
Download
ORIGINAL ARTICLE
Simvastatin alleviates experimental autoimmune encephalomyelitis through regulating the balance of Th17 and Treg in mice
Dongsheng Xu, Manxia Wang*, Lijuan Wang
Department of Neurology, The Second Hospital of Lanzhou University, Lanzhou, China
Abstract
The aim of this study was to elucidate the therapeutic effect of simvastatin on experimental autoimmune encephalomyelitis (EAE) by regulating the balance between Th17 and Treg cells in mice. C57BL/6 mice were randomly divided into four groups: normal group, EAE group, simvastatin (2 and 10 mg/kg) group, and AG490 group (with AG490 serving as the positive control). Neurological function scores of mice were assessed daily. The four groups received treatments of normal saline, normal saline, and simvastatin (2 and 10 mg/kg), respectively. In the AG490 group, mice were injected intraperitoneally with AG490 (1 mg) every other day, and treatment was halted after 3 weeks. The spinal cord was stained with hematoxylin and eosin (H&E), and immunohistochemical staining for retinoic acid receptor-related orphan receptor γ(RORγ) and Foxp3 (Foxp3) was performed. Spleen samples were taken for Th17 and Treg analysis using flow cytometry. The levels of interleukin-17 and transforming growth factor-β (TGF-β) were detected using enzyme-linked immunosorbent assay (ELISA). In the simvastatin and AG490 groups, recovery from neurological impairment was earlier compared to the EAE group, and the symptoms were notably improved. Both simvastatin and AG490 reduced focal inflammation, decreased RORγ-positive cell infiltration, and significantly increased the number of FOXP3-positive cells. The number of Th17 cells and the level of IL-17 in the spleen were decreased in the simvastatin and AG490 treatment groups, while the number of Treg cells and TGF-β levels were significantly increased across all treatment groups. Simvastatin exhibits anti-inflammatory and immunomodulatory effects, potentially alleviating symptoms of neurological dysfunction of EAE. Regulating the balance between Th17 and Treg may represent a therapeutic mechanism for simvastatin in treating EAE.
Key words: Experimental Autoimmune Encephalomyelitis, Multiple Sclerosis, Simvastatin, Th17, Treg
*Corresponding author: Wang Man Xia, Department of Neurology, The Second Hospital of Lanzhou University, Lanzhou 730000, China. Email address: [email protected]
Received 22 March 2024; Accepted 22 May 2024; Available online 1 September 2024
Copyright: Xu D, et al.
License: This open access article is licensed under Creative Commons Attribution 4.0 International (CC BY 4.0). http://creativecommons.org/licenses/by/4.0/
Introduction
Multiple sclerosis (MS) is a common disease of the central nervous system (CNS) characterized by autoimmune demyelination. Experimental autoimmune encephalomyelitis (EAE) is often considered as a mouse model of MS1 and is the most widely used animal model of demyelinating disease.
MS is a neurological disorder of the CNS, affecting young people.2 It is essentially a chronic inflammation with demyelinating, degenerative, and autoimmune properties, and within the CNS.3 Genetic factors and environmental exposures can trigger MS, with increased incidence potentially related to lifestyle changes and increased stress.4,5 Due to the complex pathogenesis of MS, the treatment strategy of “one drug for one target for one disease” does not always succeed.6 Effective treatments are lacking, highlighting the clinical need to develop more efficient drugs with fewer side effects to treat MS. The main therapeutic mechanism of teriflunomide is to disrupt DNA synthesis and subsequently inhibit proliferating T and B lymphocytes,7 which can also cause liver enzyme abnormalities, infection, hair loss, skin allergies, and other side effects.7 Therefore, monitoring for adverse reactions of these drugs is necessary. Although some countries have developed drugs for treating MS, curing MS remains a distant goal.
In immune cells, CD4+ T cells play an important role in the occurrence and development of MS.3 Among these cells, CD4+ T-helper 17 (Th17) cells can produce IL-17, a heterogeneous population, and can be divided into two types, pathogenic and non-pathogenic Th17 cells based on their immune effects.8 Non-pathogenic Th17 cells help maintain tissue homeostasis9, while pathogenic Th17 cells are closely associated with human autoimmune diseases, such as multiple sclerosis, rheumatoid arthritis, and so on.10 Th17 cells are recognized as one of the important immune cells in mouse models of EAE.11 Regulatory T cells (Treg cells) primarily play an inhibitory role, inhibiting the activation of other immunoreactive cells, and crucial in maintaining immune homeostasis and suppressing autoimmune responses in MS/EAE diseases.12,13 Abnormalities in TREGs cell number or function can lead to various autoimmune diseases, including multiple sclerosis.14
Statins, such as simvastatin, lovastatin, have biosynthetic effects on cholesterol. They inhibit hydroxymethylglutaryl Coenzyme A (HMG-CoA) reductase, which catalyzes the conversion of HMG-CoA and reduce to mevalonate, a class of drugs.15 Statins not only have potential cholesterol-lowering effects but also have other therapeutic properties. For example, lovastatin has been shown to treat Alzheimer’s disease,16 MS,17 and other conditions through its immunomodulatory properties.18
In summary, simvastatin has shown promise in treating EAE. potentially through its effects on Th17 and Treg cells. Therefore, the purpose of our study is to determine whether simvastatin has therapeutic effects. The results demonstrates that it can alleviate the neurological symptoms of EAE, improve pathological changes in the nervous system, balance the distribution of Th17 and Treg cells in the CNS, and ultimately provide a valuable therapeutic basis for using simvastatin to treat MS.
Materials and Methods
Materials
The Simvastatin used in this study was provided by Merk Sharp & Dohme.
Experimental animals
The experimental animals were female C57BL/6 mice. They were 8–10 weeks old and weighed 17–20 g. They were purchased from the Animal Experimental Center of Yangzhou University. The mice were placed in a specific, pathogen-free animal barrier environment. The mice were kept in a controlled, 12-h alternating light and dark environment and provided with constant food and water for a week. This study was approved by the Animal Ethics Committee of Lanzhou University Second Hospital (Lanzhou, China).
EAE model preparation and clinical assessment
Briefly,10 mg/ml of MOG35-55 peptide in PBS was first emulsified in a ratio of 1:1. Then, 4 mg/mL of Mycobacterium tuberculosis was added to the complete Freund adjuvant. These emulsions were then injected into four different points on the back of the mice. Later, 250 ng of pertussis toxin (PTX) in PBS was administered on day 1 (hour 1) and day 3 (hour 48). The PBS used (pH = 7.4) was prepared in the laboratory.
On day 1, after monitoring, mice in the simvastatin treatment group was administered normal saline and simvastatin (2 and 10 mg/kg) orally at night. This treatment regimen continued until day 21 post-immunization. The control group received the same amount of normal saline using the same method. All mice were weighed daily on the first day after immunization. In addition, the mice were scored to assess clinical symptoms of EAE. The symptom scoring criteria are as follows: Grade 0 indicated no obvious symptoms; Grade 1 represented complete tail paralysis; Grade 2 indicated mild hind limb paralysis; Grade 3 represented complete paralysis of one hind limb; Grade 4 represented bilateral hind limb paralysis; and Grade 5 indicated complete limb paralysis (quadriplegia), near-death state, or state of death.19
Grouping and treatment
The mice were randomly divided into different treatment groups: normal, EAE, simvastatin (2 and 10 mg/kg), and AG490 groups. The simvastatin treatment groups were treated with simvastatin doses of 2 and 10 mg/kg through gavage, respectively from the first day post EAE induction until the 21st day. The normal and EAE groups followed the same treatment regimen as the simvastatin groups, but only normal saline was given instead. AG490 is a JAK2-specific tyrosine kinase inhibitor, which mainly affects the differentiation and functional maintenance of Th17 cells by inhibiting the JAK2-STAT3 signaling pathway. AG490 served as the positive control. In the AG490 group, mice were intraperitoneally injected with 1 mg of AG490 every other day.
Histological Evaluation
Following 21 days of treatment post-immunization, histological assessment was conducted on mice from each group. Initially, the spinal cords were dissected from all five groups. The specimens were then numbered and fixed. Tissue samples were subsequently embedded with paraffin wax. Specimens were sectioned into 5 μm slices. Histopathological analysis was performed after staining with hematoxylin and eosin (H&E).
Immunohistochemistry
The sections were initially dewaxed and antigen-repaired under high temperature and pressure. Then, the slices were added to an H2O2 solution for 20 min to eliminate the effect of endogenous enzymes. The primary antibody was then added to the specimen. The slices were incubated at 4°C for 15 h. Afterward, they were rinsed with PBS three times.
The primary antibodies used were anti-retinoic acid receptor-related orphan receptor γ (anti-RORγ)(dilution 1:200, Abcam, Cambridge, UK) and anti-forkhead box protein P3(anti-Foxp3) (dilution 1:100, Abcam). The secondary antibody (ZSCB-BIO) was purchased.
Flow cytometry
On day 21 post-immunization, specimens were examined by flow cytometry. The test indexes were the number of CD4+ interleukin-17, CD25+ Foxp3+T cells, Th17 cells, and Treg cells in the mouse spleen. Spleen mononuclear cells were isolated first. Subsequently, PE-labeled anti-mouse CD4 or FITC-labeled anti-mouse CD25 was added to the specimen. PE-labeled anti-IL-17 antibody (Bioscience) were used to stain the Th17 cells in mouse samples. Therefore, mouse anti-CD25 (Biolegend) and mouse anti-FoxP3 (Ebioscience) antibodies were used to stain Treg cells.
ELISA
On the 21st day after immunization, the spleen was collected after the mice were sacrificed. Tissue samples were then analyzed to determine the levels of IL-17 and transforming growth factor-β (TGF-β). An enzyme-linked immunosorbent assay (ELISA) kit (Elabscience, Wuhan, China) was used to detect specimens. The concentrations of IL-17 and TGF-β in the spleen were detected according to the instructions.
Statistical analysis
The SPSS 26.0 statistical software (IBM, USA) was used for statistical analysis of the relevant experimental data. One-way analysis of variance (ANOVA) was used to analyze the differences between groups. All the data are expressed as mean ± standard deviation (SD). P < 0.05 was considered statistically significant.
Results
Effect of simvastatin on the symptoms of neurological deficit in EAE mice
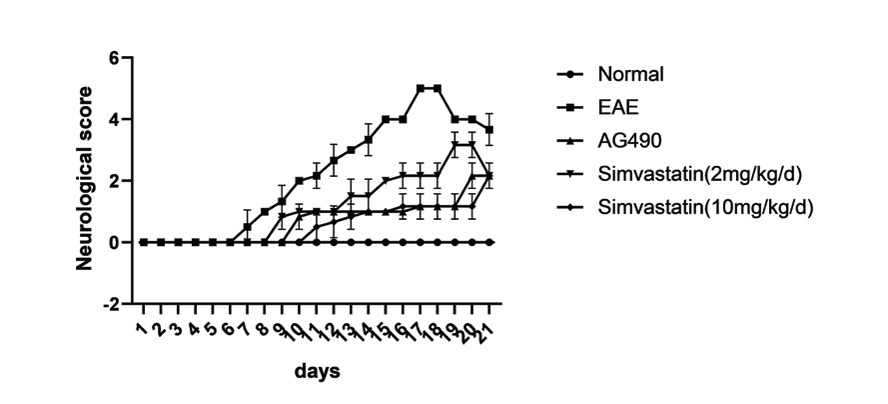
On the 6th day after immunization, EAE mice began to show neurological deficits. The treatment was started on day 1 after immunization. As shown in the figure, symptoms in the EAE, simvastatin (2 mg/kg), AG490, and simvastatin (10 mg/kg) groups peaked on days 17, 19, 20, and 21 (Figure 1).
Figure 1 Simvastatin alleviated the symptoms of neurological deficit in experimental autoimmune encephalomyelitis (EAE) mice. The 5-point EAE score was assessed in mice from Days 1 to 21 post-immunization. All data are expressed as mean ± standard deviation (n=6 in each group).
Effect of simvastatin on pathological changes of spinal cord tissue in EAE mice
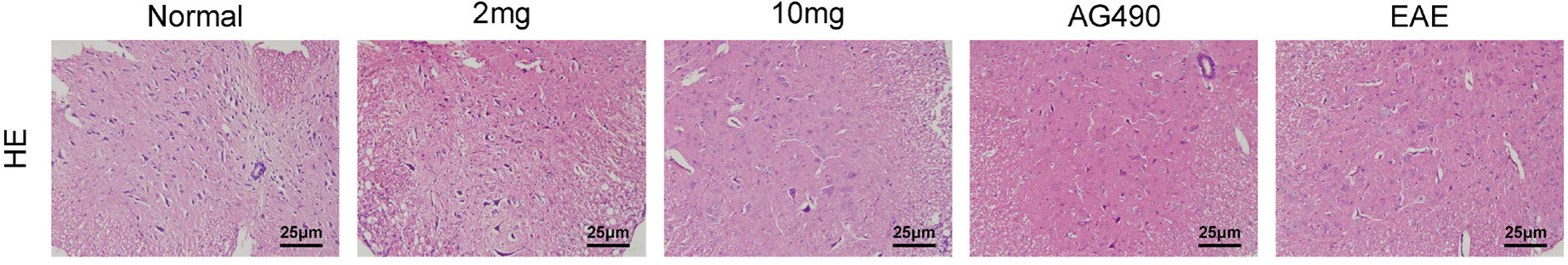
The pathological changes of the spinal cord in the 5 groups were observed 21 days after immunization. HE staining showed no evident inflammation in the control group. A significant number of inflammatory cells were observed in specimens from the EAE group. In addition, the number of inflammatory cells in the specimen was reduced after treatment with simvastatin (2 and 10 mg/kg) and AG490. Mice in the EAE group showed increased demyelinating foci compared to the control group. The demyelinating lesions in the simvastatin- and AG490-treated groups were less than in the EAE group (Figure 2).
Figure 2 Simvastatin reduced inflammation and demyelination in the spinal cord of EAE mice. On the 21st day post-immunization, mice’s spinal cords were separated, sectioned, and stained with hematoxylin-eosin (H&E). Representative sections of H&E.
Effect of simvastatin on the infiltration of Th17 and Treg cells in the spinal cord of EAE
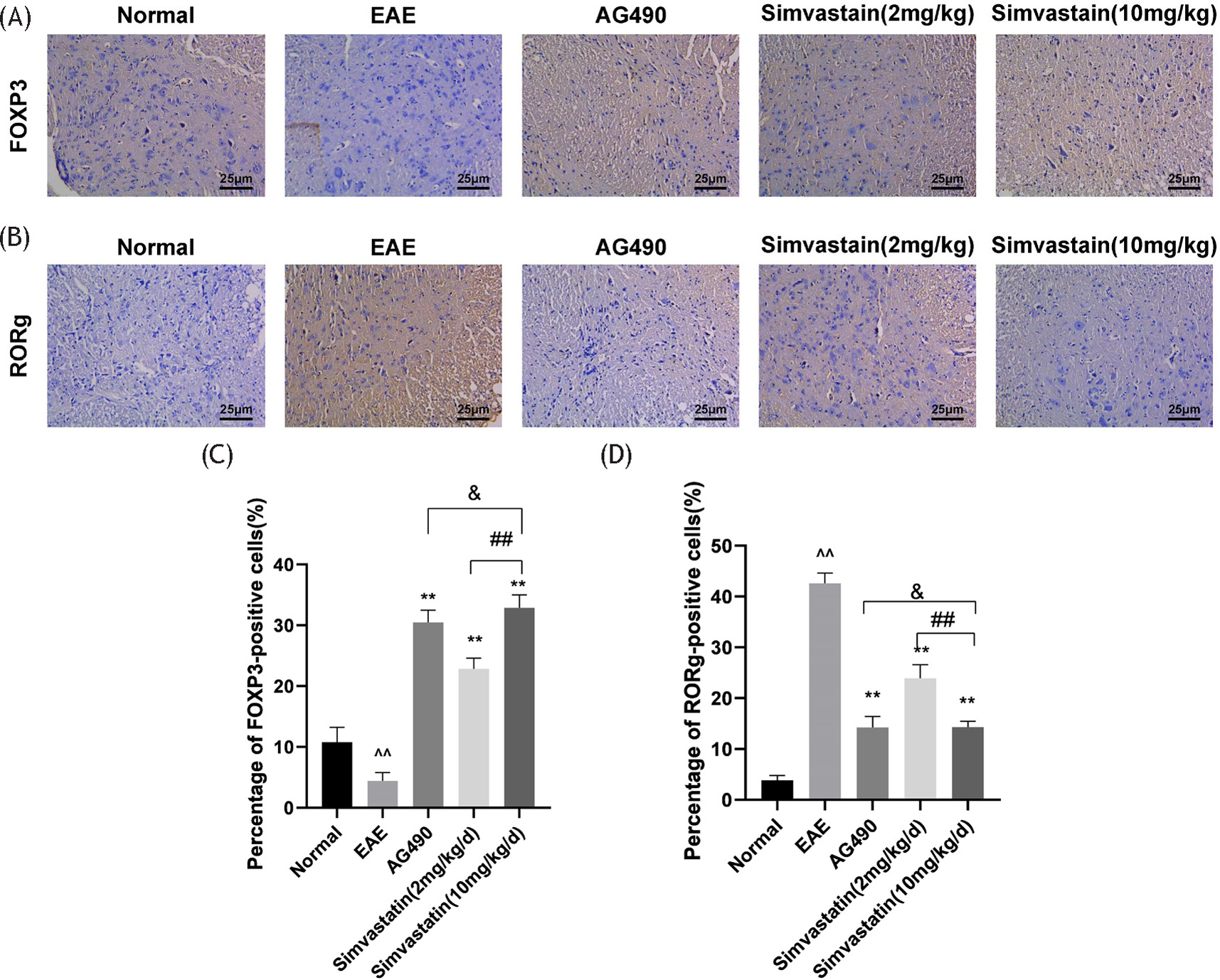
The spinal sections of mice were examined by immunohistochemical staining to detect the effect of simvastatin on Th17 and Treg cell infiltration. Among them, RORγ staining reflected the number of Th17 cells, and Foxp3 staining reflected the number of Treg cells. In the EAE group, the number of Treg cells in the spinal cord of mice was decreased compared to the control group. Treg cell count increased after treatment with simvastatin (2 and 10 mg/kg) or AG490. In addition, the number of Treg cells in the spinal cord of mice treated with simvastatin (2 mg/kg) was lower compared with that in the simvastatin (10 mg/kg) group (Figure 3A and 3C).
Figure 3 Effects of simvastatin on the numbers of T helper 17 (Th17) and regulatory T (Treg) cells in the spinal cord of EAE mice. The mice spinal cord sections were stained by immunohistochemistry. (A) The number of Treg cells is indicated by forkhead box protein P3 (Foxp3) staining, (B) while the number of Th17 cells is reflected by retinoic acid receptor-related orphan receptor γ (RORγ) staining, (C) Treg cell number, (D) Th17 cell number. All data are expressed as mean ± standard deviation (n=5 in each group). **P<0.01, EAE vs. AG490 or Simvastatin (2 mg/kg) or Simvastatin (10 mg/kg), *P<0.05, EAE vs. AG490 or Simvastatin (2 mg/kg) or Simvastatin(10 mg/kg); ^^P<0.01, EAE vs. normal,^P<0.05, EAE vs. normal. ##P<0.01, Simvastati n(10 mg/kg) vs. Simvastatin (2 mg/kg), #P<0.05, Simvastatin (10 mg/kg) vs. simvastatin (2 mg/kg). & no significant difference, Simvastatin(10 mg/kg) vs. AG490.
The infiltration of Th17 cells in the EAE group was increased compared with the control group. Treatment with simvastatin (2 and 10 mg/kg) or AG490 can reduce the number of Th17 cells infiltrating the spinal cord. In addition, Th17 cell infiltration was increased in the simvastatin (2 mg/kg) group compared with the simvastatin (10 mg/kg) group (Figure 3B and 3D).
Effects of simvastatin on the numbers of Th17 and Treg cells in EAE mice spleen and levels of IL-17 and TGF-β in the spleen
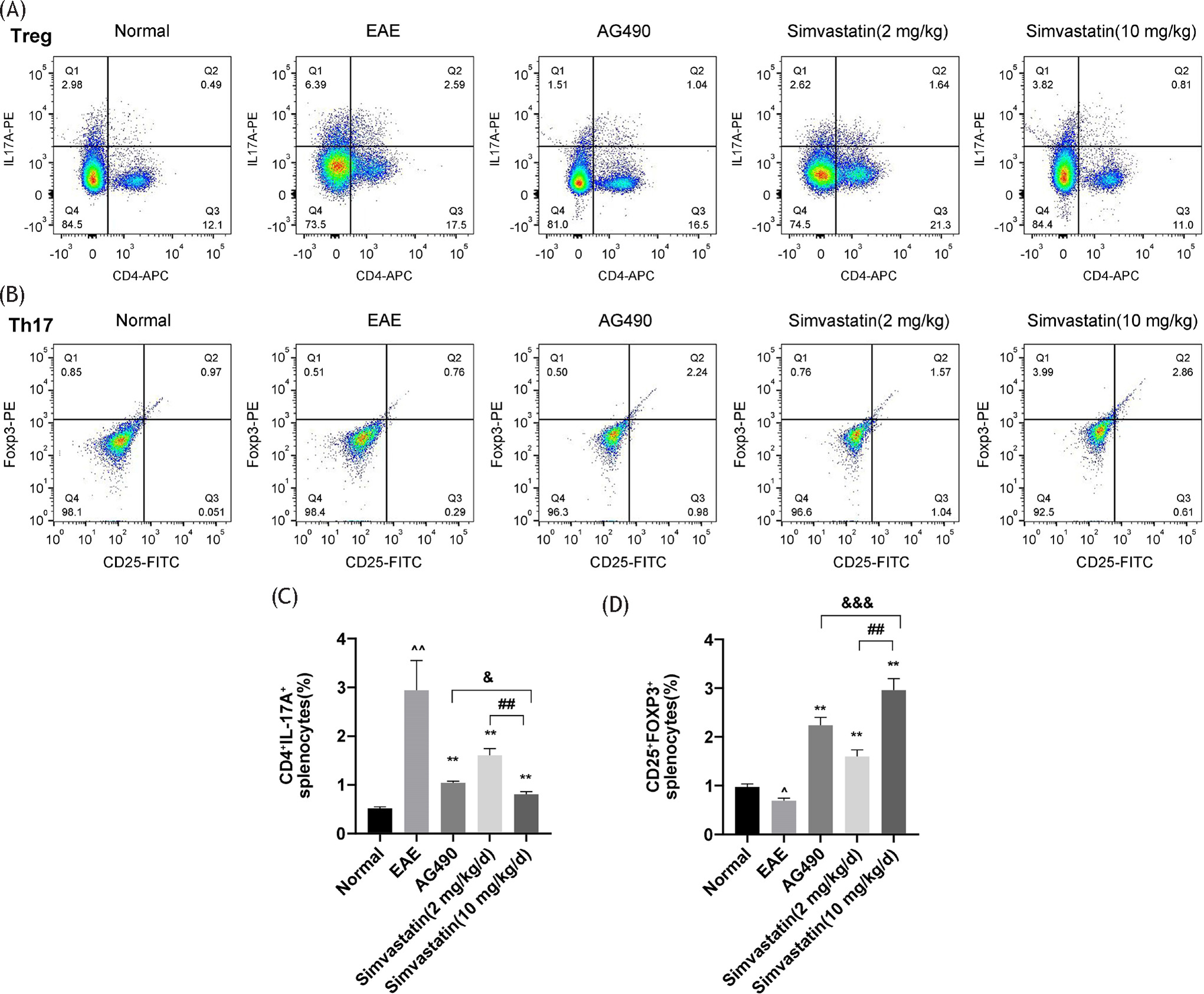
Flow cytometry revealed a higher count of Th17 cells in the EAE group compared to the control group. The number of Th17 cells in the simvastatin (2 and 10 mg/kg) and AG490 treatment groups was lower than in the EAE group. The number of Th17 cells in the simvastatin (2 mg/kg) group was higher than that in the simvastatin (10 mg/kg) group. The number of Th17 cells in the simvastatin (10 mg/kg) group was higher than that in the AG490 group, while the number of Th17 cells in the simvastatin (10 mg/kg) group was higher than that in the AG490 group. In addition, the number of Treg cells in the EAE group was lower than that in the control group, while the number of Treg cells in the simvastatin (2 and 10 mg/kg) or AG490 treatment groups was higher than that in the EAE group, and the number of Treg cells in the simvastatin (2 mg/kg) group was lower than that in the simvastatin (10 mg/kg) group (Figure 4A–D).
Figure 4 Effects of simvastatin on the numbers of T helper 17 (Th17) and regulatory T (Treg) cells in EAE mice spleen (A,B) The numbers of Th17 cells (CD4+ IL-17A) and Treg cells (CD25+ Foxp3) in mice spleen were determined by flow cytometry. (C,D) Statistical analyses of the numbers of Th17 cells and Treg cells. The data are expressed as mean ± standard deviation (n=4 in each group). **P<0.01, EAE vs. AG490 or Simvastatin (2 mg/kg) or Simvastatin (10 mg/kg), *P<0.05, EAE vs. AG490 or Simvastatin (2 mg/kg) or Simvastatin (10 mg/kg); ^^P<0.01, EAE vs. normal, ^P<0.05, EAE vs. normal. ##P<0.01, Simvastatin (10 mg/kg) vs. Simvastatin (2 mg/kg), #P<0.05, Simvastatin (10 mg/kg) vs. Simvastatin (2 mg/kg). & no significant difference, Simvastatin (10 mg/kg) vs. AG490, &&P<0.05, Simvastatin (10 mg/kg) vs. AG490, &&&P<0.01, Simvastatin (10 mg/kg) vs. AG490.
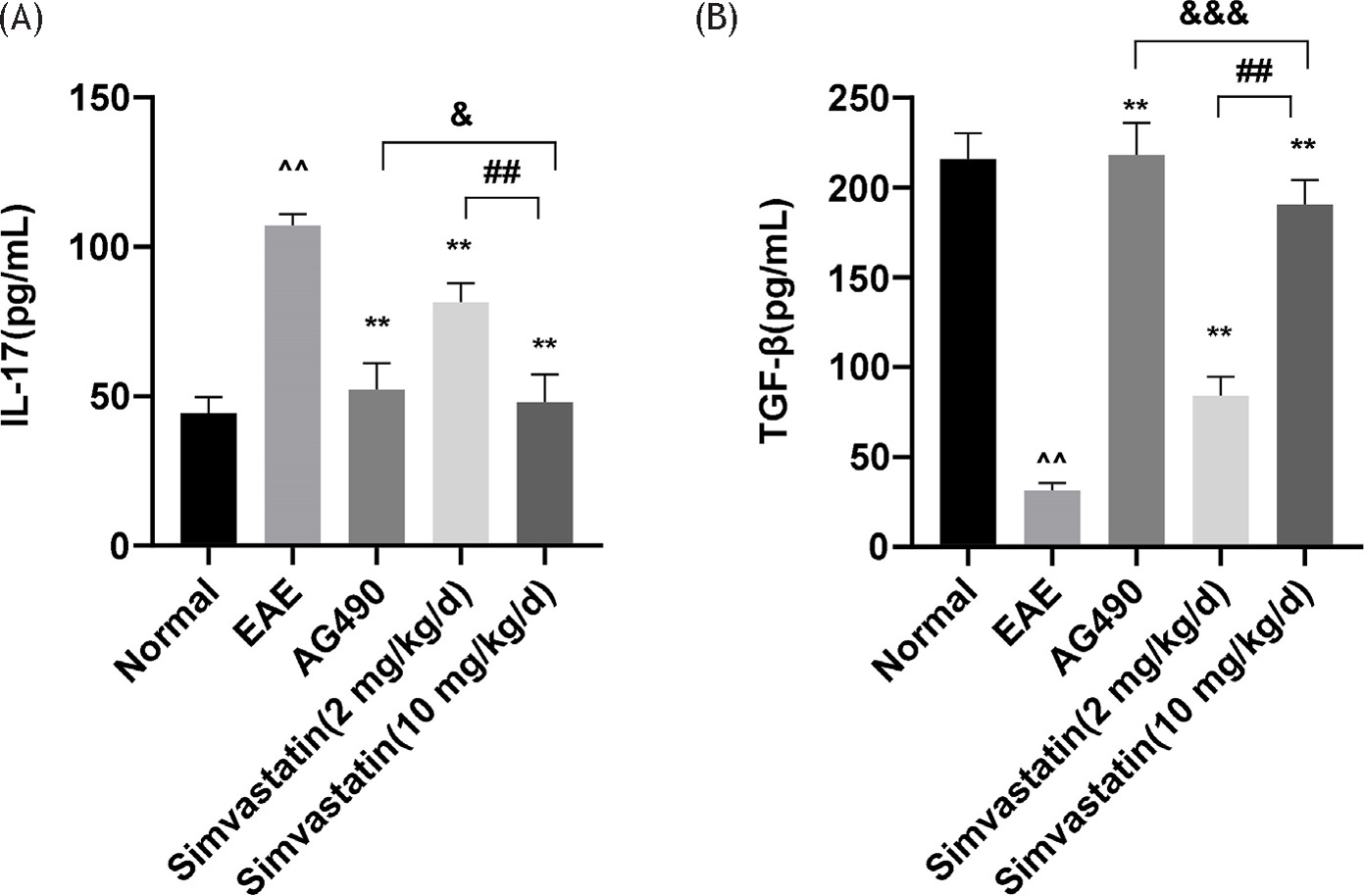
Meanwhile, ELISA was used to measure the levels of IL-17 and TGF-β in the spleens of mice. The results showed that the EAE group exhibited increased IL-17 levels and decreased TGF-β levels in the spleen. It was also found that simvastatin (2 and 10 mg/kg) or AG490 groups reduced the expression level of IL-17 and increased the level of TGF-β. In addition, IL-17 levels were increased and TGF-β levels were decreased in the simvastatin (2 mg/kg) group compared with the simvastatin (10 mg/kg) group. Compared with the AG490 group, the simvastatin (10 mg/kg) group showed decreased TGF-β levels in the spleens of mice (Figure 5A and 5B).
Figure 5 Effects of simvastatin on levels of interleukin (IL)-17 and TGF-β in EAE mice spleen. (A,B)The levels of IL-17 and TGF-β in the spleen were determined by enzyme-linked immunosorbent assay (ELISA). The data are expressed as mean ± standard deviation (n=5 in each group). **P<0.01, EAE vs. AG490 or Simvastatin (2 mg/kg) or Simvastatin (10 mg/kg), *P<0.05, EAE vs. AG490 or Simvastatin (2 mg/kg) or Simvastatin (10 mg/kg); ^^P<0.01, EAE vs. normal, ^P<0.05, EAE vs. normal. ##P<0.01, Simvastatin (10 mg/kg) vs. Simvastatin (2 mg/kg), #P<0.05. Simvastatin (10 mg/kg) vs. Simvastatin (2 mg/kg). & no significant difference, Simvastatin (10 mg/kg) vs. AG490, &&P<0.05, Simvastatin (10 mg/kg)vs. AG490, &&&P<0.01,Simvastatin (10 mg/kg) vs. AG490.
Discussion
Some studies have indicated that certain types of drugs, such as antilipidemic and chemotherapeutic drugs, have anti-inflammatory properties.20 Therefore, these drugs may reveal novel therapeutic effects for various diseases, which could hold significant potential for treating certain diseases, such as autoimmune diseases and so on. It could potentially provide substantial therapeutic value and benefits to many patients.
Statins were previously used to treat hyperlipidemia. However, in recent years, they have been described to negatively regulate inflammation in various models, such as autoimmune neuritis and inflammatory bowel disease.21 It was found that simvastatin treatment can effectively prevent disease progression in patients with RRMS and reduce central nervous system lesions.22 However, the pharmacological mechanisms remain unclear.
Th17 cells play an important role in the occurrence and development of EAE. When Th17 cells are impaired in function, the Th17 characteristic gene is reduced, reducing CNS inflammation.23 Treg cells can also limit the occurrence and development of EAE.24 Interestingly, simvastatin and atorvastatin therapy can increase Treg and decrease Th17 cell count in animal inflammatory disease models.21
In this study, we observed the following: (i) simvastatin treatment reduced the number of inflammatory cells and effectively improved neural function scores. (ii) It reduced the number of Th17 cell infiltrating lesions and increased the number of Treg cells. (iii) After simvastatin treatment, the relevant specimens were found to reduce the number of Th17 cells and the levels of IL-17 in the spleen of EAE mice and increase the number of Treg cells and TGF-β levels.
To evaluate the therapeutic effect of simvastatin on MS, we treated EAE mice with simvastatin; using AG490 as the positive control and normal saline as the negative control. Our findings demonstrated that simvastatin treatment effectively improved the neurological deficits in EAE mice. Images showed that mice in the EAE group treated with normal saline experienced spontaneous symptom improvement after the disease peaked. However, improvements in symptoms of mice treated with simvastatin (low and high dose groups) or AG490 occurred earlier and were more pronounced than in the EAE group. The starting point of treatment in all studies was EAE induction day. Treatment initiation coincided with the induction of EAE, highlighting the positive therapeutic efficacy of simvastatin administered at the onset of EAE symptoms.
Treg cells act as regulatory cells that suppress CNS autoimmune response in EAE by suppressing Th17-mediated responses. TGF-β, a Treg cell-related cytokine, plays a key role in the inhibition and recovery of EAE.25
There is evidence that high levels of TGF-β contribute to EAE recovery and remission.26 Increasing TGF-β can reduce the severity of EAE disease.27 However, Treg and TGF-β-producing autoantigen-reactive Th2 cells were able to transfer reduced and/or reverse EAE, suggesting that Treg cells may play a key role in downregulating the pathogenic T cell response in the pathogenesis of EAE.28
Properly balancing the pro-inflammatory Th17 cell population and immunosuppressive Treg cells is important in maintaining immune homeostasis.29 Imbalances have been linked to inflammatory responses and autoimmune diseases, including MS. This is linked to increased levels of Th17 cells in the body.30
At the same time, inhibition of peripheral T cells with certain drugs has been identified as another valuable treatment for the initial stages of EAE.31,32 This suggests that defects in Treg cells may contribute to the onset of MS. Lower Treg inhibition may promote increased production of pro-inflammatory cytokines, such as IL-17.33 The loss of balance between Th17 and Treg cells plays a crucial role in the onset and development of MS.34 Th17 and Treg cells exhibit differentiation plasticity, being able to transform into each other under different inflammatory conditions in autoimmune diseases 34,35 The Th17/Treg ratio reflects the interaction between these two T-cell subpopulations, highly correlating with disease severity in EAE and MS.36 The ability to regulate the balance between Th17 and Treg is considered a new drug target for treating MS.37
Th17 cells exert pro-inflammatory effects, primarily involved in autoimmune responses and tissue damage, whereas Treg cells possess anti-inflammatory properties and primarily safeguard the central nervous system.38 Therefore, adjusting the Th17/Treg balance is critical for treating EAE. Several studies indicate that the migration of Th17 cells to the central nervous system is a mechanisms of EAE in mice.39 In contrast, Treg cells produce TGF-β, a key regulator of EAE. Inhibiting Th17 cells and upregulating Treg cells are effective strategies to alleviate EAE severity.40
Consequently, we further investigated simvastatin’s regulatory effects on Th17 and Treg cells. Experimental results demonstrated that simvastatin reduces the number of Th17 cells infiltrating lesions, while increasing Treg counts. Simvastatin decreases Th17 cells and IL-17 levels in the spleen of EAE mice, and increase the number of Treg cells and TGF-β levels in the spleen. This suggests that targeting the Th17/Treg ratio is an effective mechanism of simvastatin therapy. Further experiments will be necessary to confirm histologically and verify the roles of Th17 and Treg, cells.
Moreover, we observed that simvastatin decreased Th17 cell counts in both lesions and spleens. This implies that simvastatin may affect the differentiation of immature CD4+ T cells into Th17 cells. TGF-β, a pivotal cytokine, controls this differentiation and maintains stability.41 The protective effect in the brain appears related to reduced brain damage caused by decreased T cell levels. Due to time and funding constraints, further experiments to validate whether T cell level changes contribute to brain protection could not be conducted.
Finally, cytokines related to Th17 and Treg cell differentiation in the spleens of EAE mice were detected. Simvastatin reduced IL-17 levels, suggesting it inhibits native CD4+T cells from differentiating into Th17 cells by reducing IL-17 levels, Thereby reducing Th17 cells infiltrating in EAE lesions. Simvastatin also increased TGF-β levels. Combined with the results from immunohistochemistry, flow cytometry staining, and ELISA, the high-dose group exhibited significantly better therapeutic effects than the low-dose group. This suggests that higher doses of simvastatin are more effective. Given the complexity of the inflammatory response, simvastatin appears to have effects similar to AG490 in this experiment. Since AG490 inhibits the JAK2-STAT3 pathway, further experiments are needed to confirm whether simvastatin has a similar effect.
Conclusions
In short, simvastatin may alleviate EAE symptoms through its anti-inflammatory and immunosuppressive effects. Maintaining a balance between Th17 and Treg cells appears to be its primary mechanism. Simvastatin treats EAE by adjusting the Th17/Treg ratio, and higher doses of simvastatin are more effective. Therefore, simvastatin holds potential value in MS treatment, although further research is needed to understand its specific actions on Th17 and Treg cells in EAE.
Acknowledgments
This work was supported by the 2019 Lanzhou Chengguan District Talent Innovation and Entrepreneurship Project. (Grant No. 2019RCCX0068).
Author contributions
DX and MW carefully designed the study. DX established a mouse model. DX conducted experimental research and wrote and edited the paper. DX and LW collected and analyzed the data. All authors have read and approved the final manuscript and are responsible for the integrity and security of the data. Compliance with ethics guidelines
Conflict of Interest
The authors declare no conflict of interest.
REFERENCES
1. Filippi M, Bar-Or A, Piehl F, et al. Multiple sclerosis. Nat Rev Dis Primers. 2018;4(1):43. 10.1038/s41572-018-0041-4
2. Hosseini A, Masjedi A, Baradaran B, et al. Dimethyl fumarate: regulatory effects on the immune system in the treatment of multiple sclerosis. J Cell Physiol. 2019;234(7):9943-9955. 10.1002/jcp.27930
3. Wunsch M, Jabari S, Voussen B, et al. The enteric nervous system is a potential autoimmune target in multiple sclerosis. Acta Neuropathol. 2017;134(2):281-295. 10.1007/s00401-017-1742-6
4. Hosseinzadeh A, Baneshi MR, Sedighi B, et al. Incidence of multiple sclerosis in Iran: a nationwide, population-based study. Public Health. 2019;175:138-144. 10.1016/j.puhe.2019.07.013
5. Spear ET, Holt EA, Joyce EJ, et al. Altered gastrointestinal motility involving autoantibodies in the experimental autoimmune encephalomyelitis model of multiple sclerosis. Neurogastroenterol Motil. 2018;30(9):e13349. 10.1111/nmo.13349
6. Baecher-Allan C, Kaskow BJ, Weiner HL. Multiple sclerosis: mechanisms and immunotherapy. Neuron. 2018;97(4):742-768. 10.1016/j.neuron.2018.01.021
7. TaSKapilioGLu ÖZ. Recent advances in the treatment for multiple sclerosis; current new drugs specific for multiple sclerosis. Noro Psikiyatr Ars. 2018;55(Suppl1):S15-S20. 10.29399/npa.23402
8. Stockinger B, Omenetti S. The dichotomous nature of T Helper 17 cells. Nat Rev Immunol. 2017;17(9):535-544. 10.1038/nri.2017.50
9. Omenetti S, Bussi C, Metidji A, et al. The intestine harbors functionally distinct homeostatic tissue-resident and inflammatory Th17 cells. Immunity. 2019;51(1):77-89.e6. 10.1016/j.immuni.2019.05.004
10. Yasuda K, Takeuchi Y, Hirota K. The pathogenicity of Th17 cells in autoimmune diseases. Semin Immunopathol. 2019;41(3):283-297. 10.1007/s00281-019-00733-8
11. Balasa R, Barcutean L, Balasa A, et al. The action of TH17 cells on blood-brain barrier in multiple sclerosis and experimental autoimmune encephalomyelitis. Hum Immunol. 2020;81(5):237-243. 10.1016/j.humimm.2020.02.009
12. Kaskow BJ, Baecher-Allan C. Effector T cells in multiple sclerosis. Cold Spring Harb Perspect Med. 2018;8(4):a029025. 10.1101/cshperspect.a029025
13. Stadhouders R, Lubberts E, Hendriks RW. A cellular and molecular view of T helper 17 cell plasticity in autoimmunity. J Autoimmun. 2018;87:1-15. 10.1016/j.jaut.2017.12.007
14. Yang SJ, Xie CC, Chen Y, et al. Differential roles of TNFalpha-TNFR1 and TNFalpha-TNFR2 in the differentiation and function of CD4(+)Foxp3(+) induced Treg cells in vitro and in vivo periphery in autoimmune diseases. Cell Death Dis. 2019;10(1):27. 10.1038/s41419-018-1266-6
15. Rodrigues KA, Rocha RT, Mulinari FF, et al. Exploring the Brazilian diversity of aspergillus sp. strains for lovastatin and itaconic acid production. Fungal Genet Biol. 2020;138:103367. 10.1016/j.fgb.2020.103367
16. Loera-Valencia R, Goikolea J, Parrado-Fernandez C, et al. Alterations in cholesterol metabolism as a risk factor for developing Alzheimer’s disease: potential novel targets for treatment. J Steroid Biochem Mol Biol. 2019;190:104-114. 10.1016/j.jsbmb.2019.03.003
17. Hasan H, Rahim MHA, Campbell L, et al. Overexpression of acetyl-CoA carboxylase in Aspergillus terreus to increase lovastatin production. N Biotechnol. 2018;44:64-71. 10.1016/j.nbt.2018.04.008
18. Yoon LS, Goodman MT, Rimel BJ, et al. Statin use and survival in elderly patients with endometrial cancer. Gynecol Oncol. 2015;137(2):252-257. 10.1016/j.ygyno.2015.01.549
19. Schneider C, Schuetz G, Zollner TM. Acute neuroinflammation in lewis rats—a model for acute multiple sclerosis relapses. J Neuroimmunol. 2009;213(1-2):84-90. 10.1016/j.jneuroim.2009.05.015
20. Robichon K, Patel V, Connor B, et al. Clozapine reduces infiltration into the CNS by targeting migration in experimental autoimmune encephalomyelitis. J Neuroinflammation. 2020;17(1):53. 10.1016/j.jneuroim.2009.05.015
21. Côté-Daigneault J, Mehandru S, Ungaro R, et al. Potential immunomodulatory effects of statins in in flammatory bowel disease. Inflamm Bowel Dis. 2016;22(3):724-732. 10.1097/MIB.0000000000000640
22. Vollmer T, Key L, Durkalski V, et al. Oral simvastatin treatment in relapsing-remitting multiple sclerosis. Lancet. 2004;363(9421):1607-1608. 10.1016/S0140-6736(04)16205-3
23. Haines CJ, Chen Y, Blumenschein WM, et al. Autoimmune memory T helper 17 cell function and expansion are dependent on interleukin-23. Cell Rep. 2013;3(5):1378-1388. 10.1016/j.celrep.2013.03.035
24. Kohm AP, Carpentier PA, Anger HA, et al. Cutting edge: CD4+CD25+ regulatory T cells suppress antigen-specific autoreactive immune responses and central nervous system inflammation during active experimental autoimmune encephalomyelitis. J Immunol. 2002;169(9):4712-4716. 10.4049/jimmunol.169.9.4712
25. Haghmorad D, Mahmoudi MB, Salehipour Z, et al. Hesperidin ameliorates immunological outcome and reduces neuroinflammation in the mouse model of multiple sclerosis. J Neuroimmunol. 2017;302:23-33. 10.1016/j.jneuroim.2016.11.009
26. Bettelli E, Das MP, Howard ED, et al. IL-10 is critical in regulating autoimmune encephalomyelitis, as demonstrated by studies of IL-10-and IL-4-deficient and transgenic mice. J Immunol. 1998;161(7):3299–3306. 10.4049/jimmunol.161.7.3299
27. Speck S, Lim J, Shelake S, et al. TGF-beta signaling initiated in dendritic cells instructs suppressive effects on Th17 differentiation at the site of neuroinflammation. PLoS One. 2014;9(7):e102390. 10.1371/journal.pone.0102390
28. Zhang XM, Koldzic DN, Izikson L, et al. IL-10 is involved in the suppression of experimental autoimmune encephalomyelitis by CD25+ CD4+ regulatory T cells. Int Immunol. 2004;16(2):249-256. 10.1093/intimm/dxh029
29. Wu DW, Wang JT, Zhang M, et al. Altered Th17/Treg ratio in nasal polyps with distinct cytokine profile: association with patterns of inflammation and mucosal remodeling. Medicine. 2016;95(10):e2998. 10.1097/MD.0000000000002998
30. Venkatesha SH, Dudics S, Weingartner E, et al. Altered Th17/Treg balance and dysregulated IL-1b response influence susceptibility/resistance to experimental autoimmune arthritis. Int J Immunopathol Pharmacol. 2015;28(3):318-328. 10.1177/0394632015595757
31. Ghadiri M, Rezk A, Li R, et al. Dimethyl fumarate-induced lymphopenia in MS due to differential T-cell subset apoptosis. Neurol Neuroimmunol Neuroinflamm. 2017;4(3):e340. 10.1212/NXI.0000000000000340
32. Dargahi N, Katsara M, Tselios T, et al. Multiple sclerosis: immunopathology and treatment update. Brain Sci. 2017; 7(7):78. 10.3390/brainsci7070078
33. Danikowski KM, Jayaraman S, Prabhakar BS. Regulatory T cells in multiple sclerosis and myasthenia gravis. J Neuroinflammation. 2017;14(1):117. 10.1186/s12974-017-0892-8
34. Venkatesha H, Dudics S, Weingartner E, et al. Altered Th17/Treg balance and dysregulated IL-1β response influence susceptibility/resistance to experimental autoimmune arthritis. Int J Immunopathol Pharmacol. 2015;28(3):318-328. 10.1177/0394632015595757
35. Lu PX, Cao YP, M Wang MH, et al. Mature dendritic cells cause Th17/Treg imbalance by secreting TGF-β1 and IL-6 in the pathogenesis of experimental autoimmune encephalomyelitis. Cent Eur J Immunol. 2016;41(2):143-152. 10.5114/ceji.2016.60987
36. Segal BM. The diversity of encephalitogenic CD4+ t cells in multiple sclerosis and its animal models. J Clin Med. 2019;8(1):120. 10.3390/jcm8010120
37. Qu XB, Han JJ, Zhang Y, et al. MiR-384 regulates the Th17/Treg ratio during experimental autoimmune encephalomyelitis pathogenesis. Front Cell Neurosci. 2017;11:88. 10.3389/fncel.2017.00088
38. Lee PW, Severin ME, Lovett-Racke AE. TGF-β regulation of encephalitogenic and regulatory T cells in multiple sclerosis. Eur J Immunol. 2017;47(3):446-453. 10.1002/eji.201646716
39. Zhao Q, Cheng WJ, I Xi YB, et al. FN-beta regulates Th17 differentiation partly through the inhibition of osteopontin in experimental autoimmune encephalomyelitis. Mol Immunol. 2018;93:20-30. 10.1016/j.molimm.2017.11.002
40. Teixeira NB, Sant’Anna MB, Giardini AC, et al. Crotoxin down-modulates proinflammatory cells and alleviates pain on the MOG35-55-induced experimental autoimmune encephalomyelitis, an animal model of multiple sclerosis. Brain Behav Immun. 2020;84:253-268. 10.1016/j.bbi.2019.12.009
41. Cerboni S, Gehrmann U, Preite S, et al. Cytokine-regulated Th17 plasticity in human health and diseases. Immunology. 2021;163(1):3-18. 10.1111/imm.13280