
Download
REVIEW ARTICLE
Research progress on the role of Ndfip1 (Nedd4 family interacting protein 1) in immune cells
Ying-Le Tang, Lin Zhu, Hai-Fan Yang, Yan Tao, Wen Lu, Hong Cheng*
Medical College, Yangzhou University, Yangzhou, China
Abstract
Nedd4 family interacting protein 1 (Ndfip1) was first mentioned in an article in 2000. Since its discovery, related studies have shown that this protein is associated with apoptosis, neuroprotection, substance transport, ubiquitination, and immune regulation. It is noteworthy that the lack of Ndfip1 can lead to death in fetal mice. Researchers generally believe that the function of Ndfip1 is closely related to individual immune capacity and have published a large number of articles. However, a comprehensive classification of the immune regulatory function of Ndfip1 is still lacking. In this review, we will overview and discuss this new perspective, focusing on the role of Ndfip1 in the proliferation, differentiation, and cell activity of CD4+ T cells, CD8+ T cells, mast cells, and eosinophils. This review provides an updated summary of Ndfip1, which will unveil novel therapeutic targets. Finally, the conclusion is that Ndfip1 mainly plays a negative regulatory role in immune cells by maintaining the stability of the immune response and limiting its overexpression.
Key words: CD4+ T cells, cell differentiation, cell proliferation, immune regulation, Ndfip1
*Corresponding author: Hong Cheng, Medical College, Yangzhou University, Yangzhou, China. Email address: [email protected]
Received 25 July 2022; Accepted 23 October 2022; Available online: 1 January 2023
Copyright: Tang Y-L, et al.
License: This open access article is licensed under Creative Commons Attribution 4.0 International (CC BY 4.0). http://creativecommons.org/licenses/by/4.0/
Introduction
Ndfip1 is a protein that can bind to the ubiquitin E3 ligase Nedd4.1 Studies have reported that Ndfip1 can show a protective effect on nerve cells by mediating protein ubiquitination,2–4 and the protective mechanism may also be closely related to the migration of divalent metal ions.5–9 Since its immunological function has gradually attracted attention, many studies on the role of Ndfip1 in CD4+ T cells, CD8+ T cells, mast cells, and eosinophils have been published. To solve the lack of a comprehensive classification of the immune regulatory function of Ndfip1, we will summarize the main immune function performed by Ndfip1 and address its role on several immune cells in this paper.
Ndfip1 negatively regulates T-cell proliferation and differentiation
Ndfip1 inhibits the proliferation and differentiation of CD4+ T cells
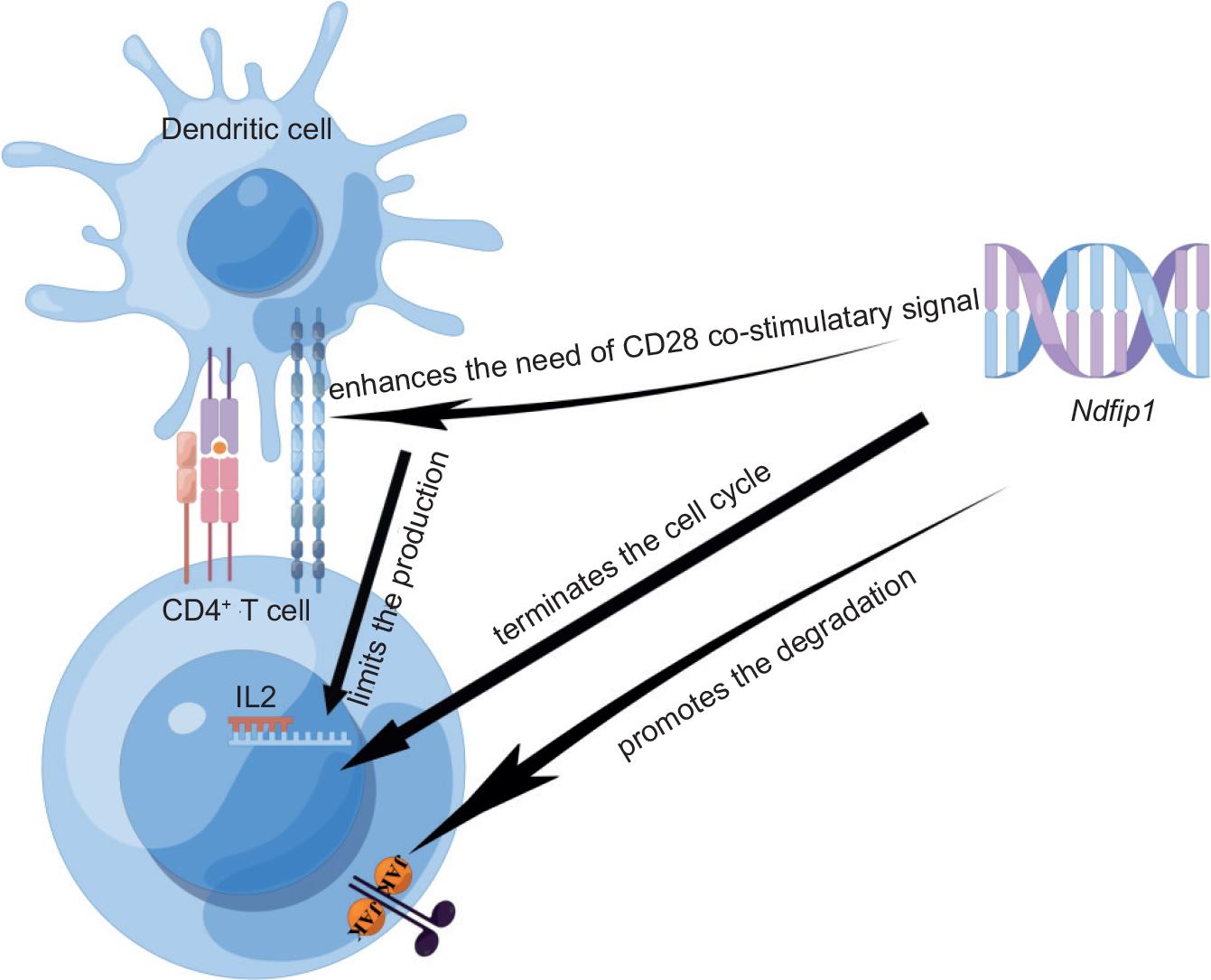
A study in 2006 found that CD4+ T cells were increased and activated in mice with Ndfip1 deficiency (Ndfip1 knockout); while mice lacking Ndfip1 developed spontaneous skin and lung inflammation and died prematurely.10 Later, Ramon et al.11 found that CD4+ T cells increased in the digestive tract of Ndfip1-deficient mice, especially in the esophagus. The reason why inflammation occurs in the known sites of environmental antigen exposure may be that the immune system of these mice responds to environmental antigens as though they are pathogenic. These results suggest that the function of Ndfip1 may be related to the regulation of tolerance to environmental antigens. Further, it was found that broad-spectrum antibiotic treatment drastically reduced the bacterial load in stool but hardly affected the activation of CD4+ T cells, gastrointestinal eosinophil infiltration, or esophageal epithelial cell hyperplasia, indicating that antibiotic treatment could not prevent the proliferation and differentiation of CD4+ T cells caused by Ndfip1 defects. In addition, this result also indicated that intestinal inflammation in NDFIP1-CKO mice (conditional knockout mice, mice lacking Ndfip1 only in T cells) was not caused by the loss of tolerance to intestinal microbiota but might be due to T-cell activation and eosinophilia caused by environmental antigens.12 To a certain extent, this confirmed the conclusion of Ramon et al.11 Further studies then reported that Ndfip1 inhibits IL-2 production by enhancing CD28 costimulation, while Ndfip1-deficient CD4+ T cells continuously produce IL-2 without CD28 costimulation, thus promoting the proliferation and differentiation of a large number of T cells.13 A year later, Altin et al.14 stated that Ndfip1 can prevent autoimmune diseases mainly by inducing the termination of the CD4+ T-cell cycle to prevent its proliferation and differentiation in their study. Afterwards, O’Leary et al.15 further demonstrated the mechanism that Ndfip1 promotes Jak1 degradation and regulates the cytokine signaling pathway to limit the proliferation of CD4+ T cells. In addition, the proliferation of Ndfip1-deficient CD4+ T cells was significantly higher than that of wild-type (WT) cells in the same cytokine environment, suggesting that Ndfip1-deficient T cells were more likely to be induced by cytokines promoting division and differentiation. At the same time, it was also found that Ndfip1-deficient CD4+ T cells were more likely to cause colitis. All of the above studies indicated that an abnormal CD4+ T-cell response occurred in Ndfip1 defects, which caused excessive proliferation, differentiation, and spontaneous aggregation of CD4+ T cells (Figure 1).13–15
Figure 1 The mechanism by which Ndfip1 inhibits the proliferation and differentiation of CD4+ T cells
Ndfip1 inhibits the proliferation and differentiation of CD4+ Th cells
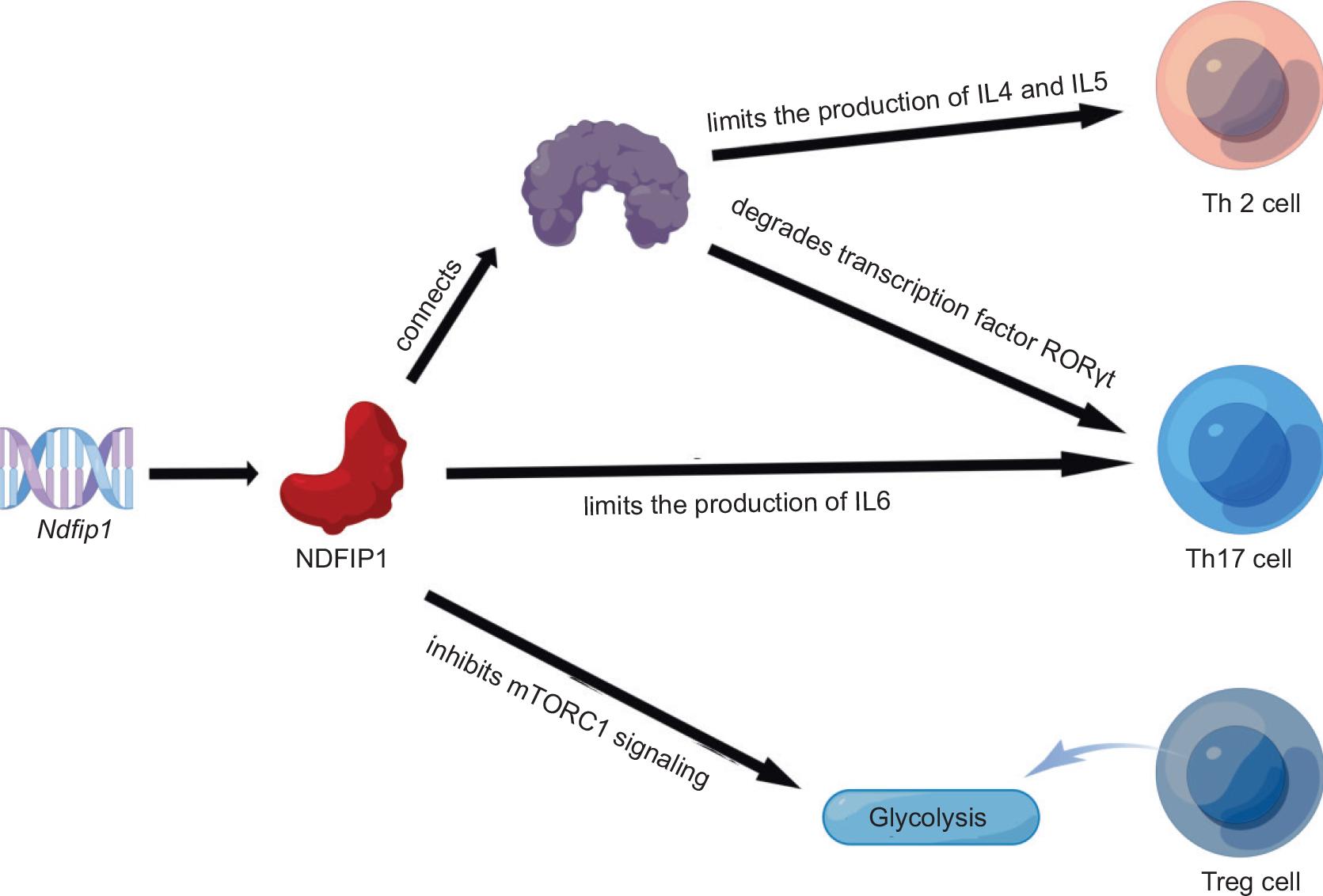
Ndfip1 inhibits the differentiation of CD4+ Th2 cells
Early literature suggested that inflammatory diseases of the skin, especially those involving eosinophils, are typically enhanced by Th2 CD4+ T cells.16,17 The hypothesis was that Ndfip1 deletion predispositions T cells to differentiate into Th2 cells and can cause Th2-mediated fatal inflammation.10,14 Following this, experiments show that Itch activity was not high and that Jun-B accumulated in the absence of Ndfip1. ITCH is a monomeric protein belonging to the homologous HECT-type ubiquitin ligase E3 family, which is involved in the ubiquitinated degradation of various proteins in vivo; in immune responses; in various cellular processes, including cell recycling, proliferation, and apoptosis; and plays an important role in the formation of tumors. ITCH cannot initiate the degradation of Jun-B in the absence of NDFIP1, which is a connector protein of Itch, and the accumulation of Jun-B leads to excessive production of IL-4, which promotes the differentiation of T cells into Th2 cells.10 Many subsequent studies revealed that Ndfip1 may be phosphorylated by T cells in a JNK-dependent pathway to accelerate the degradation of C-Jun and Jun-B, which is a transcription factor promoting IL-4 and IL-5 production.18–20 Therefore, several studies above showed that a prominent feature of Itch or Ndfip1 deficiency is increased differentiation of Th2 cells.19,21,,22
Ndfip1 inhibits the proliferation and differentiation of CD4+ Th17 cells
Two studies (including the references of Korn et al.23 and Louten et al.24) found that while Th17 cells play a protective role in the body, they also worsen the pathogenesis of inflammatory diseases, such as multiple sclerosis, arthritis, inflammatory bowel disease, and asthma. Later, Ramon et al.25 found that Ndfip1 inhibited the proliferation and differentiation of Th17 cells by blocking the production of proinflammatory cytokines (such as IL-6) in vivo. The results showed that the number of neutrophils and Th17 cells in the lungs of Ndfip1-deficient mice was increased, and the number of eosinophils in the spleen was increased and then induced by inflammatory conditions in secondary lymphoid organs and tissues, producing higher levels of IL-6 to promote Th17 differentiation. Th17 cells express the transcription factors RORγT4, RORα5, and STAT36 and can secrete IL-17F, IL-22, IFNγ, TNFα, and other cytokines.26 On this basis, further studies found that Ndfip1 could bind Itch to restrict Th17-cell proliferation by degrading the transcription factor RORγt and inhibiting the production of proinflammatory cytokines, thus preventing severe tissue inflammation and inflammatory bowel disease.27
Ndfip1 inhibits the proliferation and differentiation of CD4+ Treg cells
It was found that the expression of Ndfip1 was temporarily upregulated in a TGF-β-dependent manner during the differentiation of iTreg (induced regulatory T) cells, and TGF-β induced the expression of Ndfip1 to inhibit the production of IL-4, thereby preventing iTreg cell differentiation.28 Following this several studies reported that the mechanistic target of rapamycin (mTOR) is a serine-threonine kinase, which constitutes a part of the protein complexes of mTORC1 and mTORC2. Since appropriate regulation of cellular energetics and metabolism is important to the function and lineage stability of Treg cells, these two complexes, as key regulators of cell metabolism, can limit glycolysis in Treg cells by promoting lineage stability.29–31 Moreover, mTORC1/mTORC2 deficiency impaired CD4+ T-cell accumulation and immunoglobulin A production.32 In 2017, the study of Layman et al.33 supported the inherent role of Ndfip1 in limiting IL-4 production in Treg cells but also showed that Ndfip1-deficient Treg cells were more likely to produce IL-10, IFNγ, and IL-17A wild-type controls. Furthermore, they explored and proved that Ndfip1 could prevent high proliferation and active metabolism of Treg cells by inhibiting mTORC1 signaling of glycolysis of Treg cells, maintain the stability of Treg lineage and immune homeostasis, and prevent spontaneous inflammation in some body parts, such as lungs and skin. As mentioned in the article, Ndfip1 limits both the accumulation and proliferation of Treg cells and prevents the production of IL-4 by Treg cells, and Treg cells lacking Ndfip1 show increased conserved noncoding DNA sequence 2 (CNS2) methylation and are prone to losing Foxp3 expression. Based on the above research, we speculate that Ndfip1 promotes the generation of mTORC1/mTORC2 protein complexes and therefore increases the lineage stability of Treg cells by inhibiting mTORC1 glycolysis signaling (Figure 2).10,25,27,28,32
Figure 2 The mechanism by which Ndfip1 inhibits the proliferation and differentiation of CD4+ Th cells and CD4+ Treg cells.
Ndfip1 inhibits the proliferation and differentiation of CD8+ T cells under high tolerance antigen concentrations
In 2018, Wagle et al.34 confirmed that Ndfip1 can act as a key checkpoint for inhibiting the proliferation and differentiation of CD8+ T cells in an environment with high tolerance antigen levels and can autonomously prevent the aggregational destruction of tissue cells targeted by CD8+ T cells and prevent the occurrence of autoimmune diabetes. In contrast, Ndfip1 hardly affected the proliferation and differentiation of CD8+ T cells during acute infection or at low tolerance antigen concentrations. Furthermore, the peripheral tolerance checkpoints of CD8+ T cells are usually mediated by cytolytic protease granzyme B (GzmB), and the production of cytokines inhibits the differentiation of effector cells.35,36 Therefore, the increased expression of GzmB in Ndfip1-deficient CD8+ T cells indicates that Ndfip1 can inhibit the type 1 immune response, while the deletion of Ndfip1 in CD4+ T cells has almost no effect on Th1 differentiation.10,27,34
Ndfip1 can inhibit mast cell activity
The study of Yip et al.37 revealed that Ndfip1 limited the intensity and duration of positive signal transduction induced by IgE-FcɛRI in mast cells through ubiquitination of p-Syk, thereby controlling the release of mast cell media and limiting in vivo overreaction and pathological changes. Syk is an indispensable tyrosine kinase for the activity of the downstream FcɛRI signaling body. After phosphorylated Syk is ubiquitinated, the loss of its tyrosine kinase activity leads to a reduction in calcium degranulation and cytokine production.38 Conversely, loss of Nedd4-2 or Ndfip1 in mast cells leads to prolonged phosphorylation of Syk and its substrates, increased histamine release, and significantly increased production of the proinflammatory cytokines TNF, IL-6, and CCL2, resulting in intensification and prolongation of IgE-mediated skin allergic reactions .39,40
Defects in Ndfip1 can increase eosinophil cell activity
Oliver et al.10 found that Ndfip1 knockout mice were prone to develop an inflammatory state with a large number of infiltrating eosinophils. Ramon et al.11 found that the infiltrating cells in the esophagus, stomach, cecum, and colon of Ndfip1-deficient mice were characterized by eosinophils. In addition, it was confirmed that a large amount of IL-5 produced by over-proliferating CD4+ T cells promoted the recruitment of gastrointestinal eosinophils in Ndfip1-deficient mice. Gastrointestinal diseases are caused by the recruitment of eosinophils in Ndfip1-deficient mice, such as eosinophilic esophagitis and eosinophilic gastrointestinal disease or food allergy. As the activation of eosinophils is generally considered a byproduct of excess cytokines produced by the proliferation of CD4+ T cells due to the loss of Ndfip1, there is no much literature specifically available that study the effect of Ndfip1 on eosinophils.
Future perspective
Currently, Ndfip1 has been found to protect against autoimmune diseases, antiviral infections, anti-tumor (such as hepatocellular carcinoma,41 uveal melanoma,42 pancreatic ductal adenocarcinoma,43 head and neck squamous cell carcinoma,44,45 glioblastoma multiforme,46 breast cancer,47 etc.) and has a neuroprotective effect in neurodegenerative diseases and traumatic brain injury. However, the impact of Ndfip1 on numerous functions of infection and tumors remains to be explored.
Moreover, as seen in Table 1, related studies on immune cells are not in-depth enough. Strengthening the study of other immune cells will more widely contribute to a more comprehensive understanding of the immune function of Ndfip1 and may lead to the development of novel Ndfip1-based therapeutic research axes in the future.
Table 1 Different immune cells and methods of validation.
| Immune cell | Main validation methods | Main validation model | Species | Reference | ||
|---|---|---|---|---|---|---|
| T-cell | CD4+ T cell | WB, PCR, FCM, IP, MBMCE | Ubi-GFP mice, C57BL/6 mice | mice | 10 | |
| IHC, ELISA, FCM, MBMCE | Ndfip1−/− mice, Rag1−/− mice, Itch mutant mice, C57BL/6 mice | mice | 11 | |||
| PCR, PCR, FCM, IHC | Ndfip1-cKO mice | mice | 12 | |||
| WB, PCR, FCM, ELISA, IHC | Ndfip1−/− mice, Itch mutant mice, CD45.1 mice, IL-4 mice, CD28 mice, OT-II mice, Rag1 mice, CD4-Cre mice | mice | 13 | |||
| WB, FCM, MBMCE | 3A9 TCR Tg mice, insHEL Tg mice, Aire−/− mice, Bim−/− mice, Foxp3−/− mice | mice | 14 | |||
| GST pulldown, PUA, UCA, FLC, IGD, K-ɛgg IP, LC-MS/MS, WB, PCR, FCM, IHC | Ndfip1−/− mice, Ndfip1fl/fl CD4-Cre mice, IL-4−/− mice, Ndfip1−/− IL-4−/− (DKO) mice, IL-17A GFP mice, Tcell B3K506 mice | mice | 15 | |||
| CD4+ Th cell | Th2 cell | WB, PCR, FCM, IP, MBMCE | Ubi-GFP mice, C57BL/6 mice | mice | 10 | |
| WB, FCM, MBMCE | 3A9 TCR Tg mice, insHEL Tg mice, Aire−/− mice, Bim−/− mice, Foxp3−/− mice | mice | 20 | |||
| Th 17 cell | ATTE, WB, PCR, ELISA, FCM, IHC, MBMCE | C57BL/6 mice, CD45.1 mice, CD4-Cre mice, IL-17A GFP mice, Tcell B3K506 mice | mice | 17 | ||
| WB, FCM, ELISA | Ndfip1−/− mice, IL-4−/− mice | mice | 27 | |||
| Tregcell | WB, PCR, ELISA, FCM, IHC | CD45.1 mice, IL-4−/− mice, OT-II mice, Rag1−/− mice, Itch mutant mice, Ndfip1fl/fl CD4-Cre mice | mice | 29 | ||
| BS, WB, PCR, ELISA, FCM, IHC, MBMCE, MS | CD45.1 mice, Foxp3-Cre mice, Ndfip1fl/fl mice | mice | 34 | |||
| CD8+ T cell | FCM, IHC, MBMCE, HPAE | Rag1−/− Ndfip1kru/kru OT-I mice, Ndfip1kru/krumice, Ndfip1f/f (floxed Ndfip1 gene) mice, GzmB-cre mice, C57BL/6 mice, OT-I mice, B6.129S7-Rag1tm1Mom/J (Rag1−/−) mice, B6.SJL-PfpreaPep3b/BoyJ (CD45.1) mice, RIP-OVAhl mice | mice | 16 | ||
| Mast cell | EIA, WB, PCR, ELISA, IP, FCM | Mcl-1fl/fl mice, Mcl-1+/+ mice, C57BL/6 mice, Kit mice, Cpa3-cre mice, Nedd4-2−/− mice, Ndfip1−/− mice, C57BL/6 mice | mice | 37 | ||
| Eosi-nophil cell | WB, PCR, FCM, IP, MBMCE | Ubi-GFP mice, C57BL/6 mice | mice | 10 | ||
| FCM, IHC, ELISA, MBMCE | Ndfip1−/− mice, Rag1−/− mice, Itch mutant mice, C57BL/6 mice | mice | 11 | |||
WB: western blot; PCR: polymerase chain reaction; FCM: flow cytometric analysis; IP: immunoprecipitation; MBMCE: mixed bone marrow chimera experiments; IHC: immunohistochemistry; ELISA: enzyme-linked immunosorbent assay; GST pulldown: glutathione-S-transferase pulldown; PUA: poly ubiquitination assay; UCA: ubiquitin charging assay; FLC: fetal liver chimeras; IGD: In-gel digest; K-ɛgg IP: K-ɛ-GG peptide immunoprecipitation; LC-MS/MS: liquid chromatograph-mass spectrometry/mass spectrometry; ATTE: adoptive Th17 transfer experiments; BS: bisulfite sequencing; EIA: enzyme immunoassay.
Conclusions
As reviewed above, recent studies indicate that Ndfip1 is an important player in immune regulation and negatively regulates the functions of immune cells. In the case of Ndfip1 deficiency, excessive proliferation and differentiation of several immune cells will produce a large number of cytokines, leading to autoimmune diseases. Therefore, the role of Ndfip1 is to maintain the stability of the immune response and limit its overexpression.
Conflicts of interest
The authors declare no potential conflicts of interest concerning the research, authorship, and/or publication of this article.
Acknowledgments
The figures in this review were created with fig draw.
Funding
The project is supported by National Natural Science Foundation of China (31071216), Natural Science Foundation of Jiangsu Province (BK2010306) and Jiangsu Students’ innovation training program (202211117079Y).
REFERENCES
1. Jolliffe CN, Harvey KF, Haines BP, Parasivam G, Kumar S. Identification of multiple proteins expressed in murine embryos as binding partners for the WW domains of the ubiquitin-protein ligase Nedd4. Biochem J. 2000;351(Pt 3):557–65. 10.1042/bj3510557
2. Lackovic J, Howitt J, Callaway JK, Silke J, Bartlett P, Tan SS. Differential regulation of Nedd4 ubiquitin ligases and their adaptor protein Ndfip1 in a rat model of ischemic stroke. Exp Neurol. 2012;235(1):326–35. 10.1016/j.expneurol.2012.02.014
3. Howitt J, Gysbers AM, Ayton S, Carew-Jones F, Putz U, Finkelstein DI, et al. Increased Ndfip1 in the substantia nigra of Parkinsonian brains is associated with elevated iron levels. PLoS One. 2014;9(1):e87119. 10.1371/journal.pone.0087119
4. Sang Q, Kim MH, Kumar S, Bye N, Morganti-Kossman MC, Gunnersen J, et al. Nedd4-WW domain-binding protein 5 (Ndfip1) is associated with neuronal survival after acute cortical brain injury. J Neurosci. 2006;26(27):7234–44. 10.1523/JNEUROSCI.1398-06.2006
5. Howitt J, Putz U, Lackovic J, Doan A, Dorstyn L, Cheng H, et al. Divalent metal transporter 1 (DMT1) regulation by Ndfip1 prevents metal toxicity in human neurons. Proc Natl Acad Sci U S A. 2009;106(36):15489–94. 10.1073/pnas.0904880106
6. Foot NJ, Leong YA, Dorstyn LE, Dalton HE, Ho K, Zhao L, et al. Ndfip1-deficient mice have impaired DMT1 regulation and iron homeostasis. Blood. 2011;117(2):638–46. 10.1182/blood-2010-07-295287
7. Xu H, Chang Q, Jia W, Jiang H, Sun P, Xie J. Iron status and lipopolysaccharide regulate Ndfip1 by activation of nuclear factor-kappa B. Biometals. 2013;26(6):981–8. 10.1007/s10534-013-9674-7
8. Traeger L, Wiegand SB, Sauer AJ, Corman BHP, Peneyra KM, Wunderer F, et al. UBA6 and NDFIP1 regulate the degradation of ferroportin. Haematologica. 2022;107(2):478–88. 10.3324/haematol.2021.278530
9. Garrick MD, Zhao L, Roth JA, Jiang H, Feng J, Foot NJ, et al. Isoform specific regulation of divalent metal (ion) transporter (DMT1) by proteasomal degradation. Biometals. 2012;25(4):787–93. 10.1007/s10534-012-9522-1
10. Oliver PM, Cao X, Worthen GS, Shi P, Briones N, MacLeod M, et al. Ndfip1 protein promotes the function of itch ubiquitin ligase to prevent T cell activation and T helper 2 cell-mediated inflammation. Immunity. 2006;25(6):929–40. 10.1016/j.immuni.2006.10.012
11. Ramon HE, Riling CR, Bradfield J, Yang B, Hakonarson H, Oliver PM. The ubiquitin ligase adaptor Ndfip1 regulates T cell-mediated gastrointestinal inflammation and inflammatory bowel disease susceptibility. Mucosal Immunol. 2011;4(3):314–24. 10.1038/mi.2010.69
12. Kurzweil V, Tarangelo A, Oliver PM. Gastrointestinal microbiota do not significantly contribute to T cell activation or GI inflammation in Ndfip1-cKO mice. PLoS One. 2012;7(4):e34478. 10.1371/journal.pone.0034478
13. Ramos-Hernández N, Ramon HE, Beal AM, Laroche A, Dekleva EA, Oliver PM. Ndfip1 enforces a requirement for CD28 costimulation by limiting IL-2 production. J Immunol. 2013;191(4):1536–46. 10.4049/jimmunol.1203571
14. Altin JA, Daley SR, Howitt J, Rickards HJ, Batkin AK, Horikawa K, et al. Ndfip1 mediates peripheral tolerance to self and exogenous antigen by inducing cell cycle exit in responding CD4+ T cells. Proc Natl Acad Sci U S A. 2014;111(6):2067–74. 10.1073/pnas.1322739111
15. O’Leary CE, Riling CR, Spruce LA, Ding H, Kumar S, Deng G, et al. Ndfip-mediated degradation of Jak1 tunes cytokine signalling to limit expansion of CD4+ effector T cells. Nat Commun. 2016;7:11226. 10.1038/ncomms11226
16. Del Prete G. Human Th1 and Th2 lymphocytes: Their role in the pathophysiology of atopy. Allergy. 1992;47(5):450–5. 10.1111/j.1398-9995.1992.tb00662.x
17. Romagnani S, Maggi E, Parronchi P, Macchia D, Piccinni MP, Ricci M. Increased numbers of Th2-like CD4+ T cells in target organs and in the allergen-specific repertoire of allergic patients. Possible role of IL-4 produced by non-T cells. Int Arch Allergy Appl Immunol. 1991;94(1–4):133–6. 10.1159/000235344
18. Gao M, Labuda T, Xia Y, Gallagher E, Fang D, Liu YC, et al. Jun turnover is controlled through JNK-dependent phosphorylation of the E3 ligase Itch. Science. 2004;306(5694):271–5. 10.1126/science.1099414
19. Fang D, Elly C, Gao B, Fang N, Altman Y, Joazeiro C, et al. Dysregulation of T lymphocyte function in itchy mice: A role for Itch in TH2 differentiation. Nat Immunol. 2002;3(3):281–7. 10.1038/ni763
20. Hartenstein B, Teurich S, Hess J, Schenkel J, Schorpp-Kistner M, Angel P. Th2 cell-specific cytokine expression and allergen-induced airway inflammation depend on JunB. EMBO J. 2002;21(23):6321–9. 10.1093/emboj/cdf648
21. Li B, Tournier C, Davis RJ, Flavell RA. Regulation of IL-4 expression by the transcription factor JunB during T helper cell differentiation. EMBO J. 1999;18(2):420–32. 10.1093/emboj/18.2.420
22. Perry WL, Hustad CM, Swing DA, O’Sullivan TN, Jenkins NA, Copeland NG. The itchy locus encodes a novel ubiquitin protein ligase that is disrupted in a18H mice. Nat Genet. 1998;18(2):143–6. 10.1038/ng0298-143
23. Korn T, Bettelli E, Oukka M, Kuchroo VK. IL-17 and Th17 Cells. Annu Rev Immunol. 2009;27:485–517. 10.1146/annurev.immunol.021908.132710
24. Louten J, Boniface K, de Waal Malefyt R. Development and function of TH17 cells in health and disease. J Allergy Clin Immunol. 2009;123(5):1004–11. 10.1016/j.jaci.2009.04.003
25. Ramon HE, Beal AM, Liu Y, Worthen GS, Oliver PM. The E3 ubiquitin ligase adaptor Ndfip1 regulates Th17 differentiation by limiting the production of proinflammatory cytokines. J Immunol. 2012;188(8):4023–31. 10.4049/jimmunol.1102779
26. Yang XO, Panopoulos AD, Nurieva R, Chang SH, Wang D, Watowich SS, et al. STAT3 regulates cytokine-mediated generation of inflammatory helper T cells. J Biol Chem. 2007;282(13):9358–63. 10.1074/jbc.C600321200
27. Layman AAK, Sprout SL, Phillips D, Oliver PM. Ndfip1 restricts Th17 cell potency by limiting lineage stability and proinflammatory cytokine production. Sci Rep. 2017;7:39649. 10.1038/srep39649
28. Beal AM, Ramos-Hernández N, Riling CR, Nowelsky EA, Oliver PM. TGF-β induces the expression of the adaptor Ndfip1 to silence IL-4 production during iTreg cell differentiation. Nat Immunol. 2011;13(1):77–85. 10.1038/ni.2154
29. Newton R, Priyadharshini B, Turka LA. Immunometabolism of regulatory T cells. Nat Immunol. 2016;17(6):618–25. 10.1038/ni.3466
30. Apostolidis SA, Rodríguez-Rodríguez N, Suárez-Fueyo A, Dioufa N, Ozcan E, Crispín JC, et al. Phosphatase PP2A is requisite for the function of regulatory T cells. Nat Immunol. 2016;17(5):556–64. 10.1038/ni.3390
31. Wei J, Long L, Yang K, Guy C, Shrestha S, Chen Z, et al. Autophagy enforces functional integrity of regulatory T cells by coupling environmental cues and metabolic homeostasis. Nat Immunol. 2016;17(3):277–85. 10.1038/ni.3365
32. Zeng H, Cohen S, Guy C, Shrestha S, Neale G, Brown SA, et al. mTORC1 and mTORC2 Kinase signaling and glucose metabolism drive follicular helper T cell differentiation. Immunity. 2016;45(3):540–54. 10.1016/j.immuni.2016.08.017
33. Layman AAK, Deng G, O’Leary CE, Tadros S, Thomas RM, Dybas JM, et al. Ndfip1 restricts mTORC1 signalling and glycolysis in regulatory T cells to prevent autoinflammatory disease. Nat Commun. 2017;8:15677. 10.1038/ncomms15677
34. Wagle MV, Marchingo JM, Howitt J, Tan SS, Goodnow CC, Parish IA. The ubiquitin ligase adaptor NDFIP1 selectively enforces a CD8+ T cell tolerance checkpoint to high-dose antigen. Cell Rep. 2018;24(3):577–84. 10.1016/j.celrep.2018.06.060
35. Hernandez J, Aung S, Redmond WL, Sherman LA. Phenotypic and functional analysis of CD8(+) T cells undergoing peripheral deletion in response to cross-presentation of self-antigen. J Exp Med. 2001;194(6):707–17. 10.1084/jem.194.6.707
36. Parish IA, Rao S, Smyth GK, Juelich T, Denyer GS, Davey GM, et al. The molecular signature of CD8+ T cells undergoing deletional tolerance. Blood. 2009;113(19):4575–85. 10.1182/blood-2008-10-185223
37. Yip KH, Kolesnikoff N, Hauschild N, Biggs L, Lopez AF, Galli SJ, et al. The Nedd4-2/Ndfip1 axis is a negative regulator of IgE-mediated mast cell activation. Nat Commun. 2016;7:13198. 10.1038/ncomms13198
38. Costello PS, Turner M, Walters AE, Cunningham CN, Bauer PH, Downward J, et al. Critical role for the tyrosine kinase Syk in signalling through the high affinity IgE receptor of mast cells. Oncogene. 1996;13(12):2595–605. PMid: 9000133
39. Zhang J, Chiang YJ, Hodes RJ, Siraganian RP. Inactivation of c-Cbl or Cbl-b differentially affects signaling from the high affinity IgE receptor. J Immunol. 2004;173(3):1811–18. 10.4049/jimmunol.173.3.1811
40. Gustin SE, Thien CB, Langdon WY. Cbl-b is a negative regulator of inflammatory cytokines produced by IgE-activated mast cells. J Immunol. 2006;177(9):5980–9. 10.4049/jimmunol.177.9.5980
41. Zhang Y, Zhang C, Zhao Q, Wei W, Dong Z, Shao L, et al. The miR-873/NDFIP1 axis promotes hepatocellular carcinoma growth and metastasis through the AKT/mTOR-mediated Warburg effect. Am J Cancer Res. 2019;9(5):927–44. PMid: 31218102.
42. Peng J, Liu H, Liu C. MiR-155 promotes uveal melanoma cell proliferation and invasion by regulating NDFIP1 expression. Technol Cancer Res Treat. 2017;16(6):1160–7. 10.1177/1533034617737923
43. Ben Q, Sun Y, Liu J, Wang W, Zou D, Yuan Y. Nicotine promotes tumor progression and epithelial-mesenchymal transition by regulating the miR-155-5p/NDFIP1 axis in pancreatic ductal adenocarcinoma. Pancreatology. 2020;20(4):698–708. 10.1016/j.pan.2020.04.004
44. Routila J, Suvila K, Grénman R, Leivo I, Westermarck J, Ventelä S. Cancer cell line microarray as a novel screening method for identification of radioresistance biomarkers in head and neck squamous cell carcinoma. BMC Cancer. 2021;21(1):868. 10.1186/s12885-021-08618-6
45. Routila J, Leivo I, Minn H, Westermarck J, Ventelä S. Evaluation of prognostic biomarkers in a population-validated Finnish HNSCC patient cohort. Eur Arch Otorhinolaryngol. 2021;278(11):4575–85. 10.1007/s00405-021-06650-7
46. Besse A, Sana J, Lakomy R, Kren L, Fadrus P, Smrcka M, et al. MiR-338-5p sensitizes glioblastoma cells to radiation through regulation of genes involved in DNA damage response. Tumour Biol. 2016;37(6):7719–27. 10.1007/s13277-015-4654-x
47. Tian Z, He W, Tang J, Liao X, Yang Q, Wu Y, et al. Identification of important modules and biomarkers in breast cancer based on WGCNA. Onco Targets Ther. 2020;13:6805–17. 10.2147/OTT.S258439