive and memory b cells into antibody-secreting plasma cells. J Immunol. 2005; 175(12):7867–79. 10.4049/jimmunol.175.12.7867
14. Lanzavecchia A, Sallusto F. Human B cell memory. Curr Opin Immunol. 2009; 21(3):298–304. 10.1016/j.coi.2009.05.019
15. Fagarasan S, Honjo T. T-independent immune response: new aspects of B cell biology. Science. 2000; 290(5489):89–92. 10.1016/j.isci.2022.105002
16. Capolunghi F, Cascioli S, Giorda E, Rosado MM, Plebani A, Auriti C, et al. CpG drives human transitional B cells to terminal differentiation and production of natural antibodies. J Immunol. 2008;180(2):800–8. 10.4049/jimmunol.180.2.800
17. Ameratunga R, Allan C, Woon ST. Defining common variable immunodeficiency disorders in 2020 (Vol. 40). In Immunology and Allergy Clinics of North America. W.B. Saunders; 2020. p. 403–20.
18. Cunningham-Rundles C. Common variable immune deficiency: case studies. Hematology. 2019; 2019(1):449–56. 10.1182/hematology.2019002062
19. López-Herrera G, Segura-Méndez N, O’Farril-Romanillos P, Nuñez-Nuñez M, Zarate-Hernández M, Mogica-Martínez D, et al. Low percentages of regulatory T cells in common variable immunodeficiency ( CVID ) patients with autoimmune diseases and its association with increased numbers of CD4 + CD45RO + T and CD21 low B cells. Allergol Immunopathol. 2019; 47(5):457–66. 10.1016/j.aller.2019.01.003
20. Azizi G, Abolhassani H, Kiaee F, Tavakolinia N, Rafiemanesh H, Yazdani R, et al. Autoimmunity and its association with regulatory T cells and B cell subsets in patients with common variable immunodeficiency. Allergol Immunopathol. 2018; 46(2):127–35. 10.1016/j.aller.2017.04.004
21. Grześk E, Dąbrowska A, Urbańczyk A, Ewertowska M, Wysocki M, Kołtan S. Common variable immunodeficiency: different faces of the same disease. Postepy Dermatol Alergol. 2021; 38(5):873–80. 10.5114/ada.2021.110067
22. Warnatz K, Schlesier M. Flowcytometric phenotyping of common variable immunodeficiency. Cytometry B Clin Cytom. 2008; 74(5):261–71. 10.1002/cyto.b.20432
23. Fekrvand S, Khanmohammadi S, Abolhassani H, Yazdani R. B-and T-Cell Subset Abnormalities in Monogenic common variable immunodeficiency. Front Immunol. 2022; 13(June):1–16. 10.3389/fimmu.2022.912826
24. Azizi G, Rezaei N, Kiaee F, Tavakolinia N, Yazdani R, Mirshafiey A, et al. T-cell abnormalities in common variable immunodeficiency. J Investig Allergol Clin Immunol. 2016; 26(4):233–43. 10.18176/jiaci.0069
25. Więsik-Szewczyk E, Rutkowska E, Kwiecień I, Korzeniowska M, Sołdacki D, Jahnz-Różyk K. Patients with common variable immunodeficiency complicated by autoimmune phenomena have lymphopenia and reduced treg, Th17, and NK cells. J Clin Med. 2021; 10(15). 10.3390/jcm10153356
26. Yesillik S, Gupta S. Phenotypically defined subpopulations of circulating follicular helper T cells in common variable immunodeficiency. Immun Inflamm Dis. 2020; 8(3):441–6. 10.3389/fimmu.2017.00174
27. Ueno H. Human circulating T follicular helper cell subsets in health and disease. J Clin Immunol. 2016; 36:34–9. 10.1080/25785826.2020.1776079
28. Yesillik S, Agrawal S, Gollapudi S V., Gupta S. Phenotypic analysis of CD4+ Treg, CD8+ Treg, and Breg cells in adult common variable immunodeficiency patients. Int Arch Allergy Immunol. 2019; 180(2):150–8. 10.1159/000501457
29. Good KL, Bryant VL, Tangye SG. Kinetics of human B cell behavior and amplification of proliferative responses following stimulation with IL-21. J Immunol. 2006; 177(8):5236–47. 10.4049/jimmunol.177.8.5236
30. Clemente A, Pons J, Matamoros N, Iglesias J, Ferrer JM. B cells from common variable immunodeficiency patients fail to differentiate to antibody secreting cells in response to TLR9 ligand (CpG-ODN) or anti-CD40+IL21. Cell Immunol. 2011; 268(1):9–15. 10.1016/j.cellimm.2011.01.004
31. Cunningham-Rundles C, Radigan L, Knight AK, Zhang L, Bauer L, Nakazawa A. TLR9 activation is defective in common variable immune deficiency. J Immunol. 2006; 176(3):1978–87. 10.4049/jimmunol.176.3.1978