
Download
ORIGINAL ARTICLE
Auraptene alleviates inflammatory injury and cell apoptosis in children with pneumonia in vitro
Yuebin Wang, Shuzhen Yuan, Wei Tan, Yuanyu Zhou, Ruiyun Liao, Wei Su*
Department of Pediatrics, Affiliated Hospital of Xiangnan University, Chenzhou, Hunan Province, China
*Corresponding author: Wei Su, Department of Pediatrics, Affiliated Hospital of Xiangnan University, No. 31 Renmin West Road, Beihu District, Chenzhou, Hunan Province 423000, China. Email address: [email protected].
Abstract
Objective: The aim of the present study is to investigate the effects of auraptene on inflammation and apoptosis of pneumonia cell model and uncover the mechanism.
Methods: WI-38 cells were treated with lipopolysaccharide (LPS) to construct a pneumonia model. Cell counting kit-8 assay, enzyme-linked-immunosorbent serologic assay, and quantitative polymerase chain reaction assay were conducted to confirm the effects of auraptene on the viability and inflammation of WI-38 cells. Flow cytometry (FCM) and immunoblot assays were conducted to detect the effects of auraptene on the apoptosis of WI-38 cells. Immunoblot assay was performed to confirm the mechanism.
Results: We found that auraptene stimulated cell viability in WI-38 cells upon LPS treatment. Auraptene also inhibited cellular inflammation. Furthermore, auraptene inhibited cell apoptosis of WI-38 cells upon LPS treatment. Mechanically, auraptene inhibited the nuclear factor kappa B signaling pathway, thereby suppressing the pneumonia.
Conclusion: Auraptene alleviates inflammatory injury and cell apoptosis in pneumonia, thus has the potential to act as a pneumonia drug.
Key words: apoptosis, auraptene, inflammation, NF-κB axis, pneumonia
Received 9 August 2023; Accepted 28 August 2023; Available online 1 November 2023
Copyright: Wang Y, et al.
License: This open access article is licensed under Creative Commons Attribution 4.0 International (CC BY 4.0). http://creativecommons.org/licenses/by/4.0/
Introduction
Pneumonia is a global illness with high morbidity and mortality, causing approximately 1.4 million child deaths annually.1 The incidence of pediatric pneumonia is high, with many complications and poor prognosis.2 There is still a lack of effective treatment for pediatric pneumonia, leading to an urgent requirement of the related research and developing an effective regimen3,4 Pneumonia is a serious disease featured by a combination of local and systemic complications. Therefore, it is vital to explore pathogenesis and therapeutic target for treating pneumonia.
Auraptene is isolated from many edible fruits and -vegetables5,6 Auraptene has multiple therapeutic properties, such as anti-diabetic, antigenic, antibacterial, antifungal, anti-genotoxic, anti-leishmania, anti--inflammatory, anti-Helicobacter, and immunomodulatory.7,8 Auraptene has demonstrated anti-inflammatory and neuroprotective effects in the brains of inflammatory models injected with lipopolysaccharide (LPS).9 Auraptene has alleviated gastritis by reducing production of proinflammatory mediator.10 Auraptene has shown anti-inflammatory characteristics by suppressing the production of inflammatory proteins that mediated association between fat cells and macrophages.10 Auraptene has regulated human platelet activation and prevented arterial thrombosis in mice by inhibiting nuclear factor kappa B (NF-κB) axis11 However, the role of auraptene in pediatric pneumonia has remained unclear.
NF-κB is involved in immune and inflammatory responses and has induced the levels of downstream inflammatory cytokines that mediate inflammatory response in pathological actions.11,12 TNF-α and TNF receptors are expressed on several immune cells and have regulated the survival and activation of T cells.11,13 One of the main causes of lung infection is the presence of LPS that effectively stimulates immune cells.11 Therefore, the NF-κB pathway may serve as a target for treating pneumonia.
In the present study, we proposed to elucidate the effect of auraptene on pneumonia. We constructed LPS-treated WI-38 cell model. Our results revealed that auraptene alleviated inflammatory injury and cell apoptosis in pneumonia. Therefore, auraptene has the potential to act as a pneumonia drug.
Materials and methods
Cell culture
This study was approved by the Ethics Committee of the Affiliated Hospital of Xiangnan University. The human bronchial epithelial cell line WI-38 were obtained from American Type Culture Collection (ATCC) and cultured in Dulbecco's modified eagle medium (DMEM) supplemented with 10% fetal bovine serum (FBS) in a 5% CO2 humidified atmosphere at 37°C. Cells were treated with 5-μg/mL LPS and auraptene (Sigma, St. Louis, MO, US) for 24 h to construct the cell model.
Cell viability
WI-38 cells were plated into 96-well plates. Cell viability was assessed by adding cell counting kit-8 (CCK-8) solution (Beyotime, Shanghai, China). Cells were incubated with CCK-8 solution for 4 h and optical density (OD) was observed through the measurement of OD450.
Enzyme-linked-immunosorbent serologic assay (ELISA)
The concentrations of TNF-α, IL-1β, and IL-6 in cell lysates were measured with ELISA kit (Beyotime, Beijing, China) following manufacturer’s protocol.
Real-Time Polymerase chain reaction (RT-PCR)
Total cellular RNA was extracted with TRIzol reagents (Thermo Fisher, MA, US). Total RNA was reverse-transcribed into complementary DNA (cDNA) using M-MLV reverse transcriptase (Promega, Madison, WI, US). The relative expression of target genes were determined by RT-PCR. The following primers were used: TNF-α: GGTGCCTATGTCTCAGCCTCTT, GCCATAGAACTGATGAGAGGGAG; interleukin 1β IL-1β: ACAAGGAGAAGAAAGTAATGAC, GCTGTAGAGTGGGCTTAT; IL-6: AGACAGCCACTCACC, TTCTGCCAGTGCCTCTT; and glyceraldehyde 3-phosphate dehydrogenase (GAPDH): AGAAGGCTGGGGCTCATTTG, AGGGGCCATCCACAGTCTTC.
Western blot analysis
Proteins were extracted using radioimmunoprecipitation assay (RIPA) buffer (Beyotime). Then, the samples were collected and separated by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE), and transferred on polyvinylidene fluoride (PVDF) membranes. After blocked using 5% fat-free milk, PVDF membranes were incubated with primary antibodies, including Bax (1:1000; ab32503; Abcam, Cambridge, UK), Bcl-2 (1:1000; ab182858; Abcam), p-p65 (1:1000; ab76302; Abcam), p65 (1:1000; ab32536; Abcam), p-IκBα (1:1000; ab133462; Abcam), IκBα (nuclear factor of kappa light polypeptide gene enhancer in B-cells inhibitor, alpha) (1:1000; ab237777; Abcam), and β-actin (1:3000; ab8226; Abcam) for 1 h. The membranes were incubated with secondary antibodies for 1 h. Specific proteins were visualized with enhanced chemiluminescence (ECL) kit (Thermo Fisher).
Cell apoptosis assay
Cell apoptosis kit was bought from Beyotime. Cells were fixed with 70% ethanol for 2 h. Subsequently, cells were stained with propidium iodide (PI) and fluorescein isothiocyanate (FITC) Annexin V at 4°C, and the apoptosis rate was measured. The cell apoptosis was assessed using FACS Calibur flow cytometer and analyzed by CellQuest Pro 5.1 (BD Biosciences, NJ, US).
Statistics
The GraphPad 8.0 software was used for statistical analysis. Data were represented as mean ± SD. The unpaired Student’s t-test was used to determine statistical significance between two groups. P < 0.05 was considered statistically significant.
Results
Auraptene stimulated the cell viability in LPS-stimulated WI-38 cells
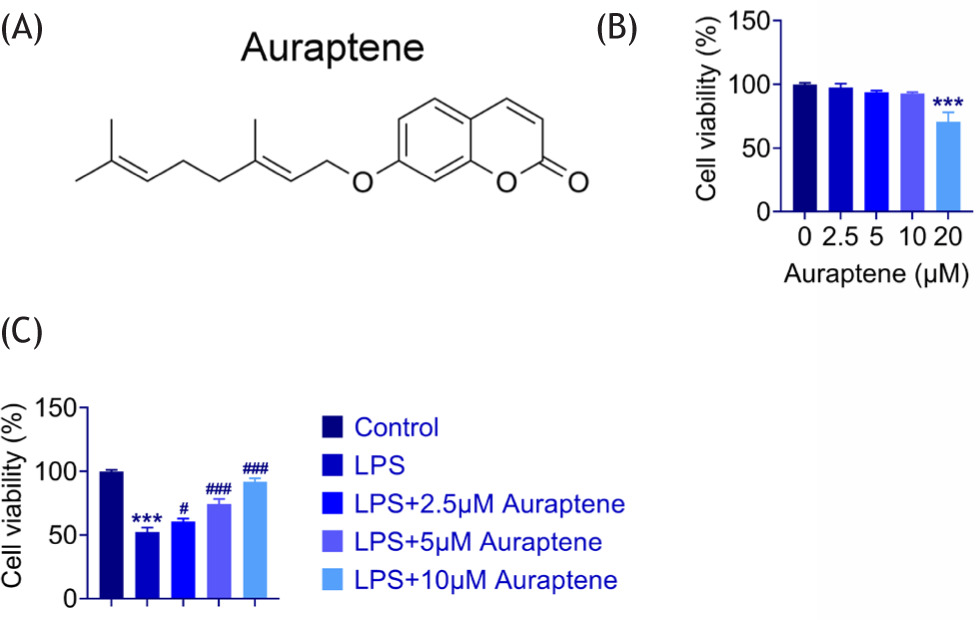
The structure of auraptene is shown in Figure 1A. LPS-stimulated WI-38 cells were constructed as a pneumonia model. To evaluate the role of auraptene in WI-38 cell growth, CCK-8 assay was performed. We found that treatment of WI-38 cells with low concentrations of auraptene (2.5-, 5-, and 10-μM) had modest effect on viability whereas high concentration (20 μM) of auraptene suppressed viability (Figure 1B). Therefore, WI-38 cells were treated with low concentration of auraptene (2.5-, 5-, and 10-μM) in the following experiments. When WI-38 cells were stimulated with 5-μg/mL LPS, cell viability was significantly impaired (Figure 1C). Subsequently, LPS-stimulated WI-38 cells were treated with increasing low concentration (2.5-, 5-, and 10-μM) of auraptene. The addition of auraptene improved cell viability (Figure 1C). Therefore, auraptene stimulated the cell viability of WI-38 cells upon LPS treatment.
Figure 1 Auraptene stimulated cell viability in LPS-induced WI-38 cells. (A) Structure of auraptene. (B) CCK-8 assay showed the cell viability of WI-38 cells exposed to auraptene at the concentrations of 0-, 2.5-, 5-, 10-, and 20-μM for 24 h. (C) CCK-8 assay showed the cell viability of WI-38 cells exposed to LPS and auraptene at a concentration of 0-, 2.5-, 5-, 10-μM for 24 h. ***P < 0.001, LPS vs. control,#P < 0.05, ###P < 0.001, LPS+auraptene vs. LPS.
Auraptene inhibited LPS-induced cell inflammation
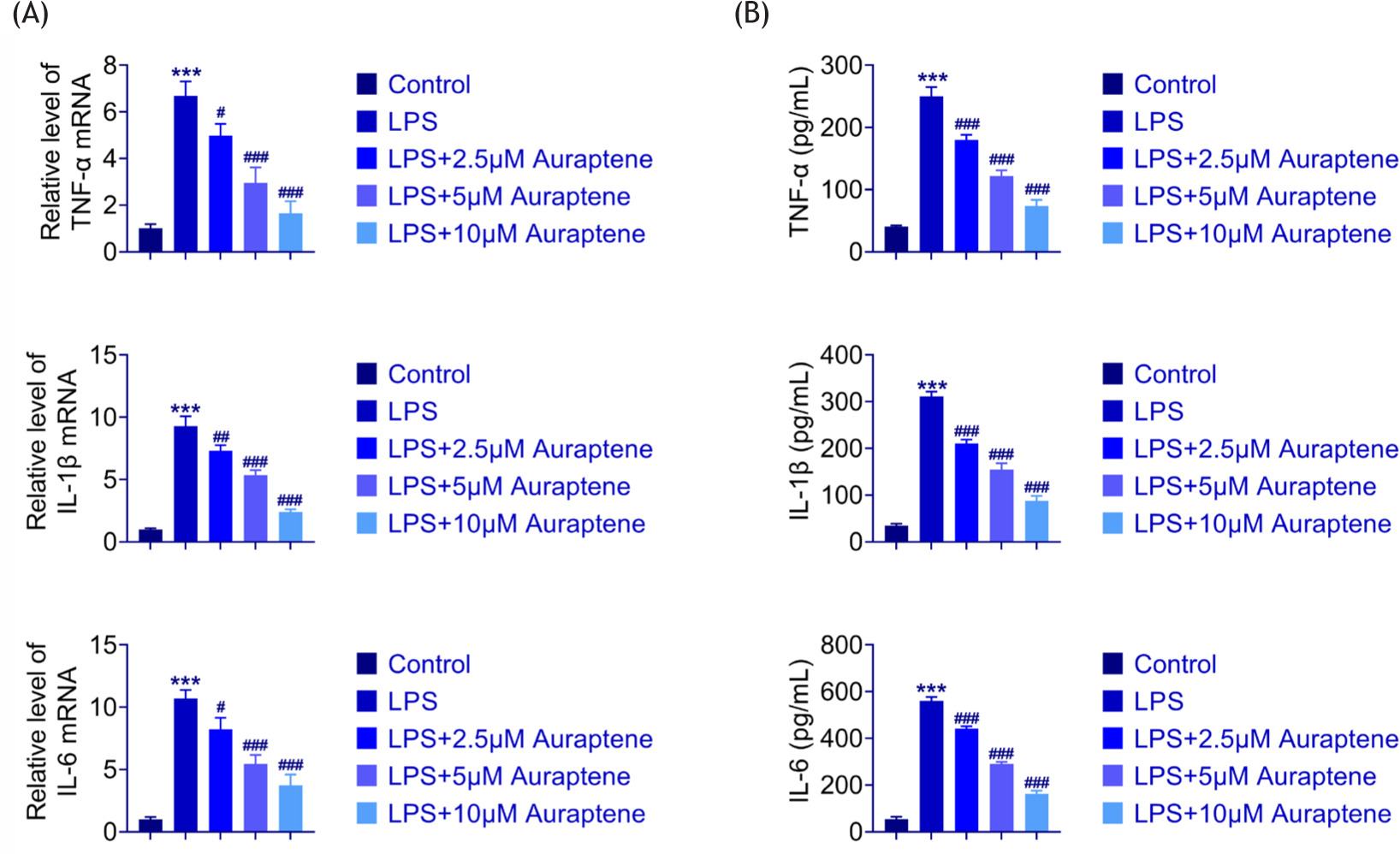
To assess inflammatory gene levels in LPS-stimulated WI-38 cells, the messenger RNA (mRNA) levels of IL-6, IL-1β, and TNF-α were discovered. LPS increased the mRNA levels of these genes through quantitative PCR (qPCR; Figure 2A). We further found that auraptene administration relieved cellular inflammation by reducing the levels of these cytokines (Figure 2A). Consistently, the protein secretion of these factors after auraptene treatment showed the same results (Figure 2B). Therefore, auraptene inhibited LPS-stimulated cell inflammation.
Figure 2 Auraptene inhibited LPS-induced cell inflammation. (A) The mRNA level of TNF-α, IL-1β, and IL-6 in control, LPS, and auraptene+LPS cells. (B) The secretion level of TNF-α, IL-1β, and IL-6 in control, LPS, and auraptene+LPS WI-38 cells. ***P < 0.001, LPS vs. control,#P < 0.05, ##P < 0.01, ###P < 0.001, LPS+auraptene vs. LPS.
Auraptene inhibited LPS-induced cell apoptosis in LPS-induced WI-38 cells
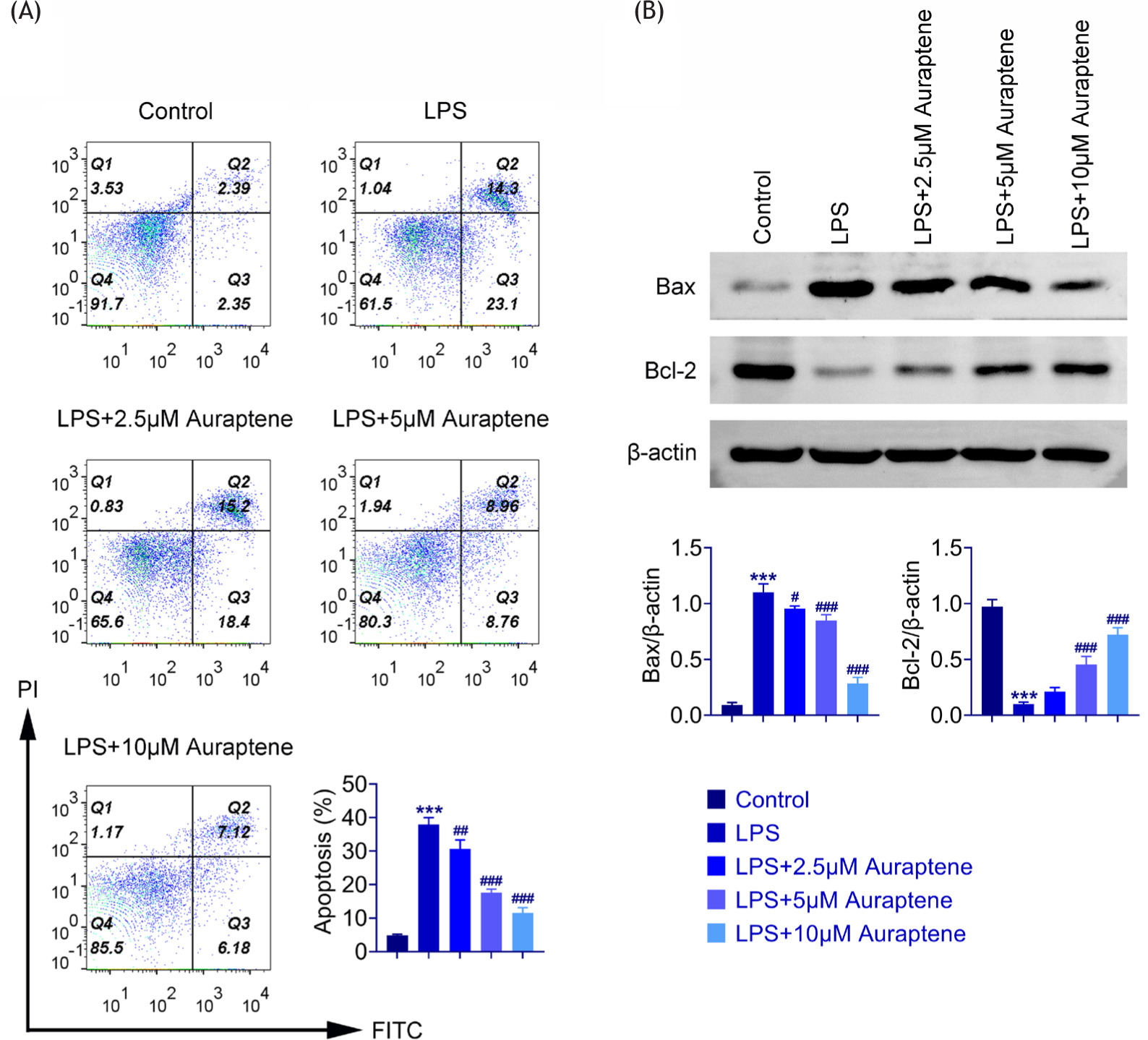
We discovered the effects of auraptene on LPS-induced WI-38 cell apoptosis. The cell apoptosis was observed by flow cytometry (FCM) assay. Induction of LPS in WI-38 cells increased cell apoptosis (Figure 3A). Auraptene treatment significantly reduced the percentage of apoptotic cells (Figure 3A). In addition, the expression levels of Bax and Bcl-2 were detected in WI-38 cells. LPS treatment obviously enhanced Bax levels and reduced Bcl-2 levels (Figure 3B). However, alterations in Bax and Bcl-2 were reversed by auraptene treatment (Figure 3B). Therefore, auraptene inhibited LPS-induced cell apoptosis in WI-38 cells upon LPS treatment.
Figure 3 Auraptene inhibited LPS-induced cell apoptosis in LPS-induced WI-38 cells. (A) FCM assay showed the cell apoptosis of WI-38 cells exposed to LPS and auraptene at the concentrations of 0-, 2.5-, 5-, and 10-μM for 24 h. The percentage of apoptosis cells was quantified. (B) Immunoblot assays showed the expression of Bax and Bcl-2 in WI-38 cells exposed to LPS and auraptene at the concentrations of 0-, 2.5-, 5-, and 10-μM for 24 h. ***P < 0.001, LPS vs. control,#P < 0.05, ##P < 0.01, ###P < 0.001, LPS+auraptene vs. LPS.
Auraptene inhibited NF-κB axis in WI-38 cells upon LPS treatment
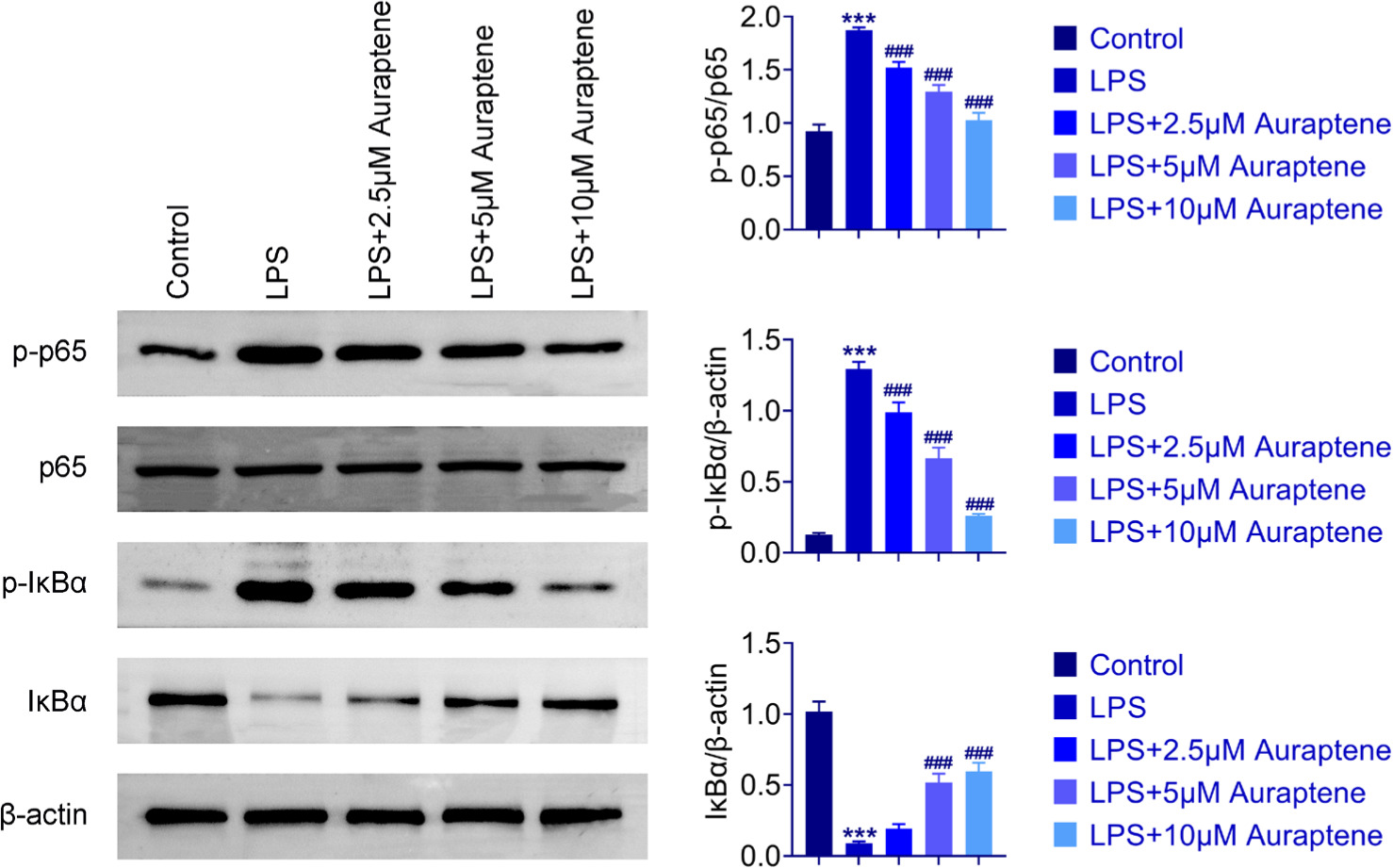
NF-κB signaling pathway was involved in the LPS-stimulated inflammation model. We observed the possible association of auraptene and NF-κB signaling pathway. LPS treatment increased the levels of p65 and IκBα phosphorylation in WI-38 cells, which were the indicators of this pathway, compared to control (Figure 4). However, auraptene reduced the p65 and IκBα phosphorylation levels in WI-38 cells upon LPS treatment (Figure 4). Thus, auraptene restrained NF-κB activation in WI-38 cells upon LPS treatment.
Figure 4 Auraptene inhibited the NF-κB pathway in LPS-induced WI-38 cells. Immunoblot assay showed the expression of indicated proteins in WI-38 cells exposed to LPS and auraptene at the concentrations of 0-, 2.5-, 5-, and 10-μM for 24 h. ***P < 0.001, LPS vs. control,###P < 0.001, LPS+auraptene vs. LPS.
Discussion
Pneumonia is an inflammation of the lungs caused by pathogens invading the alveoli.1 Pneumonia can occur at all ages, but is more common in children aged <2 years.3 For pediatric pneumonia, there is still a lack of drugs, leading to urgent requirement of related research and development work.14 The treatment of pneumonia should focus on inhibiting excessive inflammatory response and alleviating cell apoptosis.15 A variety of drugs have achieved good results in the preclinical treatment of pneumonia, but it is still difficult to meet the current clinical requirements.16 In the present study, we constructed a pneumonia LPS cell model, and found that auraptene could suppress the progression of pneumonia at cellular levels. We conceived that auraptene has the potential to act as a pneumonia drug. In addition, exercise is a rehabilitation strategy with anti-inflammatory effects and could be a good plan to treat pneumonia.1
Inflammation, a self-protective response of vascular living tissue to injury, is closely associated with neurodegenerative and cardiovascular diseases.16 By performing a series of assays, such as CCK-8, FCM, ELISA, and Immunoblot, we found that auraptene alleviated LPS-induced inflammation and apoptosis of WI-38 cells.6 Auraptene, an active secondary metabolite, is primarily isolated from the plants of the Rutaceae family.9,10 Auraptene is effective in treating several disorders, such as keloids and pain.10 Auraptene possesses numerous pharmacological properties, such as anti-hypertensive and anti-cancer.6,11 We discovered anti-inflammatory and pro-apoptosis effects of auraptene in a pneumonia cell model. Auraptene decreases the secretion of MMP-2 and key inflammatory factors.17,18 Auraptene showed wound healing effects and could control periodontal diseases by regulating inflammatory protein secretion.17,18 In addition, auraptene showed anti--inflammatory effects by restraining inflammatory factors that mediate correlation between adipocytes and macrophages.19 Similarly, we also determined that auraptene inhibited LPS-stimulated cellular inflammation.
NF-κB can stimulate the levels of inflammatory cytokines to mediate inflammatory response, and has normal physiological function in mediating immune response.20 p65 and IκBα are two key regulatory proteins of the NF-κB pathway, and their phosphorylation leads to the activation of this pathway. Therefore, we measured the phosphorylation level of these proteins to discover the activation of NF-κB signaling pathway. The inflammatory response of the body needs to activate the NF-κB signaling pathway and transcribe some cytokines to mediate immune response. NF-κB is an important transcriptional regulator that regulates the levels of multiple inflammatory genes.20 It has been shown to regulate the expression of more than 100 genes.20,21 Inflammatory response is often over activated in patients with pneumonia, and NF-κB plays a vital role in regulating cell proliferation and immunity and inflammation.21 Therefore, the NF-κB signaling pathway is widely used as a target for pneumonia, especially in children.22,23 Interestingly, we discovered that auraptene would approach the NF-κB signaling pathway, thereby alleviating lung inflammation and inhibiting apoptosis. However, the detailed molecular mechanism requires further studies.
Conclusion
In summary, we discovered that auraptene alleviated inflammatory injury and cell apoptosis in pneumonia. Therefore, Aauraptene has the potential to act as a drug for the treatment of pneumonia.
Funding
This work was supported by Hunan Provincial Health Commission (Grant No. C202306015991).
Competing interests
The authors stated that there was no conflict of interest to declare.
Ethical approval
This article did not contain any studies performed by any of the authors involving human participants or animals.
Data availability
The authors declare that all data supporting the findings of this study are available within the paper, and any raw data can be obtained from the corresponding author upon request.
Author Contributions
Yuebin Wang and Shuzhen Yuan designed and carried out the study. Yuebin Wang, Shuzhen Yuan, Wei Tan, Yuanyu Zhou, and Ruiyun Liao supervised data collection, analysis, and interpretation. Yuebin Wang, Shuzhen Yuan, and Wei Su prepared and reviewed the draft of the manuscript. All authors had read and approved the final manuscript.
REFERENCES
1. Sanders NL, Martin IMC, Sharma A, Jones MR, Quinton LJ, Bosmann M, et al. Neutrophil extracellular traps as an exacerbating factor in bacterial pneumonia. Infect and Immun. 2022;90(3):e0049121. 10.1128/iai.00491-21
2. Pulavendran S, Rudd JM, Maram P, Thomas PG, Akhilesh R, Malayer JR, et al. Combination therapy targeting platelet activation and virus replication protects mice against lethal influenza pneumonia. Am J Resp Cell Mol Biol. 2019;61(6): 689–701. 10.1165/rcmb.2018-0196OC
3. Zhang J, Zhu Y, Zhou Y, Gao F, Qiu X, Li J, et al. Pediatric adenovirus pneumonia: Clinical practice and current treatment. Front Med. 2023;10:1207568. 10.3389/fmed.2023.1207568
4. Luan Y, Liu H, Luan Y, Yang Y, Yang J, Ren KD. New insight in HDACs: Potential therapeutic targets for the treatment of atherosclerosis. Front Pharmacol. 2022;13:863677. 10.3389/fphar.2022.863677
5. Ghorbani M, Soukhtanloo M, Farrokhi AS, Hassanian SM, Ghorbani F, Afshari AR, et al. Auraptene-induced cytotoxic effects in acute myeloid leukemia cell lines. Med Oncol. 2023;40(8):231. 10.1007/s12032-023-02088-5
6. Goudarzi S, Rassouli FB, Kahrizi D, Paryan S, Mahdifar M, Mehrdad I, et al. Comparing toxicity of galbanic acid, auraptene and umbelliprenin on adult T-cell leukaemia-lymphoma in normoxia and hypoxia. Cell Mol Biol. 2022;68(12):17–20. 10.14715/cmb/2022.68.12.4
7. Kim YH, Lee SY, Kim EY, Kim KH, Koong MK, Lee KA. The antioxidant auraptene improves aged oocyte quality and embryo development in mice. Antioxidants. 2022;12(1). 10.3390/antiox12010087
8. Min JS, Jin YH, Kwon S. Auraptene has antiviral activity against human coronavirus OC43 in MRC-5 cells. Nutrients. 2023;15(13). 10.3390/nu15132960
9. Okuyama S, Minami S, Shimada N, Makihata N, Nakajima M, Furukawa Y. Anti-inflammatory and neuroprotective effects of auraptene, a citrus coumarin, following cerebral global ischemia in mice. Eur J Pharmacol. 2013;699(1–3):118–23. 10.1016/j.ejphar.2012.11.043
10. Sekiguchi H, Takabayashi F, Irie K, Murakami A. Auraptene attenuates gastritis via reduction of helicobacter pylori colonization and pro-inflammatory mediator production in C57BL/6 mice. J Med Food. 2012;15(7):658–63. 10.1089/jmf.2011.1844
11. Hsia CW, Wu MP, Shen MY, Hsia CH, Chung CL, Sheu JR. Regulation of human platelet activation and prevention of arterial thrombosis in mice by auraptene through inhibition of NF-kappaB pathway. Int J Mol Sci. 2020;21(13). 10.3390/ijms21134810
12. Luan Y, Ren KD, Luan Y, Chen X, Yang Y. Mitochondrial dynamics: Pathogenesis and therapeutic targets of vascular diseases. Front Cardiovasc Med. 2021;8:770574. 10.3389/fcvm.2021.770574
13. Ghezelayagh TS, Wu ES, Barber EL, Dao MD, Zsiros E, Urban RR, et al. Timing and duration of bevacizumab treatment and survival in patients with recurrent ovarian, fallopian tube, and peritoneal cancer: A multi-institution study. Eur J Gynaecol Oncol. 2023;44(1):17–25.
14. Sayama Y, Okamoto M, Saito M, Tamaki R, Saito-Obata M, Quicho RFN, et al. Lack of zoonotic coronavirus species detected among children hospitalized with pneumonia in the Philippines. Clin Infect Dis. 2023. 10.1093/cid/ciad430
15. Guz D, McNeil R, Buchrits S, Goshen S, Gafter-Gvili A, Avni T. Ceftriaxone 1 g versus 2 g per day, for the treatment of community-acquired pneumonia: a retrospective cohort study. internal and emergency medicine. 2023. 10.1007/s11739-023-03372-y
16. Song S, Xu Y. A retrospective study of the clinical characteristics of 9 children with pulmonary embolism associated with mycoplasma pneumoniae pneumonia. BMC Pediat. 2023;23(1):370. 10.1186/s12887-023-04188-7
17. Gao X, Fu T, Wang C, Ning C, Kong Y, Liu Z, et al. Computational discovery and experimental verification of farnesoid X receptor agonist auraptene to protect against cholestatic liver injury. Biochem Pharmacol. 2017;146:127–38. 10.1016/j.bcp.2017.09.016
18. La VD, Zhao L, Epifano F, Genovese S, Grenier D. Anti-inflammatory and wound healing potential of citrus auraptene. J Med Food. 2013;16(10):961–4. 10.1089/jmf.2013.0029
19. Takahashi N, Senda M, Lin S, Goto T, Yano M, Sasaki T, et al. Auraptene regulates gene expression involved in lipid metabolism through PPAR alpha activation in diabetic obese mice. Mol Nutrit Food Res. 2011;55(12):1791–7. 10.1002/mnfr.201100401
20. Ni Ww, Zhang Qm, Zhang X, Li Y, Yu Ss, Wu Hy, et al. Modulation effect of Lactobacillus acidophilus KLDS 1.0738 on gut microbiota and TLR4 expression in β-lactoglobulin-induced allergic mice model. Allergol Immunopathol. 2020;48(2): 149–57. 10.1016/j.aller.2019.06.002
21. Li J, Yan W, Ren F, Sang H. Tectorigenin inhibits inflammation in keratinocytes by inhibition of NLRP3 inflammasome regulated by the TLR4/NF-kappaB pathway. Allergol Immunopathol (Madr). 2023;51(2):82–9. 10.15586/aei.v51i2.780
22. Pandey P, Al Rumaih Z, Kels MJT, Ng E, Kc R, Malley R, et al. Therapeutic targeting of inflammation and virus simultaneously ameliorates influenza pneumonia and protects from morbidity and mortality. Viruses. 2023;15(2). 10.3390/v15020318
23. Wang Y, Jian S, Li W, Zhao L, Ye G, Shi F, et al. Epigallocatechin-3-gallate ameliorates liver injury secondary to pseudomonas aeruginosa pneumonia. Int Immunopharmacol. 2022;112:109239. 10.1016/j.intimp.2022.109239