
Download
ORIGINAL ARTICLE
TRIM52 aggravated inflammation and pyroptosis in dextran sulfate sodium-induced inflammatory bowel disease through activation of the TLR4/NF-κBs pathway
Jin-ping Maa, Cheng-zi Yaoa, Zhi-qiang Jiaa, A-mei Wanga, Xiang-xia Miaoa, Xiang-xiang Gaoa, Li-ping Sub*
aDepartment of General Medicine, Xian Yang Central Hospital, Xianyang, China
bDepartment of Gastroenterology, Xian Yang Central Hospital, Xianyang, China
Abstract
Introduction: Inflammatory bowel disease (IBD), which mainly leads to diarrhea, fatigue, stool blood, abdominal pain, and cramping, is threatening public health. Tripartite motif-containing 52 (TRIM52) has been reported to play an important role in inflammatory responses via activating the toll-like receptor 4 (TLR4)/nuclear factor-κB (NF-κB) pathway. However, the causes of IBD need to be elucidated, and the function of TRIM52 in IBD remains unclear. Here, we demonstrated that TRIM52 aggravated inflammation and pyroptosis in dextran sulfate sodium (DSS)-induced IBD by activating TLR4/NF-κBs pathway.
Methods: The colitis model was established on mice through DSS induction. For the TRIM52 knockdown, the mice were infected with a recombinant adenoviral vector expressing sgRNAs targeting TRIM52. RT-qPCR, western blot, and immunohistochemistry were performed to verify TRIM52 expression in DSS-induced IBD. The body weight, disease activity index, colon length, and H&E staining were used to assess the IBD symptoms in mice with TRIM52 knockdown. The inflammatory responses were examined by RT-qPCR and ELISA measuring tumor necrosis factor-α (TNF-α), inter-leukin 6 (IL-6), and interleukin 1β (IL-1β). Furthermore, the pyroptosis in colon tissue was detected by western blot. Finally, the TLR4/NF-κBs pathway activity was also examined by western blot.
Results: TRIM52 expression was up-regulated in DSS-induced IBD, and knockdown of TRIM52 could alleviate the symptoms of IBD. TRIM52 knockdown retarded DSS-induced inflammatory response and inhibited DSS-induced pyroptosis in colon tissue. In addition, TRIM52 played a role in activating TLR4/NF-κBs pathway.
Conclusion: Knockdown of TRIM52 alleviated inflammation and pyroptosis in IBD by regulating TLR4/NF-κBs pathway. TRIM52 is expected to be a novel diagnostic indicator for IBD and a target of therapeutic treatment.
Key words: inflammatory bowel disease, NF-κB, pyroptosis, TLR4, TRIM52
*Corresponding author: Li-ping Su, Department of Gastroenterology, Xian Yang Central Hospital, No. 78, Renmin East Road, Xian Yang, Shaanxi, China. Email address: [email protected]
Received 21 July 2022; Accepted 8 August 2022; Available online 1 January 2023
Copyright: Ma J, et al.
License: This open access article is licensed under Creative Commons Attribution 4.0 International (CC BY 4.0). http://creativecommons.org/licenses/by/4.0/
Introduction
Inflammatory bowel disease (IBD) is a common chronic inflammatory gastrointestinal disorder that includes ulcerative colitis and Crohn’s disease. IBD symptoms mainly include weight loss, abdominal pain, chronic diarrhea, and colon shortening in length. Although the causes of IBD need to be elucidated, its pathogenesis has been reported to be related to irregular immune responses, gut microbiota, and genetic and environmental factors.1,2
Pyroptosis is one kind of programmed cell death dependent on caspase-1 characterized by the cleavage and trans-location of Gasdermin D (GSDMD) and the rapid rupture of the plasma membrane, resulting in the release of cytokines to activate pro-inflammatory immune cell mediators, including interleukin 1β (IL-1β) and IL-18.3,4 In IBD, however, pyroptosis promotes increased intestinal epithelial cell death and disrupts the intestinal barrier, resulting in increased intestinal permeability.5,6 Therefore, inhibiting the pyroptosis of intestinal epithelial cells can alleviate the symptoms of IBD.
Tripartite motifs (TRIMs) are an ancient family of proteins that play major roles in respective cellular processes. As members of the RING-type ubiquitin E3 ligase, the TRIM family proteins have been extensively studied to exert functions in regulating the nuclear factor-κB (NF-Κb) signal pathway.7 Tripartite motif-containing 52 (TRIM52), as a member of the TRIM protein, also plays an important role in immune responses. For example, TRIM52 silencing reduces the lipopolysaccharide (LPS)-induced inflammatory response of human periodontal ligament cells by inhibiting the activation of the toll-like receptor 4 (TLR4)/NF-κB signaling pathway, providing an effective therapeutic target for periodontitis.8 The levels of TRIM52 and NF-κB are increased in benign prostatic hyperplasia (BPH) tissues, while TRIM52 knockdown inhibits the proliferation of BPH-1 cells and reduces the promoter activity at the NF-κB region.9 TRIM52 activates the NF-κB signaling pathway by promoting the ubiquitination of IκBα protein, thereby promoting LPS-induced microglia activation and increasing neuroinflammation.10 In addition, TRIM52 plays a vital role in regulating tumors as well. Overexpression of TRIM52 can promote the proliferation of colon cancer cells and inhibit apoptosis.11
Given the reported functions of TRIM52 in inflammation and immune responses, we investigated the function of TRIM52 in IBD. Firstly, TRIM52 expression was measured in dextran sulfate sodium (DSS)-induced IBD. And the symptoms and inflammatory response were evaluated in TRIM52 knockdown mice. Furthermore, the pyroptosis and TLR4/ NF-κBs pathway activity were studied in the colon tissue of TRIM52 knockdown mice. These data demonstrated that TRIM52 aggravated inflammation and pyroptosis reaction in IBD by regulating TLR4/NF-κBs pathway.
Materials and Methods
Animals
Wild-type C57BL/6 mice and Cas9 transgenic mice were purchased from SLAC experimental animal center (Changsha, China). All animal procedures were approved by the Animal Care and Use Committee of Xian Yang Central Hospital.
For the DSS-induced colitis model, the mice were subjected to 4 cycles of DSS induction, including 7 days with 1.5% DSS in drinking water, followed by normal drinking water during a 10-day recovery period. Mice received normal drinking water for 4 weeks after the last DSS cycle.12 The sham group animals were administered distilled water. Each group contains six mice.
Twelve DSS-induced mice or 12 sham mice were randomly divided into the Ad-shNC group and Ad-shTRIM52 group. To generate TRIM52 knockdown mice, two sgRNAs were designed as follows: CAGCTGCCCGCAATGCCGA (sgRNA1) and GCCACCAGTACCATCGGTT (sgRNA2) using an online platform (http://crispr.dfci.harvard.edu/SSC/).13 To generate TRIM52 knockdown mice, the recombinant adenoviral vector expressing sgRNAs targeting TRIM52 was used to infect Cas9 transgenic mice. Approximately 1 × 1011 genome copies of adenovirus were injected via the tail vein for adenovirus transduction. The TRIM52 expression was examined by western blot in these transgenic mice.
Quantitative real-time PCR (qRT-PCR)
Total RNA was isolated using TRIzol reagent (Cat No. 15596026, Invitrogen, USA) following the protocol. The RNA concentration was quantitated by Nanodrop 2000 spectrophotometer. The reverse transcription was conducted with 500 ng RNA by Prime Script RT Reagent Kit (Takara, Dalian, China). Bio-Rad real-time PCR system was used to do qRTPCR with iTaq™ Universal SYBR® Green Supermix (Cat No. 1725121, Bio-Rad Laboratories Inc., Hercules, CA, USA). The PCR procedures contain 95°C for 5 min for one cycle, followed by 95°C for 30 s, and 60°C for 45 s for 40 cycles. The transcription of target genes was determined by 2-ΔΔCT, and the relative transcription was normalized to the β-actin level. The PCR primer sequences are listed in Table 1.
Table 1 Primers for RT-qPCR.
| Target gene | Sequence (5’-3’) |
|---|---|
| TRIM52 | GCCATCTGCTTGGATTACTTC |
| TTCATCTTCCTCCTCGTTCTG | |
| TNF-α | CCCAGGCAGTCAGATCATCTTC |
| AGCTGCCCCTCAGCTTGA | |
| IL-6 | GGTACATCCTCGACGGCATCT |
| GTGCCTCTTTGCTGCTTTCAC | |
| IL-1β | AACAGGCTGCTCTGGGATTC |
| AGTCATCCTCATTGCCACTGT | |
| β-actin | GTCTGCCTTGGTAGTGGATAATG |
| TCGAGGACGCCCTATCATGG |
Western blot
The lysates of colon tissue were electrophoresed on an SDS-PAGE gel, transferred into a polyvinylidene fluoride membrane, and then processed for immunoblot with primary antibodies listed in Table 2. The membranes were then incubated with the secondary antibodies named HRP-conjugated goat anti-mouse or anti-rabbit IgG as the secondary antibodies (Cat No. SA00001-1, SA00001-2, ProteinTech Group; 1:5000) and finally visualized with Immun-Star AP Chemiluminescence Kits (Bio-Rad Laboratories, Inc., Hercules, CA, USA). For the western blot signal, the relative intensity of each band was measured by ImageJ software, and the relative expression level was normalized to β-actin levels.14
Table 2 Antibodies information.
| Protein | Cat. No. | Manufacturer | Dilution |
|---|---|---|---|
| TRIM52 | sc-135589 | Santa Cruz Biotechnology, USA | 1:1000 |
| β-actin | ab8227 | Abcam, Cambridge, MA, USA | 1:1000 |
| NLRP3 | D4d8T | Cell Signaling Technology, MA, USA | 1:1000 |
| ASC | sc-271054 | Santa Cruz Biotechnology, USA | 1:500 |
| GSDMD-N | Ab209845 | Abcam, Cambridge, MA, USA | 1:1000 |
| caspase 1 | D7F10 | Cell Signaling Technology, MA, USA | 1:1000 |
| IL-1β | D3U3E | Cell Signaling Technology, MA, USA | 1:1000 |
| IL-18 | ab207323 | Abcam, Cambridge, MA, USA | 1:1000 |
| TLR4 | 14385S | Cell Signaling Technology, MA, USA | 1:1000 |
| MyD88 | ab219413 | Abcam, Cambridge, MA, USA | 1:1000 |
| P65 NF-κB | ab32536 | Abcam, Cambridge, MA, USA | 1:1000 |
| phospho-P65 NF-κB | 3033S | Cell Signaling Technology, MA, USA | 1:1000 |
| IκBα | CST4814 | Cell Signaling Technology, MA, USA | 1:1000 |
| phospho-IκBα | 9246S | Cell Signaling Technology, MA, USA | 1:1000 |
Immunohistochemistry (IHC) analysis
The colon tissues were fixed by formalin, embedded with paraffin, and then cut into 5 μm thick sections. 0.01 M citrate sodium buffer (pH 6.0) was used in antigen retrieval at 100°C for 10 min. Endogenous peroxidase and non-specific binding were blocked with 0.3% H2O2, and 10% bovine serum albumin, respectively. Then, the sections were incubated with primary antibody against TRIM52 overnight at 4°C. After washing three times, the incubation with anti-rabbit IgG was performed for 1 h at room temperature. Finally, the sections were developed in DAB substrate and counterstained with hematoxylin.15
H&E staining
For histological analysis, the colon tissues were dissected and fixed with 10% formalin at 4°C overnight and embedded into paraffin. The fixed tissues were cut into 5 mm thick sections, placed on glass slides and. Sections were deparaffinized on slides and stained with hematoxylin and eosin (HE). Finally, the sections were observed under a light microscope.16
Assessment of disease activity index (DAI)
For disease activity index, the body weight, stool consistence, and blood in the stool were assessed according to grading system published previously.17 The weight loss score contains: no loss, score 0; 1–5% loss, score 1; 5–10% loss, score 2; 10–20% loss, score 3; >20% loss, score 4. Diarrhea grading includes: normal, score 0; loose stools, score 2; watery diarrhea, score 4. Blood in stool was scored as follows: no bleeding, score 0; slight bleeding, score 2; gross bleeding, score 4. The total DAI score, ranging from 0 (normal) to 12 (severe colitis), represents the assessment of disease severity.
Enzyme-linked immunosorbent assay
The concentrations of tumor necrosis factor-α (TNF-α), interleukin 6 (IL-6), and IL-1β in the supernatants were determined by related ELISA Kit (ab208348, ab222503, ab100704, Abcam, Cambridge, MA, USA) according to manufacturer’s instructions. Briefly, 100 μl sample was incubated 2 h in ELISA well, after being washed five times, 100 μl detected antibody was added and incubated for 1 h. Each well was washed five times and incubated with 100 μl enzyme working reagent for 30 min. Then each well was washed for five times and incubated with 100 μl TMB reagent for 30 min. Finally, the reaction was terminated by 50 μl stop solution. The absorbance value was read at 450 nm, wavelength correction at 570 nm.
Statistical Analysis
All experiments for quantification and statistical analysis were repeated at least three times. The data analysis was performed using the SPSS 22.0 statistical software. The data between two groups are more consistent with normality and homogeneity of variance using independent sample t test. ** means the comparison to sham or sham+Ad-shNC; # (#) means comparison between DSS+Ad-shTRIM52 and sham+ Ad-shTRIM52; && means comparison between DSS+AdshTRIM52 and DSS+Ad-shNC. Both one star (P < 0.05) and two stars (P < 0.01) were considered statistically significant.
Results
TRIM52 expression was up-regulated in DSS-induced IBD
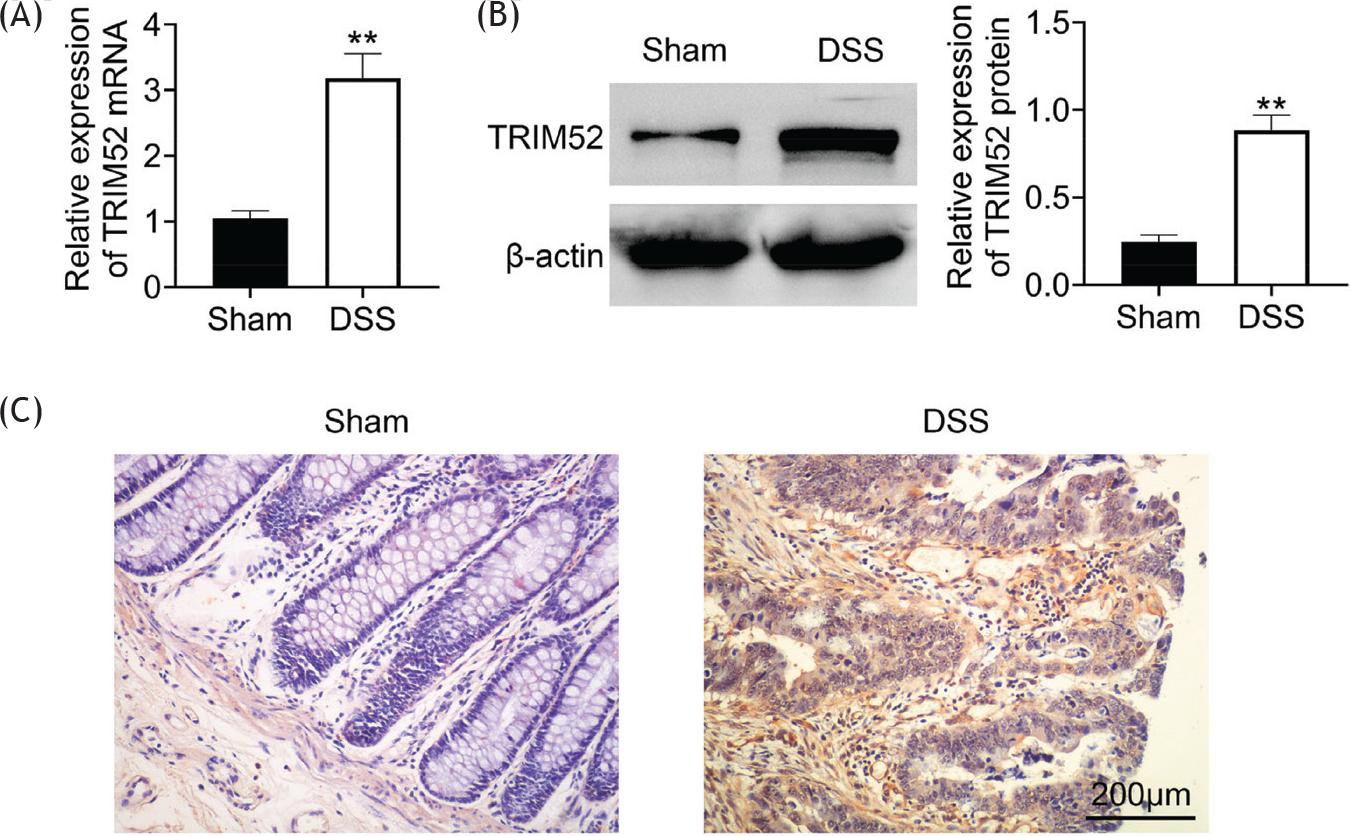
To examine TRIM52 expression in DSS-induced IBD, the mRNA level was firstly measured using RT-qPCR, and the results revealed that the TRIM52 transcription was elevated approximately three times in DSS-induced group than that in sham group (Figure 1A). Meanwhile, the protein level of TRIM52 was detected by western blot, and the data investigated that TRIM52 was significantly increased in DSS-induced mice (Figure 1B). Furthermore, IHC staining was performed in colon tissues of these mice. In DSS-induced mice, the TRIM52 positive signal was much more than in sham mice (Figure 1C). Thus, TRIM52 expression was increased in DSS-induced IBD.
Figure 1 TRIM52 expression was up-regulated in DSS-induced IBD. RT-qPCR was performed to examine TRIM52 transcription (A). The protein level of TRIM52 was measured by western blot (B). IHC staining was performed to detect the TRIM52 expression in colon tissues (C).
Knockdown of TRIM52 alleviated DSS-induced IBD symptoms
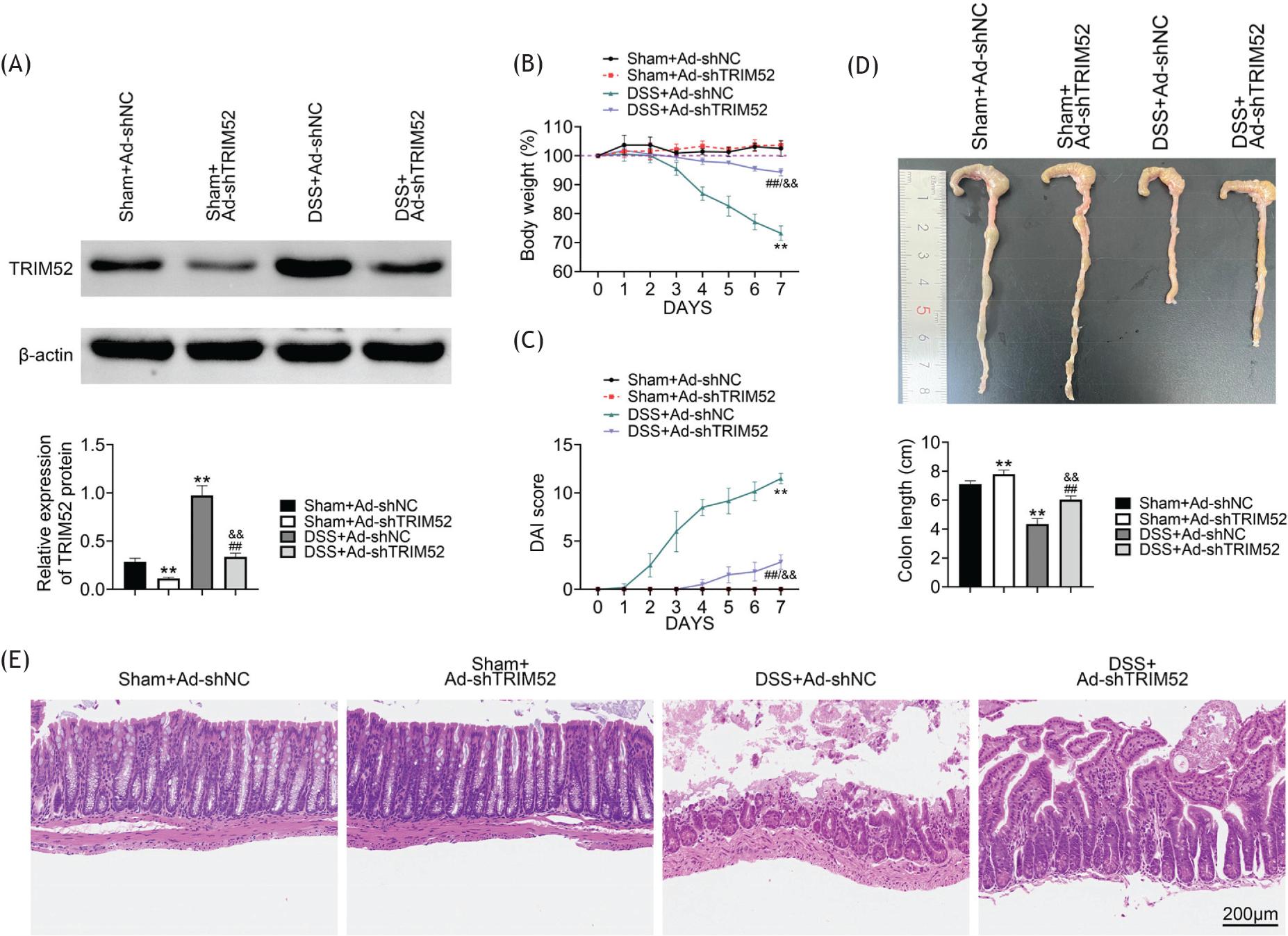
To study the function of TRIM52 in IBD, we established TRIM52 knockdown mice using recombinant adenovirus inserted with shRNA to TRIM52 (AD-shTRIM52). The TRIM52 expression was confirmed by western blot in TRIM52 knockdown mice. Specifically, TRIM52 expression with AD-shTRIM52 injection was decreased than with Ad-shNC. The DSS induction could up-regulate TRIM52 expression, however, the TRIM52 gene knockdown by AD-shTRIM52 also declined significantly TRIM52 protein level than that by Ad-shNC (Figure 2A). Next, we observed the body weight of mice with DSS-induced IBD. During the first 7 days in DSS-induced mice, the body weight decreased because of IBD. The TRIM52 knockdown by AD-shTRIM52 could attenuate the loss of body weight (Figure 2B). Disease activity index was used to assess the severity of IBD. As shown in Figure 2C, DSS could induced severe disease in DSS+Ad-shNC group, but TRIM52 knockdown by AD-shTRIM52 alleviated the disease symptoms to a large extent (Figure 2C). Moreover, the length of colon was shortened in DSS-induced mice than in sham mice. In DSS-induced mice, TRIM52 knockdown could recover some length of colon (Figure 2D). Finally, HE staining of colon indicated that the tissue structure and cytology of the control group were normal, and no inflammatory reaction was observed. However, DSS treatment resulted in severe diffuse destruction of the colonic epithelial layer with extensive inflammatory cell infiltration of the epithelium and lamina propria. Compared with DSS+ Ad-shNC group, the mucosal infiltration of inflammatory cells and tissues was significantly reduced in DSS plus TRIM52 knockdown group (Figure 2E). Together, TRIM52 knockdown could alleviate DSS-induced IBD symptoms.
Figure 2 TRIM52 knockdown alleviated DSS-induced IBD symptoms. The TRIM52 expression was confirmed by western blot in TRIM52 knockdown mice (A). The body weight of mice with TRIM52 knockdown in DSS-induced IBD (B). The disease activity index was used to assess the severity of IBD (C). The loss of colon length was alleviated by TRIM52 knockdown in DSS-induced IBD (D). HE staining of the colon to investigate TRIM52 effect on tissue structure, cytology, and inflammatory reaction (E).
Knockdown of TRIM52 inhibited DSS-induced inflammatory response
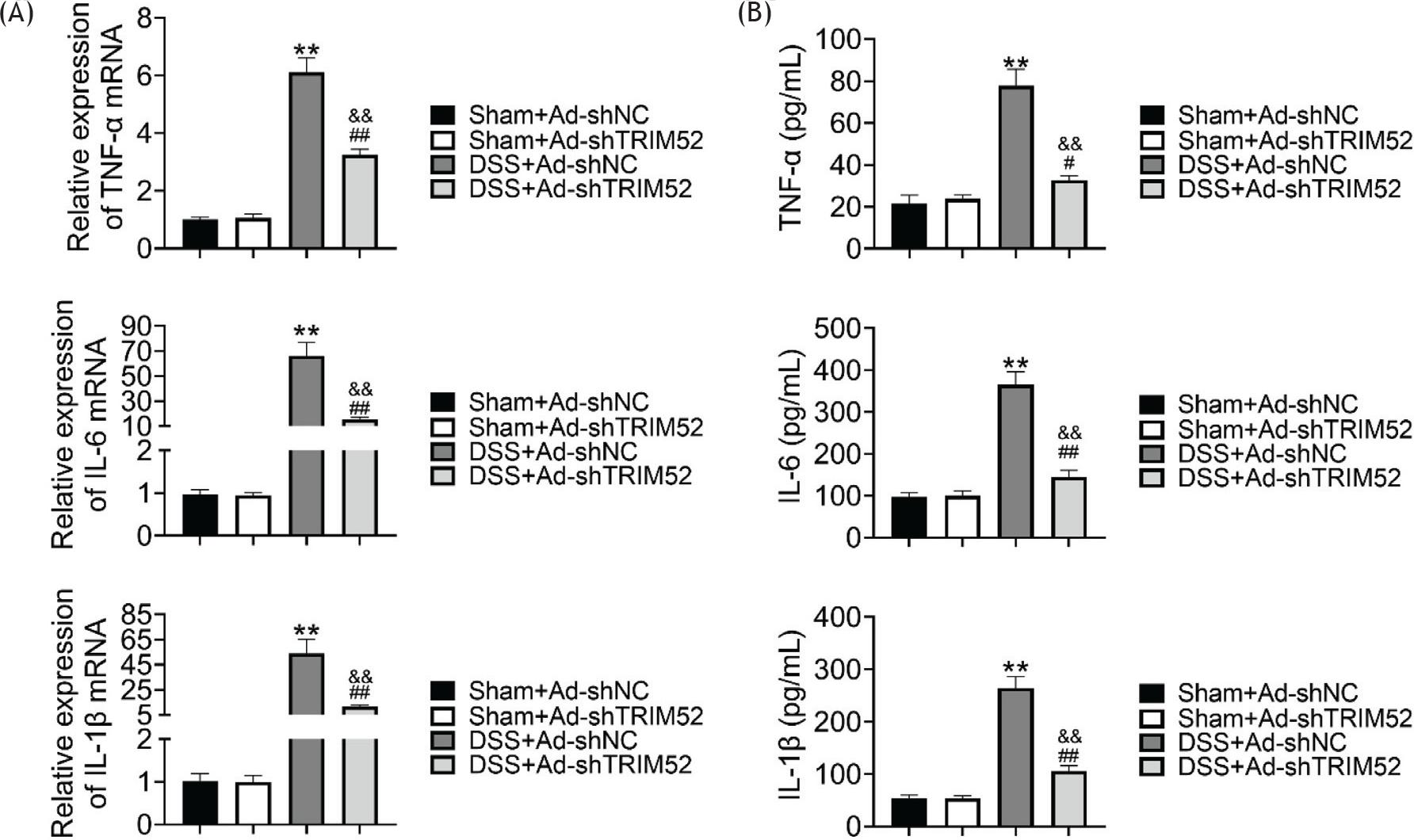
To study the function of TRIM52 in DSS-induced inflammatory response, we examined the expression of typical cytokines in inflammatory response including TNF-α, IL-6, and IL-1β. The RT-qPCR results showed the transcription of these cytokines were up-regulated significantly in DSS-induced mice than in sham mice. Meanwhile, in DSS-induced groups, TRIM52 knockdown could decline the mRNA level of TNF-α, IL-6, and IL-1β (Figure 3A). In addition, the enzyme-linked immunosorbent assay was performed to measure the protein level of these cytokines. The data demonstrated that the protein level of TNF-α, IL-6, and IL-1β were also elevated in DSS-induced inflammatory, and the TRIM52 knockdown in DSS-treated mice alleviated the inflammatory response indicated by TNF-α, IL-6, and IL-1β expression level (Figure 3B). Therefore, TRIM52 knockdown retarded DSS-induced inflammatory response.
Figure 3 TRIM52 knockdown inhibited DSS-induced inflammatory response. RT-qPCR was performed to examine the transcription of TNF-α, IL-6, and IL-1β in DSS-induced inflammatory response (A). Enzyme-linked immunosorbent assay was performed to measure the protein level of TNF-α, IL-6, and IL-1β in DSS-induced inflammatory response (B).
Knockdown of TRIM52 inhibited DSS-induced pyroptosis in colon tissue
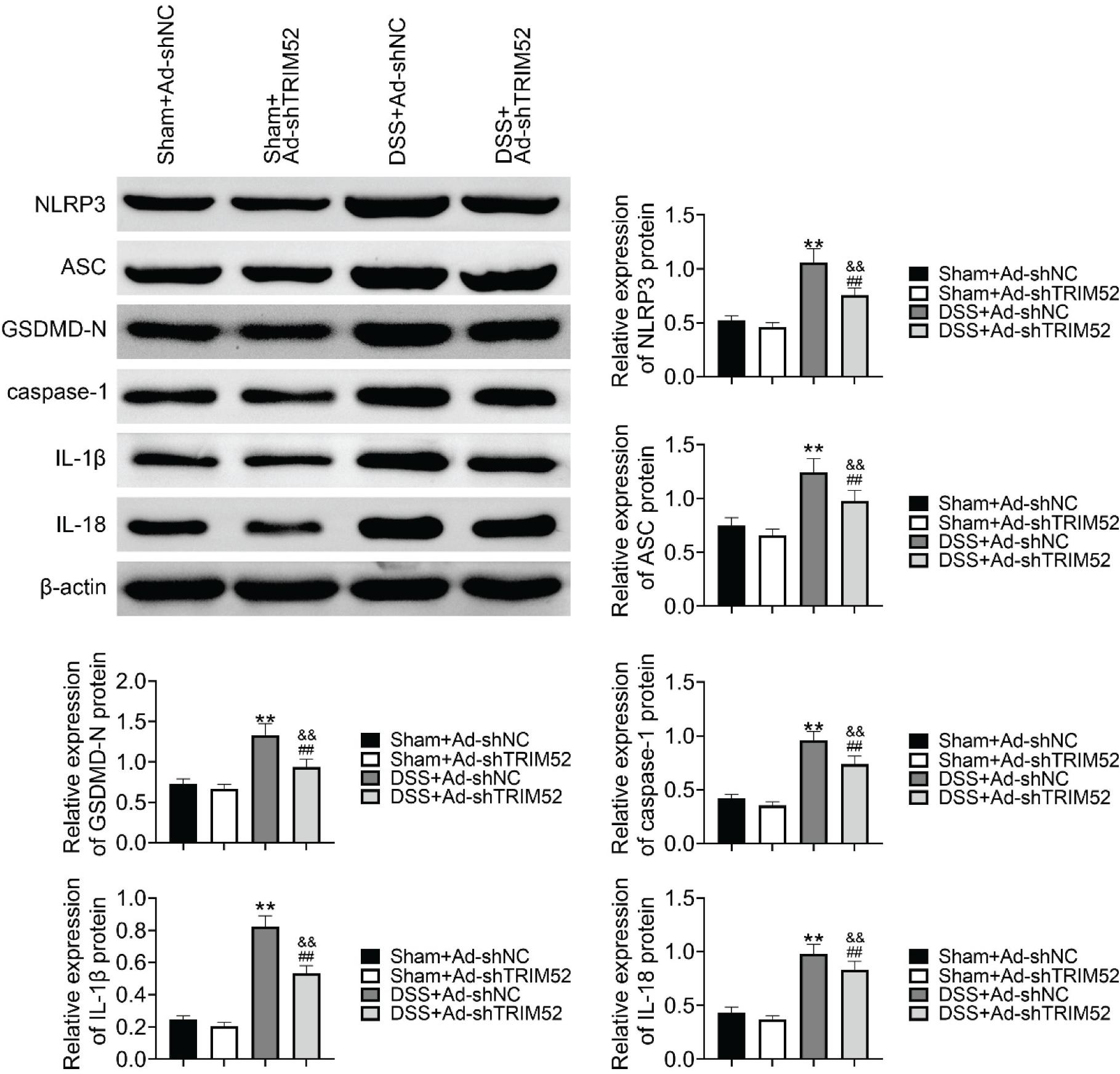
To clarify whether TRIM52 affects DSS-induced pyroptosis in colon tissue, western blot was performed to examined the expression level of NLRP3, ASC, GSDMD-N, caspase-1, IL-1β, IL-18 of which was normalized β-actin level. The results revealed that DSS treatment could induce pyroptosis in colon tissue which was supported by elevated expression of NLRP3, ASC, GSDMD-N, caspase-1, IL-1β, IL-18 in DSS+Ad-shNC compared to sham+Ad-shNC or DSS+Ad-shTRIM52 compared to sham+Ad-shTRIM52. In addition, the DSS-induced pyroptosis could be alleviated by TRIM52 knockdown as a declined expression of NLRP3, ASC, GSDMD-N, caspase-1, IL-1β, IL-18 in DSS+Ad-shTRIM52 compared to DSS+Ad-shNC (Figure 4). The results above demonstrated that TRIM52 knockdown inhibited DSS-induced pyroptosis in colon tissue.
Figure 4 TRIM52 knockdown inhibited DSS-induced pyroptosis in colon tissue. Western blot was performed to examine the expression level of NLRP3, ASC, GSDMD-N, caspase-1, IL-1β, and IL-18. The results revealed that DSS treatment could induce pyroptosis in colon tissue, and the DSS-induced pyroptosis could be alleviated by TRIM52 knockdown.
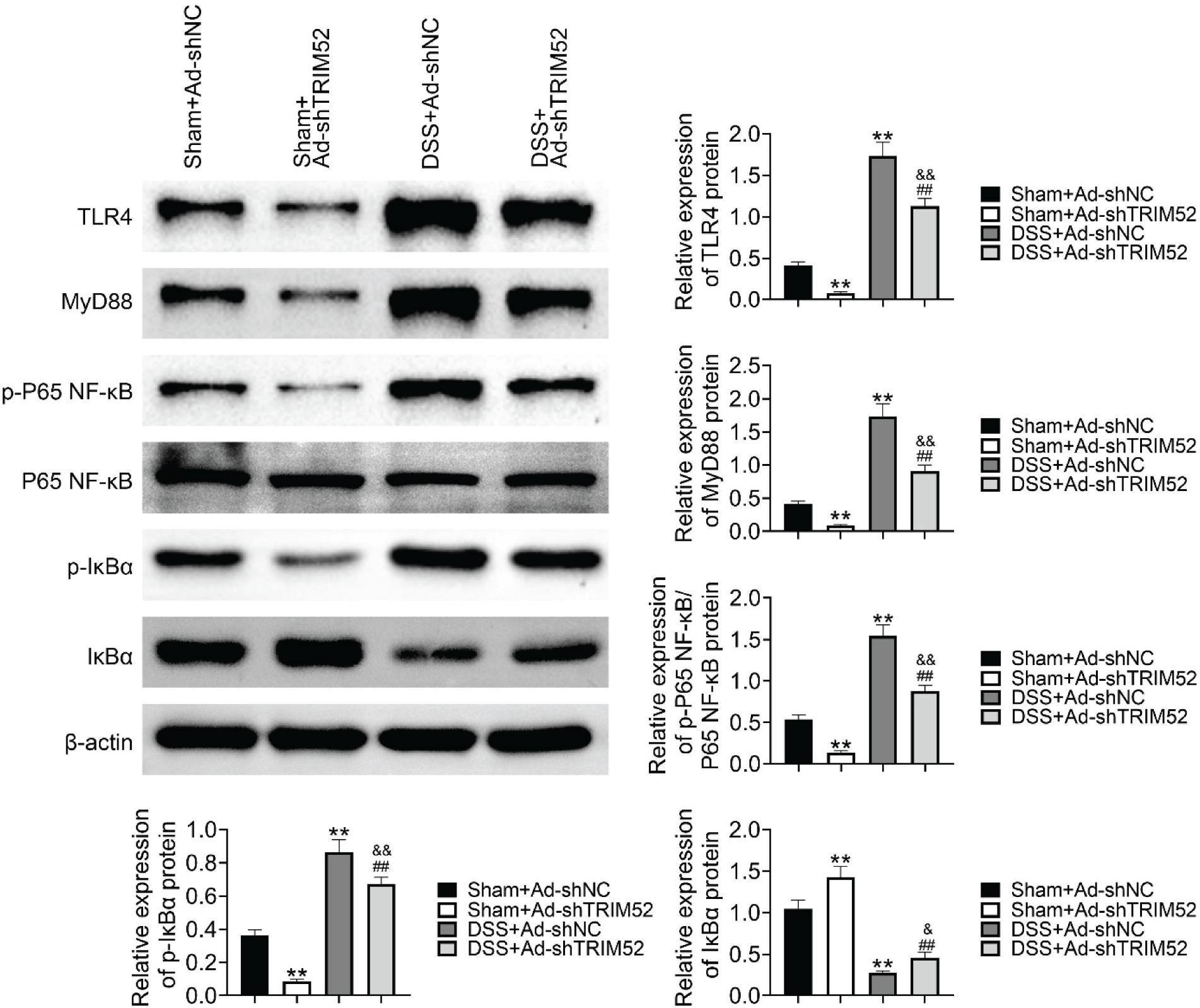
TRIM52 promoted TLR4/NF-κBs pathway activity
TRIM52 has been reported to regulate the TLR4/NF-κB pathway; here, we investigated the role of TRIM52 in DSS-induced regulation of the TLR4/NF-κB pathway. Western blot was used to examine the expression of TLR4, MyD88, phosphorated P65 NF-κB, phosphorated-IκBα, and IκBα. Specifically, in sham mice, TRIM52 knockdown decreased the expression of TLR4, MyD88, phosphorated P65 NF-κB, and phosphorated-IκBα but increased IκBα expression. DSS treatment could activate TLR4/NF-κBs pathway in colon tissue, which was demonstrated by elevated expression of TLR4, MyD88, phosphorated P65 NF-κB, phosphorated-IκBα in DSS+Ad-shNC compared to sham+Ad-shNC or DSS+AdshTRIM52 compared to sham+Ad-shTRIM52. In addition, the DSS-induced activation of the TLR4/NF-κBs pathway could be weakened by TRIM52 knockdown because of declined expression of TLR4, MyD88, phosphorated P65 NF-κB, phosphorated-IκBα, and increased expression of IκBα in DSS+Ad-shTRIM52 compared to DSS+Ad-shNC (Figure 5). The data together revealed TRIM52 knockdown weakened TLR4/NF-κBs pathway activity, indicating that TRIM52 played a role in activating TLR4/NF-κBs pathway.
Figure 5 TRIM52 promoted TLR4/NF-κBs pathway activity. Western blot was used to examine the expression of TLR4, MyD88, phosphorated P65 NF-κB, phosphorated-IκBα and IκBα. DSS treatment could activate TLR4/NF-κBs pathway in colon tissue, and the DSS-induced activation of the TLR4/NF-κBs pathway could be weakened by TRIM52 knockdown.
Discussion
IBD, a worldwide healthcare problem, has been increasing in incidence recently. TRIM52 has also been reported playing an important role in inflammatory responses including activation of TLR4/NF-κB signaling pathway.8 To explore whether TRIM52 regulates inflammatory response in DSS-induced IBD, we measured the expression level of TIRM52 in DSS-induced mice. The increased expression of TRIM52 provided us a clue that TIRM52 most likely exerted functions in IBD. To be further, the knockdown of TRIM52 weakened the LPS-induced inflammatory response via inhibiting TLR4/NF-κB signaling pathway.8 Overexpression of TRIM52 promoted the proliferation of colon cancer cells and retarded cell apoptosis.11 Thus, it would be promising that TRIM52 expression is upregulated in IBD.
Next, we examined the TRIM52 role in inflammatory response, and found TRIM52 silencing inhibited DSS-induced inflammatory response. It was reported that M1 macrophages could be induced by pathogen-associated molecular patterns, such as LPS or IFNγ, producing a series of cytokines including IL-1β, TNF-α, IL-6 to aggravate inflammation.18 TRIM52 knockdown weakened the LPS-induced inflammatory response via inhibiting TLR4/ NF-κB signaling pathway.8 So, it is reasonable that TRIM52 expression positively regulated inflammatory response in DSS-induced mice.
Pyroptosis mainly activated by NLRP3 inflammasome and caspases is a major cause of inflammatory disorders. NLRP3 and CAS1, the hallmarks of pyroptosis, were increasingly expressed in periodontal disease compared to healthy tissue.19,20 It was reported that NLRP3/caspase-1/GSDMD-mediated pyroptosis exerts a crucial role in astrocyte pathological injury in mouse model of depression.21 In vascular endothelial cells, pyroptosis could be caused by NLRP3/caspase-1/GSDMD and caspase-3/GSDME pathways which were promoted by histone deacetylase 11 (HDAC11).22 And pyroptosis could be induced by cisplatin via activating MEG3/ NLRP3/caspase-1/GSDMD pathway in triple-negative breast cancer.23 In our study, we examined the pyroptosis in TRIM52 knockdown mice through measuring the expression level of NLRP3, ASC, GSDMD-N, and caspase-1. The final results showed TRIM52 knockdown inhibited DSS-induced pyroptosis in colon tissue.
The IBD are thought to originate from dysregulation of innate and adaptive immune systems, resulting in abnormal inflammatory response to commensal bacteria.24 The diarrhea-predominant irritable bowel syndrome could be alleviated by momordica charantia polysaccharides which regulated intestinal inflammation and barrier via NF-κB pathway.25 It has been reported that TRIM52 expression was positively correlated with NF-κB expression. TRIM52 silencing decreased the promoter activity of NF-κB, and the NF-κB inhibitor, pyrrolidine dithiocarbamate (PDTC), inhibited NF-κB promoter activity.9 TRIM52 knockdown alleviated LPS-induced inflammatory injury through TLR4/NF-κB pathway.8 In this study, we also investigated the TLR4 and key molecules in NF-κBs pathway. Similarly, TRIM52 knockdown weakened TLR4/NF-κBs pathway activity, showing that TRIM52 played a role in activating TLR4/NF-κBs pathway.
In this study, we revealed that TRIM52 expression was up-regulated in DSS-induced IBD. Moreover, TRIM52 promoted inflammatory response and pyroptosis in DSS-induced IBD by promoting the activity of TLR4/NF-κBs pathway. While further studies remain to be conducted, including molecular mechanism of inflammatory response and pyroptosis, the regulation pathways of overexpression of TRIM52. More detailed data would promote TRIM52 as a novel diagnostic indicator for IBD, and make it as a potential target for developing novel therapeutic treatment.
Acknowledgements
Not applicable.
Funding
Not applicable.
Availability of Data and Materials
All data generated or analyzed during this study are included in this published article.
Competing Interests
The authors state that there are no conflicts of interest to disclose.
Ethics Approval
Ethical approval was obtained from the Ethics Committee of Xian Yang Central Hospital.
Author’s Contribution
Conceptualization, methodology, and writing - original draft were performed by Jin-ping Ma and Cheng-zi Yao; Formal analysis, resources, and Investigation were performed by Zhi-qiang Jia; formal analysis, visualization, and data curation were performed by A-mei Wang; project administration, supervision, and validation were performed by Xiang-xia Miao; validation, supervision, and writing - review & editing were performed by Xiang-xiang Gao and Li-ping Su. All authors read and approved the final manuscript.
REFERENCES
1. De Souza HS, Fiocchi C. Immunopathogenesis of IBD: current state of the art. Nat Rev Gastroenterol Hepatol. 2016;13(1):13–27. 10.1038/nrgastro.2015.186
2. Baumgart DC, Carding SR. Inflammatory bowel disease: cause and immunobiology. Lancet. 2007;369(9573):1627–40. 10.1016/S0140-6736(07)60750-8
3. Coll NS, Epple P, Dangl JL. Programmed cell death in the plant immune system. Cell Death Differ. 2011;18(8):1247–56. 10.1038/cdd.2011.37
4. Ma Y, Jiang J, Gao Y, Shi T, Zhu X, Zhang K, et al. Research progress of the relationship between pyroptosis and disease. Am J Transl Res. 2018;10(7):2213.
5. Gibson PR. Increased gut permeability in Crohn’s disease: is TNF the link? Gut. 2004;53(12):1724–5. 10.1136/gut.2004.047092
6. Xu Z, Liu R, Huang L, Xu Y, Su M, Chen J, et al. CD147 aggravated inflammatory bowel disease by triggering NF-κB-Mediated pyroptosis. Biomed Res Int. 2020; 2020. 10.1155/2020/5341247
7. Tomar D, Singh R. TRIM13 regulates ubiquitination and turnover of NEMO to suppress TNF induced NF-κB activation. Cell Signal. 2014;26(12):2606–13. 10.1016/j.cellsig.2014.08.008
8. Liu P, Cui L, Shen L. Knockdown of TRIM52 alleviates LPS-induced inflammatory injury in human periodontal ligament cells through the TLR4/NF-κB pathway. Biosci Rep. 2020;40(8):BSR20201223. 10.1042/BSR20201223
9. Sun J, Liu P, Mao J, Han W, Jiang X, Gu Y. TRIM52 positively mediates NF-κB to promote the growth of human benign prostatic hyperplasia cells through affecting TRAF2 ubiquiti-nation. Life Sci. 2020; 259:118380. 10.1016/j.lfs.2020.118380
10. Zhang P, Wu Y, Li R, Lv H, Yu B. Tripartite motif containing 52 positively regulates NF-κB signaling by promoting IκBα ubiquitination in lipopolysaccharide-treated microglial cell activation. Med Sci Monit Basic Res. 2020;26:e925356-925351. 10.12659/MSM.925356
11. Pan S, Deng Y, Fu J, Zhang Y, Zhang Z, Ru X. TRIM52 promotes colorectal cancer cell proliferation through the STAT3 signaling. Cancer Cell Int. 2019;19(1):1–11. 10.1186/s12935-019-0775-4
12. Han X, Wu Y, Zhuang Y, Zhang J. Dexmedetomidine reduces dextran sulfate sodium (DSS)-induced NCM460 cell inflammation and barrier damage by inhibiting RhoA/ROCK signaling pathway. Allergol Immunopathol. 2022;50(3):85–92. 10.15586/aei.v50i3.569
13. Xu H, Xiao T, Chen C-H, Li W, Meyer CA, Wu Q, et al. Sequence determinants of improved CRISPR sgRNA design. Genome Res. 2015;25(8):1147–57. 10.1101/gr.191452.115
14. Rahmati M, Keshvari M, Xie W, Yang G, Jin H, Li H, et al. Resistance training and Urtica dioica increase neurotrophin levels and improve cognitive function by increasing age in the hippocampus of rats. Biomed Pharmacother. 2022;153:113306. 10.1016/j.biopha.2022.113306
15. Rahmati M, Taherabadi SJ. The effects of exercise training on Kinesin and GAP-43 expression in skeletal muscle fibers of STZ-induced diabetic rats. Sci Rep. 2021;11(1):1–9. 10.1038/s41598-021-89106-6
16. Chen X, Liu G, Yuan Y, Wu G, Wang S, Yuan L. NEK7 interacts with NLRP3 to modulate the pyroptosis in inflammatory bowel disease via NF-κB signaling. Cell Death Dis. 2019;10(12):1–12. 10.1038/s41419-019-2157-1
17. da Costa Gonçalves F, Schneider N, Mello HF, Passos EP, Meurer L, Cirne-Lima E, et al. Characterization of acute murine dextran sodium sulfate (DSS) colitis: severity of Infl ammation is dependent on the DSS molecular weight and concentration. Acta Sci Vet. 2013;41(1):1–9.
18. Gordon S, Martinez FO. Alternative activation of macrophages: mechanism and functions. Immunity. 2010;32(5):593–604. 10.1016/j.immuni.2010.05.007
19. Lu W, Song D, Yue J, Wang T, Zhou X, Zhang P, et al. NLRP 3 inflammasome may regulate the inflammatory response of human periodontal ligament fibroblasts in an apoptosis-associated speck-like protein containing a CARD (ASC)-dependent manner. Int Endod J. 2017;50(10):967–75. 10.1111/iej.12722
20. Yu C, Zhang C, Kuang Z, Zheng Q. The role of NLRP3 inflammasome activities in bone diseases and vascular calcification. Inflammation. 2021;44(2):434–49. 10.1007/s10753-020-01357-z
21. Li S, Sun Y, Song M, Song Y, Fang Y, Zhang Q, et al. NLRP3/ caspase-1/GSDMD–mediated pyroptosis exerts a crucial role in astrocyte pathological injury in a mouse model of depression. JCI Insight. 2021;6(23): e146852. 10.1172/jci.insight.146852
22. Yao F, Jin Z, Zheng Z, Lv X, Ren L, Yang J, et al. HDAC11 promotes both NLRP3/caspase-1/GSDMD and caspase-3/GSDME pathways causing pyroptosis via ERG in vascular endothelial cells. Cell Death Discov. 2022;8(1):1–11. 10.1038/s41420-022-00906-9
23. Yan H, Luo B, Wu X, Guan F, Yu X, Zhao L, et al. Cisplatin induces pyroptosis via activation of MEG3/NLRP3/caspase-1/ GSDMD pathway in triple-negative breast cancer. Int J Biol Sci. 2021;17(10):2606. 10.7150/ijbs.60292
24. Halpin SJ, Ford AC. Prevalence of symptoms meeting criteria for irritable bowel syndrome in inflammatory bowel disease: systematic review and meta-analysis. Am J Gastroenterol. 2012;107(10):1474–82. 10.1038/ajg.2012.260
25. Ji S, Zhang Q. Momordica charantia polysaccharides alleviate diarrhea-predominant irritable bowel syndrome by regulating intestinal inflammation and barrier via NF-κB pathway. Allergol Immunopathol. 2022; 50(3):62–70. 10.15586/aei.v50i3.584