
Download
ORIGINAL ARTICLE
Galangin inhibits lipopolysaccharide-induced inflammation and stimulates osteogenic differentiation of bone marrow mesenchymal stem cells via regulation of AKT/mTOR signaling
Xiaoting Wanga, Xinglei Xiaob*
aDepartment of Trauma 2, The Sixth Affiliated Hospital of Xinjiang Medical University, Urumqi, China
bDepartment of Orthopedics, Jiangsu Huai’an Hospital of Traditional Chinese Medicine, Huai’an, China
Abstract
Background: Bone marrow mesenchymal stem cells (BMSCs), with the abilities of multidirectional differentiation and self-renewal, have been widely used in bone repair and regeneration of inflammation-stimulated oral diseases. Galangin is a flavonoid isolated from Alpinia officinarum, exerts anti-obesity, antitumor, and anti-inflammation pharmacological effects. The roles of galangin in lipopolysaccharide-induced inflammation and osteogenic differentiation of BMSCs were investigated.
Methods: BMSCs were isolated from rat bone marrow and identified by flow cytometry. The isolated BMSCs were treated with 1 μg/mL lipopolysaccharides or cotreated with lipopolysaccharides and different concentrations of galangin. Cell viability and apoptosis were detected by MTT (tetrazolium component) and flow cytometry. ELISA was used to detect inflammation. Alizarin red staining was used to investigate osteogenic differentiation.
Results: The rat BMSCs showed negative rate of CD34, and positive rate of CD29 and CD44. Lipopolysaccharides treatment reduced cell viability of BMSCs, and promoted the cell apoptosis. Incubation with galangin enhanced cell viability of lipopolysaccharide-stimulated BMSCs, and suppressed the cell apoptosis. Galangin decreased levels of TNF-α, IL-1β, and IL-6 in lipopolysaccharide-stimulated BMSCs through down-regulation of NF-κB phosphorylation (p-NF-κB). Galangin up-regulated expression of osteo-specific proteins, collagen type I alpha 1 (COL1A1), osteopontin (OPN), and runt-related transcription factor 2 (RUNX2), to promote the osteogenic differentiation of lipopolysaccharide-stimulated BMSCs. Protein expression of p-AKT and p-mTOR in lipopolysaccharide-stimulated BMSCs were increased by galangin treatment.
Conclusion: Galangin exerted an anti-inflammatory effect against lipopolysaccharide- stimulated BMSCs and promoted osteogenic differentiation through the activation of AKT/ mTOR signaling.
Key words: AKT/mTOR, bone marrow mesenchymal stem cells, galangin, inflammation, lipopolysaccharides, osteogenic differentiation
*Corresponding author: Xinglei Xiao, Department of Orthopedics, Jiangsu Huai’an Hospital of Traditional Chinese Medicine, No. 3 Heping Road, Qingjiangpu District, Huai’an, Jiangsu Province, China. Email address: [email protected]
Received 26 July 2022; Accepted 5 August 2022; Available online 1 January 2023
Copyright: Wang X, et al.
License: This open access article is licensed under Creative Commons Attribution 4.0 International (CC BY 4.0). http://creativecommons.org/licenses/by/4.0/
Introduction
Immuno-inflammatory oral diseases, including periodontitis, peri-implantitis, and periodontitis are caused by microbial infections and characterized by the destruction and irreversible loss of the alveolar bone and loosening and loss of teeth.1 Immuno-inflammatory oral diseases with high prevalence are generally treated with surgical or non-surgical management.2 Repair of the alveolar bone loss is considered a promising strategy for the treatment of immuno-inflammatory oral diseases.3 Bone marrow mesenchymal stem cells (BMSCs), with the abilities of multidirectional differentiation and self-renewal, have been widely used in periodontal regeneration.4
Oral bacteria, Porphyromonas gingivalis, with the ability to secrete virulence factors, such as lipopolysaccha-rides, have been considered an important risk factor in the immuno-inflammatory oral disease.5 Lipopolysaccharides have been shown to induce the secretion of osteoclastic cytokines, TNF-α, IL-1β, and receptor activator of nuclear factor κB ligand, in various cell types, including BMSCs.6 Moreover, lipopolysaccharides also impair osteoblast differentiation and suppress osteogenic differentiation tendency of BMSCs.7 Suppression of lipopolysaccharide-stimulated inflammation and differentiation in BMSCs contributed to the amelioration of immuno-inflammatory oral diseases.8
Galangin is a natural flavonoid extracted from Alpinia officinarum with pharmacological activities, such as anti-cancer, antibacterial, anti-inflammatory, hepatoprotective, and antiarthritic effects.9 For example, galangin inhibited cell proliferation and metastasis of osteosarcoma, and promoted cell apoptosis.10 Galangin induced differentiation of Th cells into immunosuppressive Tregs, and conferred tolerogenic property of dendritic cell in response to lipopolysaccharides.11 Galangin also inhibited lipopolysaccharide- stimulated microglial activation and suppressed the inflammatory response.12 In orthopedic research, galangin protected osteoblast against zoledronate-induced cell death,13 and suppressed osteoclast formation and inflammation in bone marrow-derived macrophages.14 Moreover, galangin promoted alkaline phosphatase activity to induce osteogenic differentiation in osteosarcoma cells.15 Therefore, it was hypothesized that galangin might inhibit inflammation of lipopolysaccharide-stimulated BMSCs, and promote the osteogenic differentiation. The effects of galangin on inflammation and osteogenic differentiation of lipopolysaccharide-stimulated BMSCs, as well as the related mechanism, were determined in this study.
Materials and Methods
Isolation and culture of BMSCs
Male Sprague–Dawley rats (4 weeks old and 120–150 g weight) were purchased from Slac Laboratory Animal Corp. (Shanghai, China). Rats were sacrificed using cervical dislocation, and the tibia and femur bones were carefully harvested from the rats. Eye scissors were used to intersect the terminals of bones, and the bone marrow was collected followed by washing with complete α-minimum essential medium (Invitrogen, Carlsbad, CA, USA). The cell suspensions were then seeded and cultured in the medium with 10% fetal bovine serum (Invitrogen) at 37°C in an incubator containing a 5% CO2 atmosphere. Twenty-four hours later, the nonadherent cells were removed and the isolated BMSCs were cultured in 10-cm cell culture dishes. Cells at passages 3–5 were used for the functional assays. All animal experiments were approved by the Ethics Committee of The Sixth Affiliated Hospital of Xinjiang Medical University for the use of animals and conducted in accordance with the National Institutes of Health Laboratory Animal Care and Use Guidelines.
BMSCs identification
The isolated BMSCs (passages 3–5) were harvested by incubation with 0.05% Trypsin (Sigma-Aldrich, St Louis, MO, USA). Cells were resuspended in a 100-μL phosphate- buffered saline solution (Beyotime, Beijing, China) and stained with fluorescent isothiocyanate (FITC)-anti-rat CD34 (Abcam, Cambridge, UK), phycoerythrin-anti-rat CD29 and CD44 (Abcam) antibodies. Cells were then analyzed by the FACS flow cytometer (Life Technologies, Darmstadt, Germany) for identifying BMSCs.
Cell viability and apoptosis assays
BMSCs were seeded in 96-wells plates and treated with 1 μg/mL of lipopolysaccharides (Sigma-Aldrich) or cotreated with lipopolysaccharides and different concentrations of galangin (5, 10, or 20 μM; Sigma-Aldrich) for 24 h. The plates were then added with a 5 mg/mL MTT solution (10 μL) (Beyotime) and incubated for another 4 h. Dimethyl sulfoxide was added into each well, and the absorbance at 450 nm was measured by microplate reader (Bio-Rad, Hercules, CA, USA). For cell apoptosis analysis, BMSCs post lipopolysaccharides or galangin treatment were harvested using trypsin and then resuspended in binding buffer of Annexin V FITC/propidium iodide (PI) staining kit (Invitrogen). Cells were stained with 5 µL of PI and 5 μL of FITC-annexin V (Invitrogen). The apoptotic ratio was calculated using the FACS flow cytometer (Life Technologies).
ELISA
The cultured medium of BMSCs were collected, and the BCA kit (Applygen, Beijing, China) was used to determine the protein concentration. Levels of TNF-α, IL-6, and IL-1β were determined via the ELISA kits (ExCell Biology, Inc., Shanghai, China).
Alizarin Red staining
BMSCs in osteogenic medium (90% DMEM, 10% fetal bovine serum, 0.25 mM ascorbate, 10 mM β-glycerophosphate, and 0.1 μM dexamethasone; HyClone, Logan, UT, USA) were seeded in 6-wells plates, and treated with 1 μg/mL lipopolysaccharides (Sigma-Aldrich) or cotreated with lipopolysaccharides and different concentrations of galangin (5, 10 or 20 μM; Sigma-Aldrich). The medium was changed every 3 days, and 2 weeks later, the cells were fixed in 4% paraformaldehyde and incubated with Alizarin Red (Sigma-Aldrich).
The BMSCs were photographed under microscope (Olympus, Tokyo, Japan), and the absorbance at 620 nm was measured by the microplate reader (Bio-Rad).
Western blotting
BMSCs were lysed in RIPA buffer (Beyotime), the isolated protein samples were separated by 10% SDS-PAGE, and then transferred onto nitrocellulose membranes. The membranes were blocked in 5% BCA, and then incubated with specific antibodies: anti-NF-κB and anti-p-NF-κB (1:1500, Abcam), anti-COL1A1 and anti-OPN (1:2000, Abcam), anti-RUNX2 and anti-β-actin (1:2500, Abcam), anti-AKT and anti-p-AKT (1:3000, Abcam), anti-mTOR and anti-p-mTOR (1:3500, Abcam) overnight. Following a wash with phosphate buffered saline, the membranes were then probed with horseradish peroxidase-conjugated secondary antibody (1:4000, Abcam). The immunoreactivities were visualized using enhanced chemiluminescence (Sigma-Aldrich) followed by incubation with tetramethylbenzidine.
Statistical analysis
All data with at least triplicates were expressed as mean ± SEM, and analyzed by the student’s t test or one-way analysis of variance under SPSS software. A p value <0.05 was considered as statistically significant.
Results
Galangin suppressed cell apoptosis of lipopolysaccharide-treated BMSCs
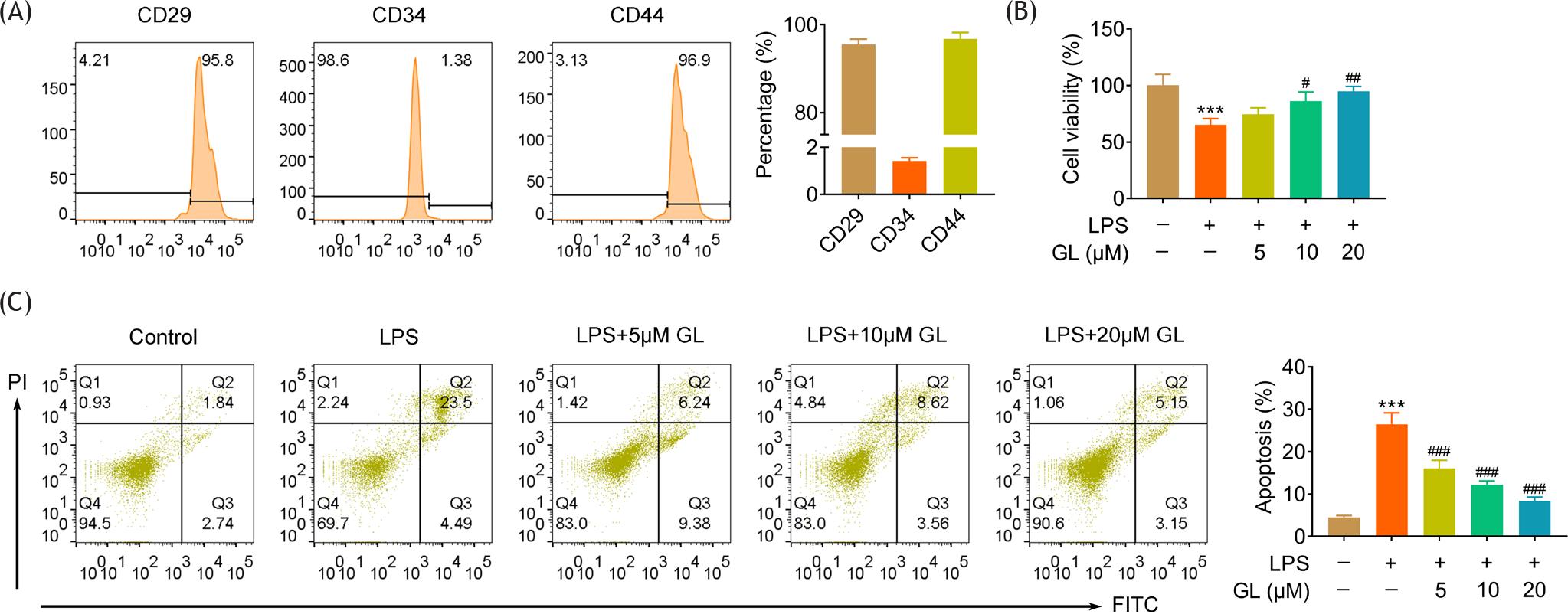
BMSCs were isolated from the rats, and flow cytometry showed that the isolated BMSCs had a negative rate of CD34 <2%, positive rate of CD29 and CD44 >95% (Figure 1A), indicating the high purity of BMSCs. BMSCs were treated with lipopolysaccharides, and the cell viability of BMSCs was decreased (Figure 1B), while the cell apoptosis was increased (Figure 1C) by lipopolysaccharides. However, incubation with galangin increased cell viability of lipopolysaccharide-treated BMSCs (Figure 1B) and reduced the cell apoptosis (Figure 1C) in a dosage-dependent manner, suggesting the anti-apoptotic effect of galangin against lipopolysaccharide-treated BMSCs.
Figure 1 Galangin suppressed cell apoptosis of lipopolysaccharide-treated BMSCs. (A) Flow cytometry showed negative CD34 and positive CD29 and CD44 expression in the isolated BMSCs. (B) Incubation with galangin increased cell viability of lipopolysaccharide-treated BMSCs in a dosage-dependent manner. (C) Incubation with galangin reduced the cell apoptosis of lipopolysaccharide-treated BMSCs in a dosage-dependent manner. # p < 0.05, ## p < 0.01, ###, *** p < 0.001.
Galangin suppressed lipopolysaccharide-induced inflammation in BMSCs
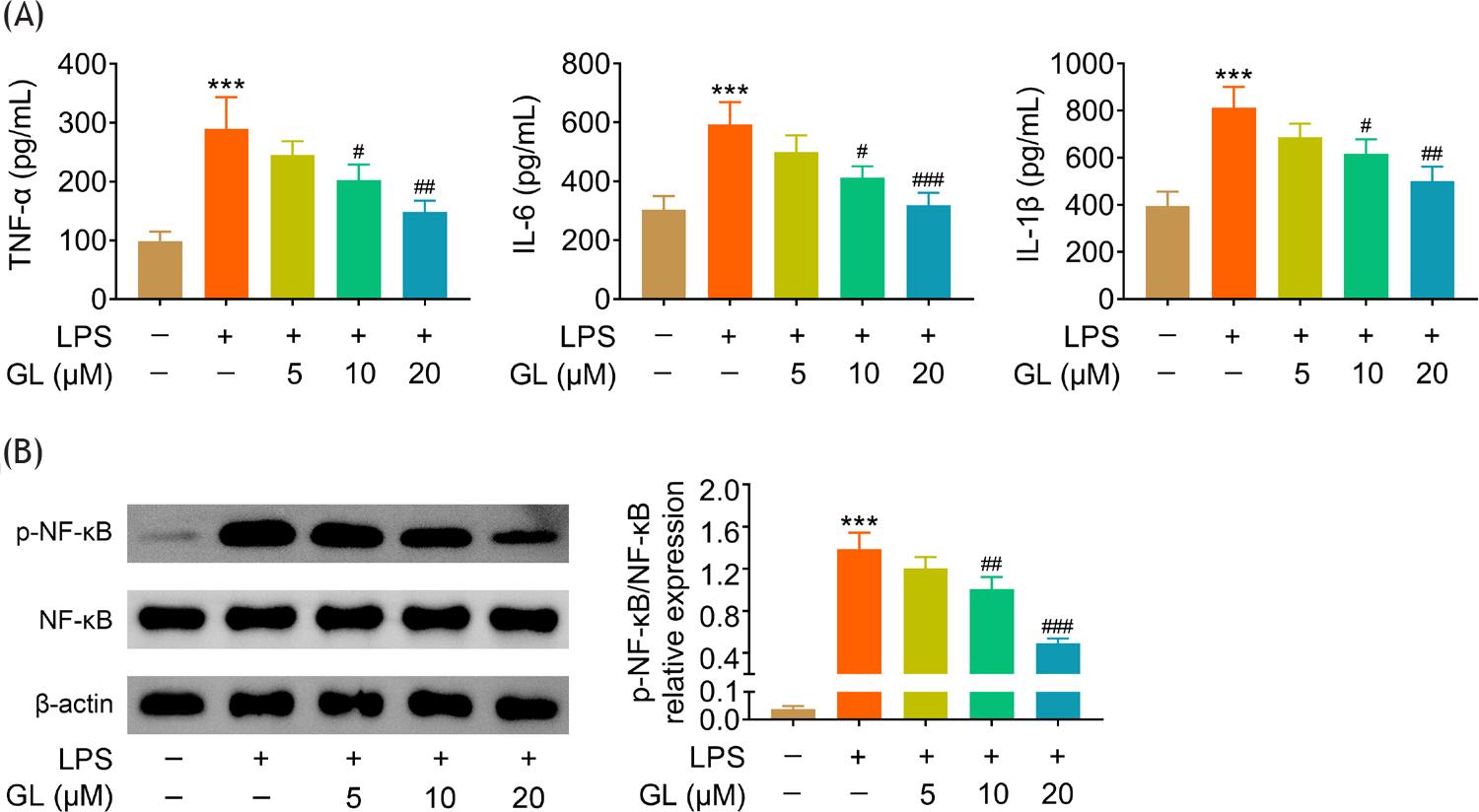
Lipopolysaccharides treatment induced up-regulation of TNF-α, IL-6, and IL-1β, thus promoting the inflammatory response in BMSCs (Figure 2A). However, galangin reduced the levels of TNF-α, IL-6, and IL-1β of lipopolysaccharide-treated BMSCs in a dosage dependent way (Figure 2A). Moreover, galangin attenuated lipopolysaccharide-induced increase of p-NF-κB in BMSCs (Figure 2B), suggesting that galangin suppressed activation of NF-κB signaling to reduce lipopolysaccharide-induced inflammation in BMSCs.
Figure 2 Galangin suppressed lipopolysaccharide-induced inflammation in BMSCs. (A) Incubation with galangin reduced TNF-α, IL-6, and IL-1β levels in lipopolysaccharide-treated BMSCs in a dosage dependent way. (B) Incubation with galangin attenuated lipopolysaccharide-induced increase of p-NF-κB in BMSCs in a dosage-dependent manner. # p < 0.05, ## p < 0.01, ###, *** p < 0.001.
Galangin promoted osteogenic differentiation of lipopolysaccharide-treated BMSCs
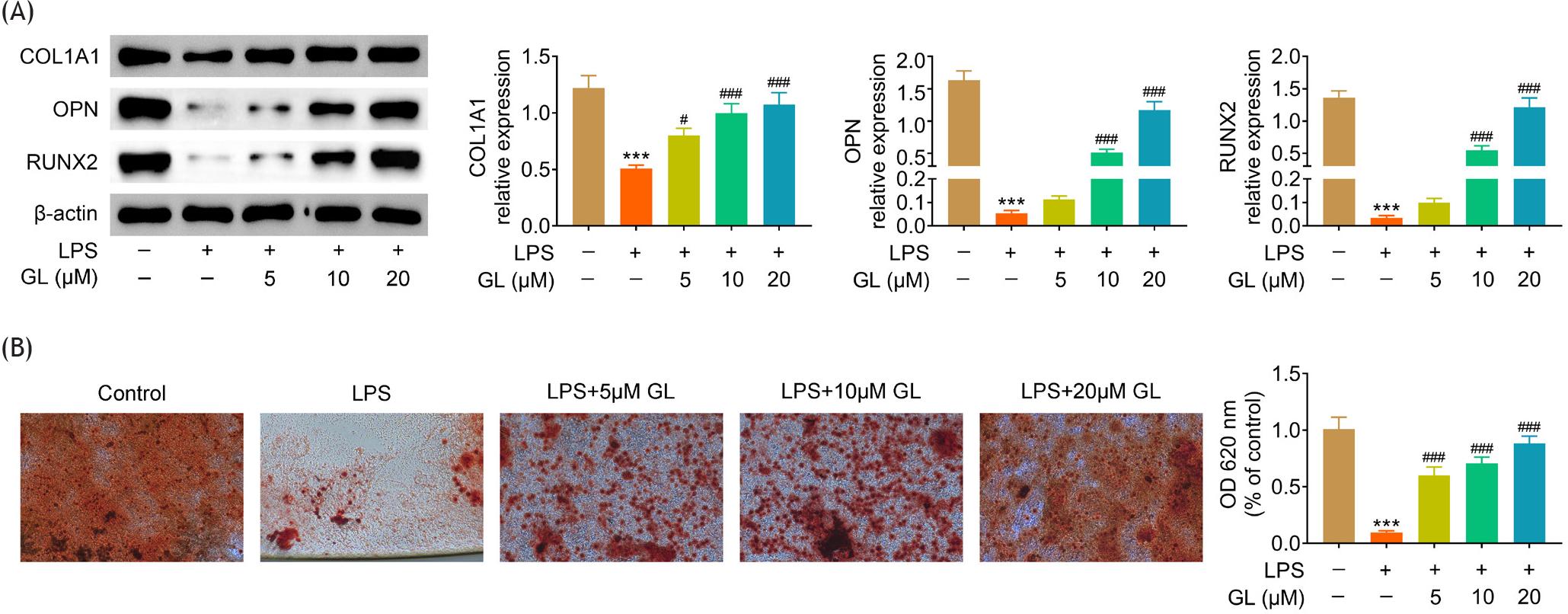
Osteoblast differentiation markers, COL1A1, OPN and RUNX2, in BMSCs, were reduced by lipopolysaccharides treatment (Figure 3A). Galangin enhanced expression of COL1A1, OPN and RUNX2 in lipopolysaccharide-treated BMSCs (Figure 3A) to stimulate the osteogenic differentiation. Alizarin Red staining demonstrated that the mineralization of BMSCs was reduced by lipopolysaccharides (Figure 3B), while galangin enhanced the mineralized nodule formation in lipopolysaccharide-treated BMSCs (Figure 3B), revealing that galangin protected BMSCs against lipopolysaccharides through promotion of osteogenic differentiation.
Figure 3 Galangin promoted osteogenic differentiation of lipopolysaccharide-treated BMSCs. (A) Incubation with galangin enhanced COL1A1, OPN, and RUNX2 expression in lipopolysaccharide-treated BMSCs in a dosage-dependent manner. (B) Alizarin Red staining demonstrated that incubation with galangin enhanced the mineralization of lipopolysaccharide-treated BMSCs in a dosage-dependent manner. # p < 0.05, ###, *** p < 0.001.
Galangin activated AKT/mTOR signaling in lipopolysaccharide-treated BMSCs
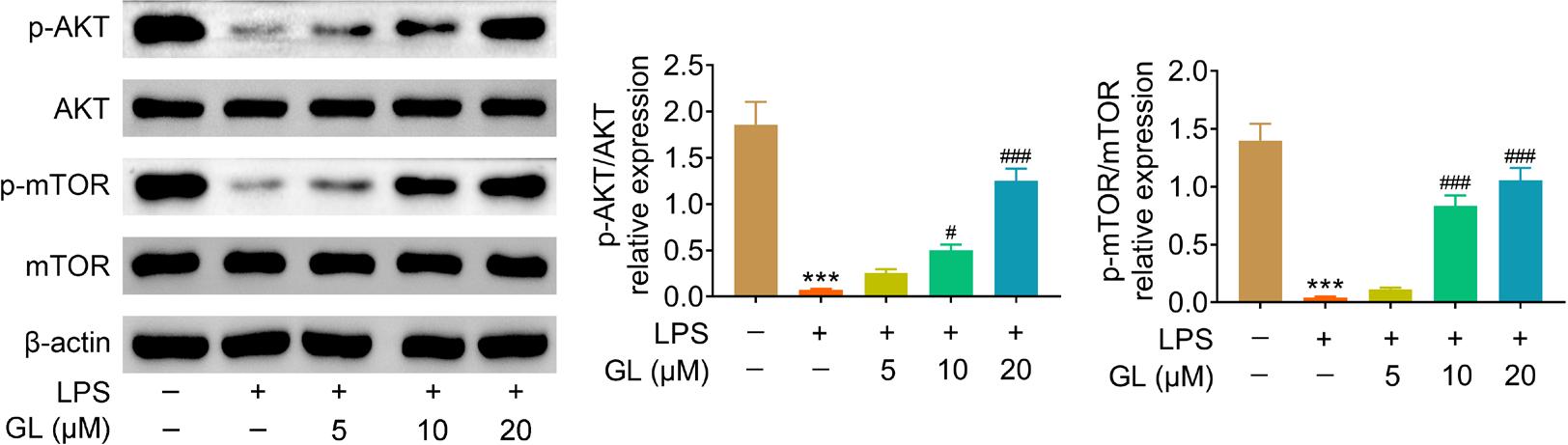
Protein expression of AKT and mTOR in BMSCs were not affected by incubation with lipopolysaccharides or galangin. (Figure 4). Phosphorylation of AKT and mTOR in BMSCs were down-regulated by lipopolysaccharides (Figure 4), whereas galangin up-regulated the phosphorylation of AKT and mTOR in lipopolysaccharide-treated BMSCs (Figure 4), indicating that AKT/mTOR signaling in lipopolysaccharide-treated BMSCs was activated by galangin treatment.
Figure 4 Galangin activated AKT/mTOR signaling in lipopolysaccharide-treated BMSCs. Incubation with galangin enhanced p-AKT and p-mTOR expression in lipopolysaccharide-treated BMSCs in a dosage-dependent manner. # p < 0.05, ###, *** p < 0.001.
Discussion
Traditional Chinese medicines have been shown to play a critical role in bone loss-related diseases.16 For example, extracts of Alpinia officinarum inhibited osteoclast differentiation and bone resorption, while enhanced osteoblast mineralization to prevent pathological bone diseases.17 Galangin, bioactive flavonoid isolated from Alpinia officinarum, suppressed osteoclast formation and inflammation and promoted osteogenic differentiation in various diseases.13–15 This study found that galangin exerted anti-inflammatory effect against lipopolysaccharide-treated BMSCs, and stimulated the osteogenic differentiation to prevent immuno-inflammatory oral diseases.
BMSCs, with the multidirectional differentiation potential and self-renewal ability, are significantly related to bone formation and resorption.18 BMSCs contribute to osseointegration through colonization of osteoblasts into the implant surface, promote alveolar bone regeneration, and facilitate for the treatment of immuno-inflammatory oral diseases.19 However, the chronic inflammatory micro-environment in immuno-inflammatory oral diseases influences the biological characteristics of stem cells, represses the repair and regeneration abilities.20 Cell viability of BMSCs was down-regulated in lipopolysaccharide-stimulated inflammatory environment, and the osteogenic differentiation was also suppressed.21 Therefore, suppressing inflammation of lipopolysaccharide-treated BMSCs might be a promising strategy for the prevention of immuno-inflammatory oral diseases. In this study, BMSCs, with the main characteristics of negative CD34 and positive CD29 and CD44 expression22 were isolated from rat bone marrow. Lipopolysaccharide treatment reduced cell viability of BMSCs, promoted cell apoptosis, and inhibited osteogenic differentiation through the down-regulation of COL1A1, OPN and RUNX2. A previous study reported that galangin promoted alkaline phosphatase activity, and increased expression of osteoblast differentiation markers (osteopontin, osteocalcin, COL1A1, alkaline phosphatase) to induce osteogenic differentiation in osteosarcoma cells.15 Here, galangin increased cell viability of lipopolysaccharide-treated BMSCs, suppressed the cell apoptosis, and promoted osteogenic differentiation through up-regulation of COL1A1, OPN, and RUNX2 in a dosage-dependent manner. Hence, galangin might be useful for treating immuno-inflammatory oral diseases.
Emerging evidence has shown that lipopolysaccha-rides induced the up-regulation of TNF-α, IL-1β, and IL-6 in BMSCs, contributed to the inflammatory responses involved in the pathogenesis of immuno-inflammatory oral diseases.23 Galangin suppressed lipopolysaccharide-stimulated inflammation through the down-regulation of TNF-α and IL-6.24 Results from this study demonstrated that galangin exerted an anti-inflammatory effect against lipopolysaccharide-treated BMSCs with decreased TNF-α, IL-1β, and IL-6 levels. NF-κB signaling, essential for the secretion of proinflammatory factors, was stimulated by lipopolysaccharides.25 The protective effect of galangin against lipopolysaccharide-induced acute inflammation in the lung was associated with NF-κB signaling inactivation.26 Here, the protein expression of p-NF-κB in lipopolysaccharide-treated BMSCs was reduced by galangin treatment in a dosage-dependent manner, suggesting that galangin protected BMSCs against lipopolysaccharide-induced inflammation through NF-κB signaling inactivation.
PI3K/AKT/mTOR signaling has been shown to be involved in synovial inflammation, subchondral bone dysfunction, and cartilage degradation, thus implicated in the pathogenesis of osteoarthritis.27 Activation of PI3K/AKT/mTOR signaling prevented bone loss,28 and promoted the osteogenic differentiation of BMSCs.29 Galangin promoted the activation of the AKT/mTOR pathway to ameliorate insulin resistance,30 suppress okadaic acid-induced cytotoxicity in neurons.31 Results from this study also indicated that galangin attenuated lipopolysaccharide-induced the down-regulation of p-AKT and p-mTOR in BMSCs in a dosage-dependent manner, thus promoting the AKT/mTOR signaling activation.
To sum up, galangin enhanced the cell viability of lipopolysaccharide-treated BMSCs and suppressed cell apoptosis. Moreover, galangin suppressed inflammation and promoted osteogenic differentiation of lipopolysaccharide-treated BMSCs through NF-κB signaling inactivation and AKT/mTOR signaling activation, respectively. However, the in vivo role of galangin in bone loss-related diseases should be investigated further.
Competing Interest
The authors declare no no conflicts of interest.
Ethics Approval
Ethical approval was obtained from the Ethics Committee of The Sixth Affiliated Hospital of Xinjiang Medical University.
Data Availability
The authors declare that all data supporting the findings of this study are available within the paper and any raw data can be obtained from the corresponding author upon request.
Author Contributions
Xiaoting Wang and Xinglei Xiao designed and conducted the experiments, analyzed and interpreted the data, and prepared the manuscript. All authors have read and approved the manuscript.
REFERENCES
1. Ramenzoni LL, Lehner MP, Kaufmann ME, Wiedemeier D, Attin T, Schmidlin PR. Oral diagnostic methods for the detection of periodontal disease. Diagnostics. 2021;11(3):571. 10.3390/diagnostics11030571
2. Shiba T, Watanabe T, Komatsu K, Koyanagi T, Nemoto T, Ohsugi Y, et al. Non-surgical treatment for periodontitis and peri-implantitis: longitudinal clinical and bacteriological findings-A case report with a 7-year follow-up evaluation. SAGE Open Med Case Rep. 2021;9:2050313X211029154–2050313X. 10.1177/2050313X211029154
3. Wennström JL, Tomasi C, Bertelle A, Dellasega E. Full-mouth ultrasonic debridement versus quadrant scaling and root planing as an initial approach in the treatment of chronic periodontitis. J Clin Periodontol. 2005;32(8):851–9. 10.1111/j.1600-051X.2005.00776.x
4. Liu L, Guo S, Shi W, Liu Q, Huo F, Wu Y, et al. Bone marrow mesenchymal stem cell-derived small extracellular vesicles promote periodontal regeneration. Tissue Eng Part A. 2020;27(13–14):962–76. 10.1089/ten.TEA.2020.0141
5. Wang L, Yang Y, Xiong X, Yu T, Wang X, Meng W, et al. Oral lichen-planus-associated fibroblasts acquire myofibroblast characteristics and secrete pro-inflammatory cytokines in response to Porphyromonas gingivalis lipopolysaccharide stimulation. BMC Oral Health. 2018;18(1):197. 10.1186/s12903-018-0656-6
6. Bandow K, Maeda A, Kakimoto K, Kusuyama J, Shamoto M, Ohnishi T, et al. Molecular mechanisms of the inhibitory effect of lipopolysaccharide (LPS) on osteoblast differentiation. Biochem Biophys Res Commun. 2010;402(4):755–61. 10.1016/j.bbrc.2010.10.103
7. Zhou R, Chen F, Liu H, Zhu X, Wen X, Yu F, et al. Berberine ameliorates the LPS-induced imbalance of osteogenic and adipogenic differentiation in rat bone marrow-derived mesenchymal stem cells. Mol Med Rep. 2021;23(5):350. 10.3892/mmr.2021.11989
8. Wang X, Jiang M, He X, Zhang B, Peng W, Guo L. N-acetyl cysteine inhibits the lipopolysaccharide-induced inflammatory response in bone marrow mesenchymal stem cells by suppressing the TXNIP/NLRP3/IL-1β signaling pathway. Mol Med Rep. 2020;22(4):3299–306. 10.3892/mmr.2020.11433
9. Basri AM, Taha H, Ahmad N. A review on the pharmacological activities and phytochemicals of alpinia officinarum (Galangal) extracts derived from bioassay-guided fractionation and isolation. Pharmacogn Rev. 2017;11(21):43–56. 10.4103/phrev.phrev_55_16
10. Yang Z, Li X, Han W, Lu X, Jin S, Yang W, et al. Galangin suppresses human osteosarcoma cells: an exploration of its underlying mechanism. Oncol Rep. 2017;37(1):435–41. 10.3892/or.2016.5224
11. Song H-Y, Kim WS, Han JM, Seo HS, Lim S-T, Byun E-B. Galangin treatment during dendritic cell differentiation confers tolerogenic properties in response to lipopolysaccharide stimulation. J Nutr Biochem. 2021;87:108524. 10.1016/j.jnutbio.2020.108524
12. Choi M, Lee E-J, Park J-S, Kim S-N, Park E-M, Kim H-S. Anti-inflammatory mechanism of galangin in lipopolysaccha-ride-stimulated microglia: Critical role of PPAR-γ signaling pathway. Biochem Pharmacol. 2017;144 :120–31. 10.1016/j.bcp.2017.07.021
13. Kim J-H, Kang H-M, Yu S-B, Song J-M, Kim C-H, Kim B-J, et al. Cytoprotective effect of flavonoid-induced autophagy on bisphosphonate mediated cell death in osteoblast. J Cell Biochem. 2018;119(7):5571–80. 10.1002/jcb.26728
14. Huh J-E, In-Tae J, Junyoung C, Yong-Hyeon B, Jae-Dong L, Dong-Suk P, et al. The natural flavonoid galangin inhibits osteoclastic bone destruction and osteoclastogenesis by suppressing NF-κB in collagen-induced arthritis and bone marrow-derived macrophages. Eur J Pharmacol. 2012; 698(1–3):57–66. 10.1016/j.ejphar.2012.08.013
15. Liu C, Ma M, Zhang J, Gui S, Zhang X, Xue S. Galangin inhibits human osteosarcoma cells growth by inducing transforming growth factor-β1-dependent osteogenic differentiation. Biomed Pharmacother. 2017;89:1415–21. 10.1016/j.biopha.2017.03.030
16. Zhu J. Influences of traditional Chinese medicine intervention on the bone growth and metabolism of rats with simulated weightlessness. Asian Pac J Trop Med. 2013;6(3):224–7. 10.1016/S1995-7645(13)60028-0
17. Ki-Shuk Shim C-JL, Nam-Hui Yim, Min Jung Gu, Jin Yeul Ma. Alpinia officinarum stimulates osteoblast mineralization and inhibits osteoclast differentiation. Am J Chin Med. 2016;44(6): 1255–71. 10.1142/S0192415X16500701
18. Arvidson K, Abdallah BM, Applegate LA, Baldini N, Cenni E, Gomez-Barrena E, et al. Bone regeneration and stem cells. J Cell Mol Med. 2011;15(4):718–746. 10.1111/j.1582-4934.2010.01224.x
19. Lu L, Liu Y, Zhang X, Lin J. The therapeutic role of bone marrow stem cell local injection in rat experimental periodontitis. J Oral Rehabil. 2020;47(S1):73–82. 10.1111/joor.12843
20. Hienz SA, Paliwal S, Ivanovski S. Mechanisms of bone resorption in periodontitis. J Immunol Res. 2015;2015:615486. 10.1155/2015/615486
21. Huang R-L, Yuan Y, Zou G-M, Liu G, Tu J, Li Q. LPS-stimulated inflammatory environment inhibits BMP-2-induced osteoblastic differentiation through crosstalk between TLR4/MyD88/NF0ºB and BMP/Smad signaling. Stem Cells Dev. 2014;23(3):277–89. 10.1089/scd.2013.0345
22. Pogozhykh O, Pogozhykh D, Neehus A-L, Hoffmann A, Blasczyk R, Müller T. Molecular and cellular characteristics of human and non-human primate multipotent stromal cells from the amnion and bone marrow during long term culture. Stem Cell Res Ther. 2015;6(1):150. 10.1186/s13287-015-0146-6
23. Zhang L, Deng S. Effects of astragaloside IV on inflammation and immunity in rats with experimental periodontitis. Braz Oral Res. 2019;33:e032. 10.1590/1807-3107bor-2019.vol33.0032
24. Lee HN, Shin SA, Choo GS, Kim HJ, Park YS, Kim BS, et al. Anti-inflammatory effect of quercetin and galangin in LPS-stimulated RAW264.7 macrophages and DNCB-induced atopic dermatitis animal models.Int J Mol Med. 2018; 41(2):888–98. 10.3892/ijmm.2017.3296
25. Čebatariūnienė A, Kriaučiūnaitė K, Prunskaitė J, Tunaitis V, Pivoriūnas A. Extracellular vesicles suppress basal and lipopolysaccharide-induced NFκB activity in human periodontal ligament stem cells. Stem Cells Dev. 2019;28(15):1037–49. 10.1089/scd.2019.0021
26. Shu Y-S, Tao W, Miao Q-B, Lu S-C, Zhu Y-B. Galangin dampens mice lipopolysaccharide-induced acute lung injury. Inflammation. 2014;37(5):1661–8. 10.1007/s10753-014-9894-1
27. Sun K, Luo J, Guo J, Yao X, Jing X, Guo F. The PI3K/AKT/ mTOR signaling pathway in osteoarthritis: a narrative review. Osteoarthr Cartil. 2020;28(4):400–9. 10.1016/j.joca.2020.02.027
28. Hiraiwa M, Ozaki K, Yamada T, Iezaki T, Park G, Fukasawa K, et al. mTORC1 activation in osteoclasts prevents bone loss in a mouse model of osteoporosis. Front Pharmacol. 2019;10:684. 10.3389/fphar.2019.00684
29. Zhao S-J, Kong F-Q, Jie J, Li Q, Liu H, Xu A-D, et al. Macrophage MSR1 promotes BMSC osteogenic differentiation and M2-like polarization by activating PI3K/AKT/GSK3β/β-catenin pathway. Theranostics. 2020;10(1):17–35. 10.7150/thno.36930
30. Liu Y, Liang X, Zhang G, Kong L, Peng W, Zhang H. Galangin and pinocembrin from propolis ameliorate insulin resistance in HepG2 cells via regulating Akt/mTOR signaling. Evid Based Complement Alternat Med. 2018;2018:7971842. 10.1155/2018/7971842
31. Huang L, Lin M, Zhong X, Yang H, Deng M. Galangin decreases p-tau, Aβ42 and β-secretase levels, and suppresses autophagy in okadaic acid-induced PC12 cells via an Akt/GSK3β/ mTOR signaling-dependent mechanism. Mol Med Rep. 2019;19(3):1767–74. 10.3892/mmr.2019.9824