
Download
ORIGINAL ARTICLE
Chryseriol attenuates the progression of OVA-induced asthma in mice through NF-κB/HIF-1α and MAPK/STAT1 pathways
Shangyao Moa, Hao Denga, Yong Xiea, Lixia Yanga, Lili Wenb*
aDepartment of Respiratory and Critical Care Medicine, Nanchong Central Hospital, Nanchong, Sichuan, China
bDepartment of Respiratory and Critical Care Medicine, Lishui City People’s Hospital, Lishui, Zhejiang, China
Abstract
Background: Asthma is a hackneyed chronic inflammatory disease of the airway. Chryseriol (CSR) is a kind of flavonoid, and has the effect of bronchiectasis, indicating its potential application for treating respiratory diseases. However, the functions of CSR in asthma have not been reported till now.
Materials and methods: The histopathologic changes of the lung tissues were assessed by hematoxylin and eosin staining. The cell apoptosis was identified through terminal deoxynucleotidyl transferase-mediated dUTP nick end labelling assay. Total numbers of eosinophils, neutrophils, and macrophages were assessed under microscope. The levels of interleukin (IL)-1β, IL-4, IL-5, and IL-13 were detected by enzyme-linked-immunosorbent serologic assay. The airway hyper-responsiveness (AHR) was evaluated by the whole body plethysmography. The levels of methane dicarboxylic aldehyde, superoxide dismutase, glutathione S-transferase, and glutathione in lung homogenates were confirmed by using corresponding commercial kits. The protein expressions were examined by Western blot analysis.
Results: The ovalbumin (OVA) was utilized to establish asthma mouse model. At first, it was revealed that CSR treatment reduced lung injury in OVA-stimulated mice. Moreover, cell apoptosis was enhanced after OVA stimulation but was attenuated by CSR treatment. In addition, CSR treatment decreased the infiltration of inflammatory cells and the production of inflammatory factors in OVA-treated mice. Further investigations demonstrated that CSR treatment relieved AHR in OVA-stimulated mice. The oxidative stress was strengthened in OVA-treated mice, but these effects were relieved by CSR treatment. Lastly, it was discovered that CSR treatment retarded nuclear factor kappa B (NF-κB)/hypoxia-inducible factor 1 alpha (HIF-1α) and p38 mitogen-activated protein kinase (MAPK)/signal transducer and activator of transcription 1 (STAT1) pathways in OVA-triggered asthma mice.
Conclusion: Our findings proved that CSR attenuated the progression of OVA-induced asthma in mice through inhibiting NF-κB/HIF-1α and MAPK/STAT1 pathways. This work might highlight the functions of CSR in the treatment of asthma.
Key words: asthma, chryseriol, NF-κB/HIF-1α and MAPK/STAT1 pathways, ovalbumin
*Corresponding author: Lili Wen, Department of Respiratory and Critical Care Medicine, Lishui City People’s Hospital, No. 15 Dazhong Street, Lishui City, Liandu District, Zhejiang 323000, China. Email address: [email protected]
Received 14 September 2022; Accepted 18 October 2022; Available online 1 January 2023
Copyright: Mo S, et al.
License: This open access article is licensed under Creative Commons Attribution 4.0 International (CC BY 4.0). http://creativecommons.org/licenses/by/4.0/
Introduction
Asthma is a kind of chronic respiratory disease characterized by reversible airflow obstruction caused by bronchial hyper-responsiveness because of chronic airway inflammation, which can be triggered by a multitude of agents, including allergens, infections, and pollutants.1,2 The common symptoms of asthma are dyspnea, chest tightness, wheezing, and cough.3 Airway remodeling is interrelated with extracellular matrix proteins in patients with severe asthma.4 Asthma can affect people at any age, special with increasing morbidity and mortality in children aged less than 18 years.5,6 Most patients are treated with low to moderate doses of inhaled corticosteroids to relieve asthma symptoms, although some patients with severe asthma require more intensive treatment, such as inhaled steroids, long-acting β2 agonists, or leukotriene antagonists as well as combined treatment.7–9 The biological treatment (e.g., omalizumab) has also been utilized as asthma therapy,10 although in spite of treatment, asthma symptoms are not controlled. Therefore, it is crucial to deeply realize the mechanism of asthma and look for novel therapeutic drugs.
Flavonoids, widely found in plants, belong to secondary metabolites of plants, and are believed to affect the progression of diseases, including asthma.11,12 Chryseriol (CSR) is one flavonoid that exists in diversified diets and herbs, such as Coix seed, honeysuckle, salvia miltiorrhiza, etc.13 CSR has certain pharmacological effects, such as modulating the immune system, reducing obesity, and anti-oxidation.14–16 CSR suppresses the activation of nuclear factor kappa B (NF-κB) and activator protein 1 (AP-1) to ameliorate toll-like receptor 4 (TLR4)-mediated inflammation.17 CSR improves tissue plasminogen activator (TPA)-stimulated acute skin inflammation in mice, and retards NF-κB and signal transducer and activator of transcription 3 (STAT3) pathways.18 CSR retards AP-1 activation to reduce lipopolysaccharide (LPS)-triggered nitric oxide production in macrophages, then incurring anti-inflammatory effects.19 Previous study has illuminated that CSR has some effects for selective bronchiectasis, indicating its potential application for the treatment of respiratory diseases.20 Nevertheless, the specific impact of CSR on asthma has not been elaborated.
The aim of this work was to investigate the role of CSR in ovalbumin (OVA)-induced asthma. In this study, it was revealed that CSR attenuated the progression of OVA-induced asthma in mice by inhibiting NF-κB/ hypoxia--inducible factor 1 alpha (HIF-1α) and mitogen-activated protein kinase (MAPK)/STAT1 pathways. This finding may contribute for the treatment of asthma.
Materials and methods
OVA mice model
Female BALB/c mice (aged 6–8 weeks, n = 25) were bought from Vital River (Beijing, China). The mice were allowed to acquire free food and water, and kept under the 12-h light/dark cycle at 25°C. Ethical approval was obtained from the Ethics Committee of Nanchong Central Hospital, and all animal experiments were conducted according to the guidelines of the Animal Experiments Ethics Committee.
Mice were randomly divided into the following five groups (n = 5 in each group): control, OVA, CSR (20 mg/kg), OVA+CSR (20 mg/kg), and OVA+CSR (40 mg/kg). The OVA-induced asthma mouse model was established as described by Casaro et al. and Dogan et al.21,22 The mice were sensitized with 20-μg OVA (Sigma Aldrich, USA) and aluminum hydroxide (2 mg; Thermo Fisher Scientific, MA, USA) in phosphate-buffered saline (PBS, 200 μL) solution intraperitoneally on day 0, 7, and 14. Next, on day 21, 22, and 23, the mice were treated with 5% OVA inhalation for 30 min each day. In the control group, the mice were subjected to PBS alone. Mice were intraperitoneally injected with CSR (20 mg/kg and 40 mg/kg body weight) from day 17 to 23. The mice were sacrificed after 24 h of post last challenge. Then, bronchoalveolar lavage fluid (BALF) and lung tissues were acquired for succeeding experiments.
Hematoxylin and eosin (H&E) staining
Lung tissues were fixed in 4% paraformaldehyde. Next, they were conducted to dehydration and paraffin-embedding, and cut into sections. The sections were stained with H&E stain solution. Finally, the histopathologic changes in lung tissues were captured under light microscope.
Terminal deoxynucleotide transferase-mediated dUTP nick end-labeling (TUNEL) assay
The in situ cell death detection kit (Cat. No. 11684817910, Roche, Basel, Switzerland) was used to assess cell apoptosis. The lung sections were permeabilized with 0.1% Triton X-100 and 0.1% sodium citrate. The TUNEL staining reagent was added for incubation in darkness at 37°C for 1 h. After washing, the sections were incubated with converter--peroxidase (POD) and diaminobenzidine (DAB). Finally, the fluorescent images were derived under one fluorescence microscope (Olympus, Japan).
Detection of eosinophils, neutrophils, and macrophages
The eosinophils, neutrophils, and macrophages present in BALF were evaluated through Giemsa staining. In general, cells in BALF were resuspended in PBS (0.1 mL), and BALF (10 μL) was put onto a glass slide, which was covered with a glass coverslip. Methyl alcohol was used to fix the slides, and the slides were dried. Next, slides were dyed by Wright–Giemsa staining solution (Nanjing Jiancheng Bioengineering Institute, Nanjing, China). The calculation of cell numbers was carried out under a light microscope.
Enzyme-linked-immunosorbent serologic assay (ELISA)
The levels of IL-1β (ab197742), IL-4 (ab100710), IL-5 (ab204523), and IL13 (ab219634) in BALF were measured by using commercial ELISA kits (Abcam, Shanghai, China) according to the manufacturer’s instructions.
Detection of AHR
The mice were anesthetized with pentobarbital sodium (70 mg/kg intraperitoneally) and put on a hot pad to assess airway hyper-responsiveness (AHR). The animals were tracheostomized with 18G cannula ventilator (RWD Inc., Shenzhen, China). Next, they were administrated with the increased concentrations of methacholine (MCh; 0, 2.5, 5, 10, 25, 50 mg/mL), and assessed through the body plethysmograph (Buxco Electronics Inc., Wilmington, NC, USA). Data were presented as airway resistance (RI).
Detection of methane dicarboxylic aldehyde (MDA), superoxide dismutase (SOD), glutathione S-transferase (GST), and glutathione (GSH)
The levels of MDA, SOD, GST, and GSH in lung homogenates were examined using commercial kits bought from the Beyotime Institute of Biotechnology (Shanghai, China) according to manufacturer’s protocols.
Western blot analysis
Proteins were isolated from lung tissues through radioimmunoprecipitation assay (RIPA) lysis buffer (Thermo Fisher Scientific). The proteins were separated by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE), and transferred onto polyvinylidene fluoride (PVDF) membranes (Beyotime). After sealing, the membranes were mixed with the following primary antibodies for 12 h at 4°C: nuclear factor erythroid 2-related factor 2 (Nrf2, 1:1000; ab137550; Abcam), Lamin B1 (0.1 µg/mL; ab16048), HO-1 (1:2000; ab13243), p-p65 (1:1000; ab76302), p65 (0.5 µg/mL; ab16502), HIF-1α (1:500; ab51608), p-p38 (1:1000; ab178867), p38 (1:10,000; ab45136), phospho (p)-STAT1 (1:1000; ab109461), STAT1 (1:1000; ab47425), and β-actin (1 µg/mL; ab8226). Next, the appropriate secondary antibodies (1:2000; ab7090; Abcam) were added on the membranes. Lastly, the chemiluminescence detection kit (Thermo Fisher Scientific) was utilized to evaluate blots.
Statistical analysis
The SPSS 20.0 software (IBM, USA) was applied for statistical analysis. The data were expressed as mean ± standard deviation (SD). All experiments were carried out in triplicate. Comparisons in groups were done by Student’s t-test (between two groups), or one-way ANOVA (for more than two groups) with Turkey test. P ˂ 0.05 was regarded as a significant difference.
Results
CSR treatment reduced lung damage in OVA-stimulated mice
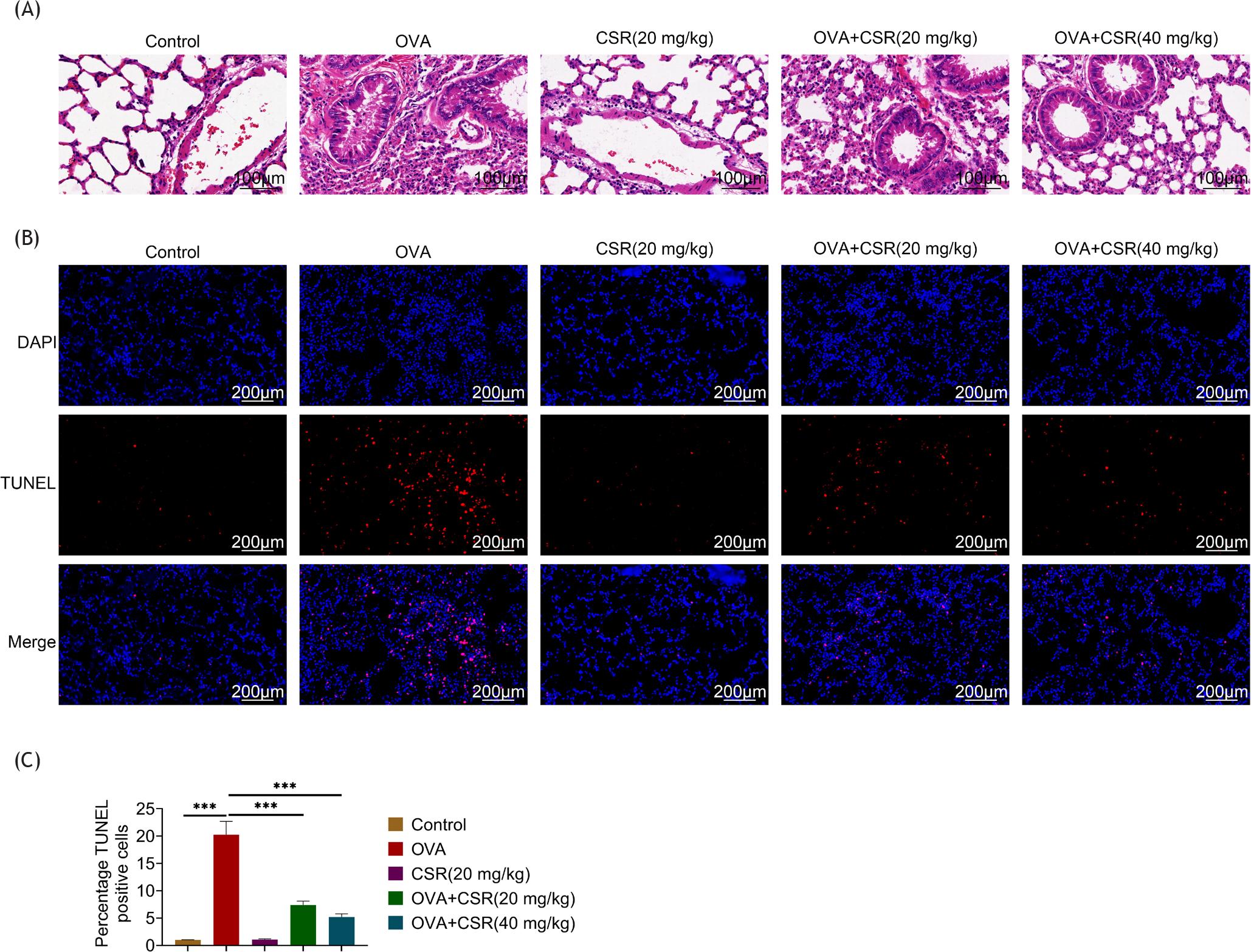
Owing to H&E staining in OVA model group, bronchial wall was thickened and damaged, airway lumen was narrowed, and eosinophil infiltration was increased. In addition, damage to the lung was gradually improved with the increased dose of CSR (Figure 1A). Next, cell apoptosis in lung tissues was examined by TUNEL assay. Results illustrated that cell apoptosis was enhanced after OVA induction but attenuated by CSR treatment (control: 1; OVA: 20.25; CSR (20 mg/kg): 1.06; OVA+CSR (20 mg/kg): 7.38; and OVA+CSR (40 mg/kg): 5.19; ***P ˂ 0.001; Figures 1B and C). These data demonstrated that CSR treatment reduced damage to the lung in OVA-stimulated mice.
Figure 1 CSR treatment reduced the lung damage in OVA-stimulated asthma mice. (A) The lung tissues were examined by H&E staining. (B) and (C) The cell apoptosis in lung tissues was evaluated by TUNEL assay. ***P ˂ 0.001.
CSR treatment decreased infiltration of inflammatory cells and production of inflammatory factors in OVA-induced mice
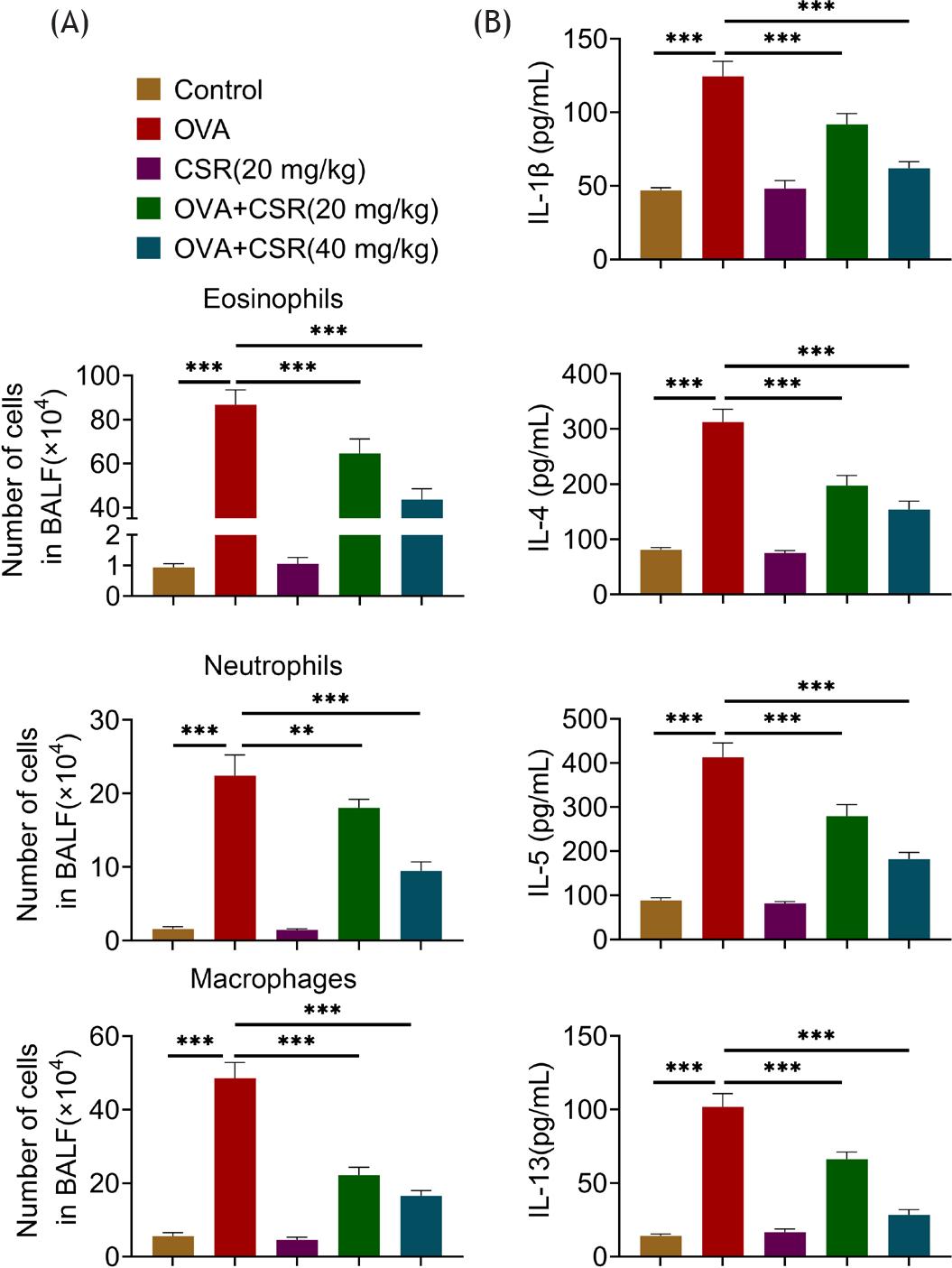
The increased number of eosinophils, neutrophils, and macrophages in OVA-induced mice were decreased by CSR treatment (**P ˂ 0.01 and ***P ˂ 0.001; Figure 2A). Furthermore, the levels of inflammatory factors (IL-1β, IL-4, IL-5, and IL-13) were up-regulated in OVA-induced mice, but these effects were offset by CSR treatment (***P ˂ 0.001; Figure 2B). Taken together, CSR treatment decreased the infiltration of inflammatory cells and the production of inflammatory factors in OVA-induced mice.
Figure 2 CSR treatment decreased the infiltration of inflammatory cells and the production of inflammatory factors in OVA-induced asthma mice. (A) Numbers of eosinophils, neutrophils, and macrophages were assessed under microscope. (B) The levels of IL-1β, IL-4, IL-5, and IL-13 were detected by ELISA. ***P ˂ 0.001.
CSR treatment relieved AHR in OVA-stimulated mice
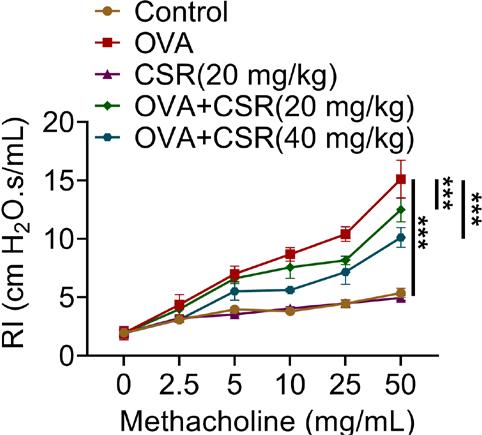
Lung inspiratory resistance index was increased after OVA stimulation, which was rescued by CSR treatment (at 50 methacholine, control: 5.38; OVA: 15.12; CSR (20 mg/kg): 4.95; OVA+CSR (20 mg/kg): 12.50; and OVA+CSR (40 mg/kg): 10.13; ***P ˂ 0.001; Figure 3). These findings suggested that CSR treatment relieved AHR in OVA-stimulated mice.
Figure 3 CSR treatment relieved the AHR in OVA-stimulated asthma mice. The airway resistance (RI) in mice was evaluated through the whole body plethysmography. ***P ˂ 0.001.
CSR treatment attenuated oxidative stress in OVA-stimulated mice
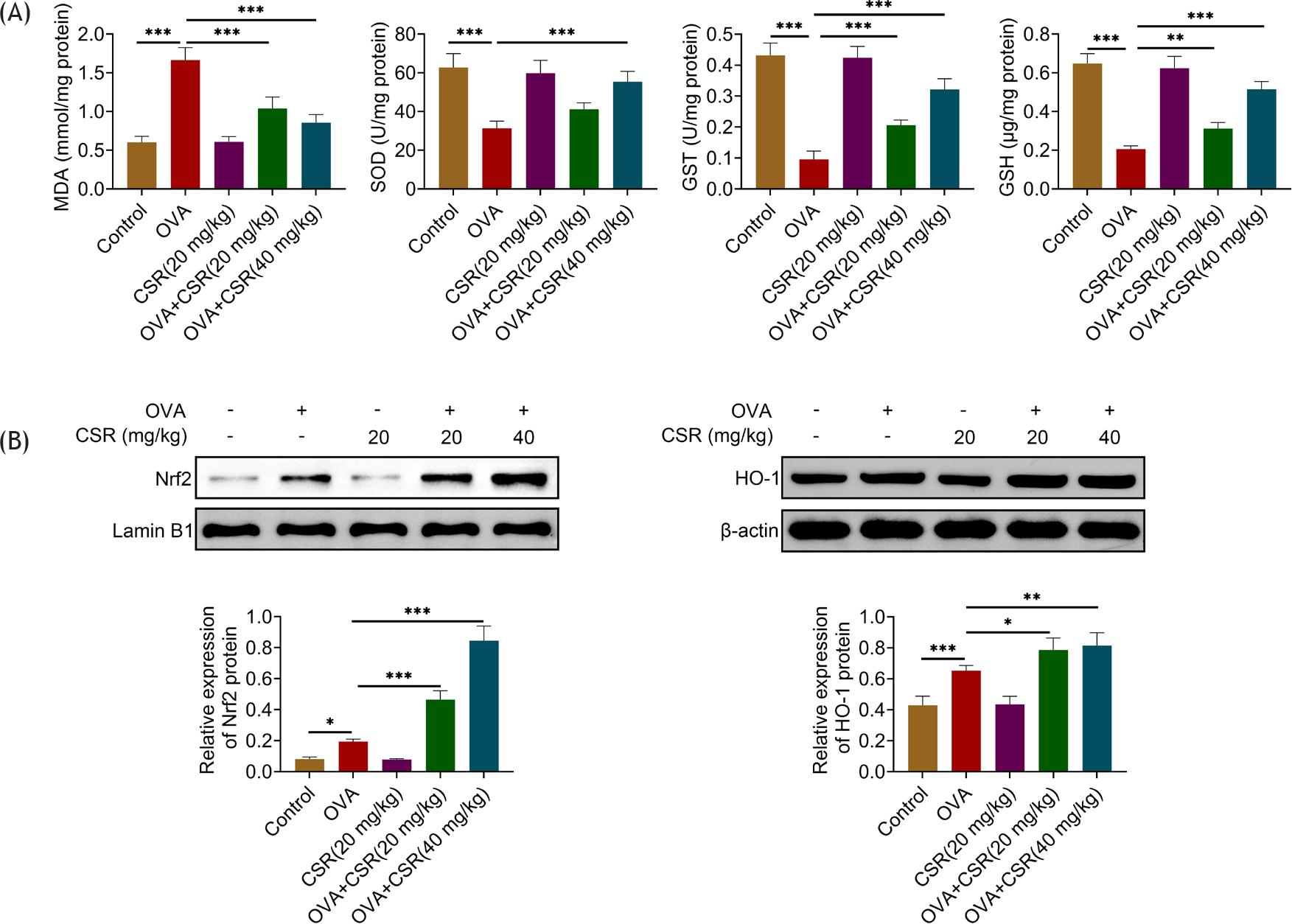
The increased MDA level as well as the decreased SOD, GST, and GSH levels in OVA-stimulated mice was reversed by CSR treatment (**P ˂ 0.01 and ***P ˂ 0.001; Figure 4A). Additionally, the Nrf2 and HO-1 protein expressions were enhanced in mice with OVA stimulation, and these effects were further strengthened by CSR treatment (*P ˂ 0.05, **P ˂ 0.01, and ***P ˂ 0.001; Figure 4B). In general, CSR treatment affected oxidative stress in OVA-stimulated mice.
Figure 4 CSR treatment attenuated oxidative stress in OVA-stimulated asthma mice. (A) The oxidative stress markers (MDA, SOD, GST, and GSH) were examined through the corresponding commercial kits. (B) The protein expressions of Nrf2 (nuclear) and HO-1 were tested by Western blot analysis. *P ˂ 0.05, **P ˂ 0.01, ***P ˂ 0.001.
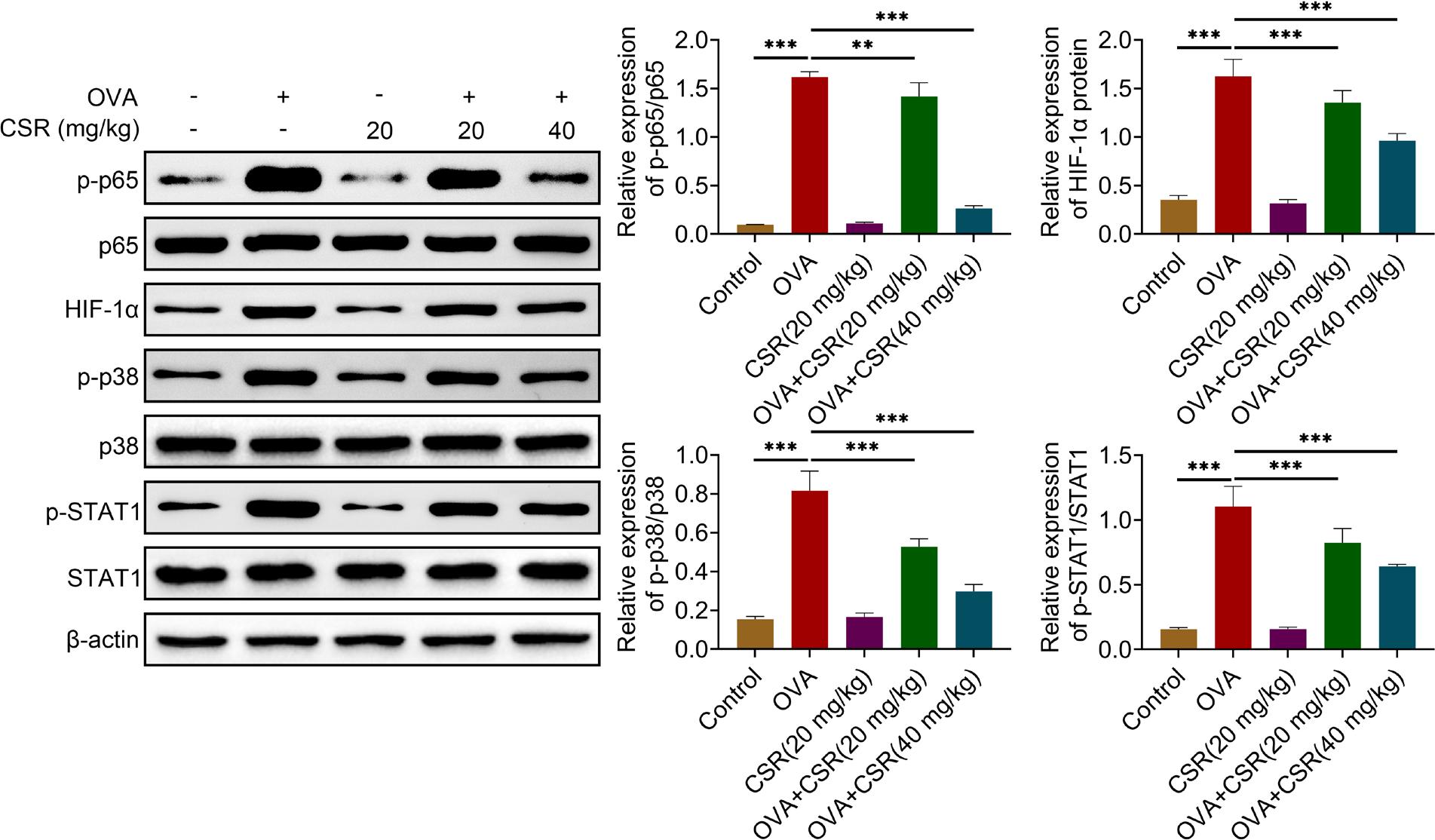
CSR treatment modulated NF-κB/HIF-1α and p38 MAPK/STAT1 pathways in OVA-triggered asthma mice
Finally, NF-κB/HIF-1α and p38 MAPK/STAT1 pathways were evaluated. The expressions of p-p65/p65, HIF-1α, p-p38/p38, and p-STAT1/STAT1 in OVA-treated mice were decreased by CSR treatment (**P ˂ 0.01 and ***P ˂ 0.001; Figure 5). These results revealed that CSR treatment modulated NF-κB/HIF-1α and p38 MAPK/STAT1 pathways in OVA-treated mice.
Figure 5 Schematic diagram of CSR-mediated inhibitive mechanism in asthma progression. CSR treatment modulated NF-κB/HIF-1α and p38 MAPK/STAT1 pathways to attenuate the progression of OVA-induced asthma in mice. The protein expressions of p-p65, p65, HIF-1α, p-p38, p38, p-STAT1, and STAT1 were assessed by Western blot analysis. **P ˂ 0.01, ***P ˂ 0.001.
Discussion
First, it was revealed that CSR treatment reduced lung damage in OVA-stimulated mice. Moreover, cell apoptosis was enhanced by OVA stimulation, but was attenuated by CSR treatment. In addition, CSR treatment decreased the infiltration of inflammatory cells and the production of inflammatory factors in OVA-induced asthma mice. Further investigations demonstrated that CSR treatment relieved AHR in OVA-stimulated mice. The oxidative stress was strengthened in OVA-induced mice, but this effect was relieved by CSR treatment. Lastly, it was discovered that CSR treatment retarded the activation of NF-κB/HIF-1α and p38 MAPK/STAT1 pathways in OVA-treated mice.
Chryseriol therapy has been investigated to be effective for treating many diseases. For instance, CSR regulates phosphatidylinositol 3-kinase–protein kinase B (PI3K/Akt) pathway to affect mitochondrial protection in 1--methyl- 4-phenylpyridinium (MPP+)-treated SH-SY5Y cells.23 In addition, CSR retards Janus kinase 2 (JAK2)/STAT3 signaling and suppresses hyperproliferation of rheumatoid arthritis fibroblast-like synoviocytes.24 Besides, CSR down-regulates early growth response 1 (EGR-1) to reduce TNF-α-triggered CYP19 expression in breast cancer cells.25 Importantly, CSR has been disclosed to have some effects for selective bronchiectasis,20 suggesting its potential role for treating respiratory diseases. However, its role in asthma needs further exploration. In this work, OVA were utilized to establish asthma mouse model. At first, it was manifested that CSR treatment reduced lung damage in OVA-stimulated mice. Moreover, cell apoptosis was enhanced after OVA induction but attenuated by CSR treatment.
Inflammation and oxidative stress are important phenotypes in the progression of asthma. For example, α1--anti-trypsin represses autophagy to ameliorate inflammation and oxidative stress in asthma.26 Furthermore, adiponectin regulates adenosine monophosphate-activated protein kinase (AMPK) pathway in obesity-related asthma mice to reduce airway inflammation and oxidative stress.27 In asthma, fucoxanthin alleviates oxidative stress and inflammation.28 Additionally, 5-aminosalicylic acid improves allergen-induced inflammation and oxidative stress in asthma.29 In this work, it was discovered that CSR treatment decreased the infiltration of inflammatory cells and the production of inflammatory factors in OVA-induced asthma mice. Further investigations demonstrated that CSR treatment relieved AHR in OVA-stimulated mice. Oxidative stress was strengthened in OVA-induced asthma mice, but these effects were relieved by CSR treatment.
The activation of NF-κB/HIF-1α and MAPK/STAT1 pathways can accelerate the occurrence and development of asthma. For instance, IL-1β enhances Muc5ac expression in asthma by affecting NF-κB-triggered HIF-1α expression.30 Eupatilin retards NF-κB/MAPK and activates Nrf2 pathway to improve OVA-induced asthma.31 Furthermore, M2 macrophage-derived exosomal miR-370 suppresses the fibroblast growth factor 1 (FGF1)/MAPK/STAT1 axis to control progression of asthma.32 Interestingly, CSR had an anti--inflammatory role by inactivating NF-κB and MAPK signaling.17 Therefore, we hypothesized that CSR can play an anti-asthmatic role by modulating NF-κB/HIF-1α and MAPK/STAT1 pathways. In this study, it was discovered that CSR treatment retarded NF-κB/HIF-1α and p38 MAPK/STAT1 pathways in OVA-triggered asthma mice.
Conclusion
For the first time, it was proved that CSR attenuated the progression of OVA-induced asthma in mice by inhibiting NF-κB/HIF-1α and MAPK/STAT1 pathways. Our work provided new ideas for searching therapeutic drug for treating asthma. Some limitations to this study included deficient human samples and other cellular progression (autophagy, glycolysis, mitochondrial membrane potential, and macrophage polarization). Interestingly, different allergenic stimulation has been reported to establish other murine model of allergen-specific T helper 2 (Th2)-related asthma.33,34 In the future, more explorations about CSR in asthma can be made to elucidate its other roles.
Availability of Data and Materials
All data generated or analyzed during this study are included in this published article.
Competing Interests
The authors stated that there were no conflicts of interest to disclose.
Author Contributions
Shangyao Mo and Hao Deng designed the study, completed experiments, and supervised data collection. Yong Xie analyzed and interpreted the collected data. Lixia Yang and Lili Wen prepared and reviewed the draft manuscript for publication. All authors read and approved the final manuscript.
REFERENCES
1. Papi A, Brightling C, Pedersen SE, Reddel HK. Asthma. Asthma. Lancet. 2018:391(10122):783–800. 10.1016/S0140-6736(17)33311-1
2. Poddighe D, Brambilla I, Licari A, Marseglia GL. Omalizumab in the therapy of pediatric asthma. Recent Pat Inflamm Allergy Drug Discov. 2018;12(2):103–109. 10.2174/1872213X12666180430161351
3. Gordon BR. Asthma history and presentation. Otolaryngol Clin North Am, 2008:41(2):375–85, vii–viii. 10.1016/j.otc.2007.11.007
4. Munakata M. Airway remodeling and airway smooth muscle in asthma. Allergol Int. 2006:55(3):235–43. 10.2332/allergolint.55.235
5. Guilbert TW, Bacharier LB, Fitzpatrick AM. Severe asthma in children. J Allergy Clin Immunol Pract. 2014:2(5):489–500. 10.1016/j.jaip.2014.06.022
6. Haktanir Abul M, Phipatanakul W. Severe asthma in children: Evaluation and management. Allergol Int. 2019;68(2):150–157. 10.1016/j.alit.2018.11.007
7. Creticos PS. Treatment options for initial maintenance therapy of persistent asthma: A review of inhaled corticosteroids and leukotriene receptor antagonists. Drugs. 2003;63(Suppl 2):1–20. 10.2165/00003495-200363002-00002
8. Chauhan BF, Chartrand C, Ni Chroinin M, Milan SJ, Ducharme FM. Addition of long-acting beta2-agonists to inhaled corticosteroids for chronic asthma in children. Coch Database Syst Rev. 2015;2015(11):Cd007949. 10.1002/14651858.CD007949.pub2
9. Lommatzsch M, Buhl R, Korn S. The treatment of mild and moderate asthma in adults. Dtsch Arztebl Int. 2020;117(25):434–444. 10.3238/arztebl.2020.0434
10. Trivedi M, Denton E. Asthma in children and adults-what are the differences and what can they tell us about asthma? Front Pediatr. 2019;7:256. 10.3389/fped.2019.00256
11. Tanaka T, Takahashi R. Flavonoids and asthma. Nutrients. 2013;5(6):2128–43. 10.3390/nu5062128
12. Maleki SJ, Crespo JF, Cabanillas B. Anti-inflammatory effects of flavonoids. Food Chem. 2019;299:125124. 10.1016/j.foodchem.2019.125124
13. Aboulaghras S, Sahib N, Bakrim S, Benali T, Charfi S, Guaouguaou FE, et al. Health benefits and pharmacological aspects of chrysoeriol. Pharmaceuticals (Basel). 2022; 15(8):973. 10.3390/ph15080973
14. Mishra B, Priyadarsini KI, Kumar MS, Unnikrishnan MK, Mohan H. Effect of O-glycosilation on the antioxidant activity and free radical reactions of a plant flavonoid, chrysoeriol. Bioorg Med Chem. 2003;11(13):2677–85. 10.1016/S0968-0896(03)00232-3
15. Han LK, et al. Anti-obesity action of Salix matsudana leaves (Part 2). Isolation of anti-obesity effectors from polyphenol fractions of Salix matsudana. Phytother Res. 2003;17(10):1195–8. 10.1002/ptr.140510.1002/ptr.1404
16. Skaltsa H, Bermejo P, Lazari D, Silvan AM, Skaltsounis AL, Sanz A, et al. Inhibition of prostaglandin E2 and leukotriene C4 in mouse peritoneal macrophages and thromboxane B2 production in human platelets by flavonoids from Stachys chrysantha and Stachys candida. Biol Pharm Bull. 2000;23(1):47–53. 10.1248/bpb.23.47
17. Yoon HS, Park CM. Chrysoeriol ameliorates COX-2 expression through NF-κB, AP-1 and MAPK regulation via the TLR4/MyD88 signaling pathway in LPS-stimulated murine macrophages. Exp Ther Med. 2021;22(1):718. 10.3892/etm.2021.10150
18. Wu JY, Chen YJ, Bai L, Liu YX, Fu XQ, Zhu PL, et al. Chrysoeriol ameliorates TPA-induced acute skin inflammation in mice and inhibits NF-κB and STAT3 pathways. Phytomedicine. 2020;68:153173. 10.1016/j.phymed.2020.153173
19. Choi DY, Lee JY, Kim MR, Woo ER, Kim YG, Kang KW. Chrysoeriol potently inhibits the induction of nitric oxide synthase by blocking AP-1 activation. J Biomed Sci. 2005;12(6):949–59. 10.1007/s11373-005-9028-8
20. Khan AU, Gilani AH. Selective bronchodilatory effect of Rooibos tea (Aspalathus linearis) and its flavonoid, chrysoeriol. Eur J Nutr. 2006;45(8):463–9. 10.1007/s00394-006-0620-0
21. Casaro M, Souza VR, Oliveira FA, Ferreira CM. OVA-induced allergic airway inflammation mouse model. Methods Mol Biol. 2019;1916:297–301. 10.1007/978-1-4939-8994-2_28
22. Dogan MF, Parlar A, Cam SA, Tosun EM, Uysal F, Arslan SO. Glabridin attenuates airway inflammation and hyperresponsiveness in a mice model of ovalbumin-induced asthma. Pulm Pharmacol Ther. 2020;63:101936. 10.1016/j.pupt.2020.101936
23. Limboonreung T, Tuchinda P, Chongthammakun S. Chrysoeriol mediates mitochondrial protection via PI3K/Akt pathway in MPP(+)-treated SH-SY5Y cells. Neurosci Lett. 2020;714:134545. 10.1016/j.neulet.2019.134545
24. Wu JY, Chen YJ, Fu XQ, Li JK, Chou JY, Yin CL, et al. Chrysoeriol suppresses hyperproliferation of rheumatoid arthritis fibroblast-like synoviocytes and inhibits JAK2/STAT3 signaling. BMC Compl Med Ther. 2022;22(1):73. 10.1186/s12906-022-03553-w
25. Min DY, Jung E, Ahn SS, Lee YH, Lim Y, Shin SY. Chrysoeriol prevents TNFα-induced CYP19 gene expression via EGR-1 downregulation in MCF7 breast cancer cells. Int J Mol Sci. 2020;21(20):7523. 10.3390/ijms21207523
26. Huang CY, Hu RC, Li J, Chen BB, Dai AG. α1-Antitrypsin alleviates inflammation and oxidative stress by suppressing autophagy in asthma. Cytokine. 2021;141:155454. 10.1016/j.cyto.2021.155454
27. Zhu L, Chen X, Chong L, Kong L, Wen S, Zhang H, et al. Adiponectin alleviates exacerbation of airway inflammation and oxidative stress in obesity-related asthma mice partly through AMPK signaling pathway. Int Immunopharmacol. 2019;67:396–407. 10.1016/j.intimp.2018.12.030
28. Wu SJ, Liou CJ, Chen YL, Cheng SC, Huang WC. Fucoxanthin ameliorates oxidative stress and airway inflammation in tracheal epithelial cells and asthmatic mice. Cells. 2021;10(6):1311. 10.3390/cells10061311
29. Raju KR, Kumar MN, Gupta S, Naga ST, Shankar JK, Murthy V, et al. 5-Aminosalicylic acid attenuates allergen-induced airway inflammation and oxidative stress in asthma. Pulm Pharmacol Ther. 2014;29(2):209–16. 10.1016/j.pupt.2014.07.007
30. Wu S, Li H, Yu L, Wang N, Li X, Chen W. IL-1β upregulates Muc5ac expression via NF-κB-induced HIF-1α in asthma. Immunol Lett. 2017;192:20–6. 10.1016/j.imlet.2017.10.006
31. Bai D, Sun T, Lu F, Shen Y, Zhang Y, Zhang B, et al. Eupatilin suppresses OVA-induced asthma by inhibiting NF-κB and MAPK and activating Nrf2 signaling pathways in mice. Int J Mol Sci. 2022;23(3):1582. 10.3390/ijms23031582
32. Li C, Deng C, Zhou T, Hu J, Dai B, Yi F, et al. MicroRNA-370 carried by M2 macrophage-derived exosomes alleviates asthma progression through inhibiting the FGF1/MAPK/STAT1 axis. Int J Biol Sci. 2021;17(7):1795–1807. 10.7150/ijbs.59715
33. Li C, Deng C, Zhou T, Hu J, Dai B, Yi F, et al. Importance of basophils in eosinophilic asthma: the murine counterpart. J Biol Regul Homeost Agents. 2018;32(2):335–9.
34. Poddighe D, Mathias CB, Freyschmidt EJ, Kombe D, Caplan B, Marseglia GL, et al. Basophils are rapidly mobilized following initial aeroallergen encounter in naïve mice and provide a priming source of IL-4 in adaptive immune responses. J Biol Regul Homeost Agents. 2014;28(1):91–103.