
Download





ORIGINAL ARTICLE
Long noncoding RNA HOXA-AS2 ameliorates chronic intermittent hypoxia-induced lung inflammation by regulating miR-17-5p/tipe2 axis
Kun Gao1#*, Aiai Lv2#, Qiang Zhang1, Yanzhong Li3, Zhiyong Yue1, Shuai Xu1*
1Department of Otorhinolaryngology Head and Neck Surgery, Shandong Provincial Hospital Affiliated to Shandong First Medical University, Jinan, Shandong, China
2Department of Internal Medicine, Public Health Clinical Center, Affiliated to Shandong University, Jinan, Shandong, China
3Department of Otorhinolaryngology, Qilu Hospital of Shandong University, NHC Key Laboratory of Otorhinolaryngology (Shandong University), Jinan, Shandong, China
#These authors contributed equally to the work.
Abstract
Purpose: The purpose is to confirm whether long noncoding RNA HOXA-AS2 relieves chronic intermittent hypoxia (CIH)-induced lung inflammation.
Methods: Male Sprague Dawley rats were used to establisha CIH rat model. Hematoxylin and Eosin staining was used on the lung tissue injury to determine the successful construction of CIH animal model. Arterial partial pressure of oxygen (PaO2) and carbon dioxide (PaCO2) were measured. HOXA-AS2 was overexpressed to evaluate its role in the progression and development of CIH. T cell differentiation and cytokine production were determined using flow cytometry. Cell apoptosis was determined using terminal deoxynucleotidyl transferase dUTP nick end labelling assay kit. The target of HOXA-AS2 and miR-17-5p was predicted by the Encyclopedia of RNA Interactomes (ENCORI) and confirmed using luciferase assay.
Results: HOXA-AS2 was downregulated in CIH rat models. Lung tissue injury was observed in CIH rats, and the injury was attenuated by the overexpression of HOXA-AS2. PaO2 was reduced and PaCO2 was induced in CIH rats, which was reversed by the overexpression of HOXA-AS2. The overexpression of HOXA-AS2 inhibited CIH-induced cell apoptosis. It also reversed alterations in the levels of interferon gamma (IFNγ), interleukin (IL)-2, IL-6, IL-1β, tumor necrosis factor alpha (TNF-α), and transforming growth factor beta1 (TGF-β1) in rats caused by CIH. The overexpression of HOXA-AS2 prevented the induction in CD4+ IFN-γ+ T cells and reduction in CD4+TGF-β1+ T cells. The overexpression of HOXA-AS2 upregulated tumor necrosis factor-alpha-induced protein 8-like 2 (tipe2) key regulator through directly targeting miR-17-5p. Further experiments proved that tipe2 was the direct target of miR-17-5p.
Conclusion: This study manifested that HOXA-AS2 acted as an anti-inflammatory regulator and protected lung tissue injury from CIH in the rat model; this was mediated by upregulation of tipe2 through directly targeting miR-17-5p. HOXA-AS2 upregulated the expression of tipe2, providing new understanding and therapeutic target for CIH.
Key words: chronic intermittent hypoxia, HOXA-AS2, lung inflammation, miR-17-5p, tipe2
*Corresponding authors: Kun Gao and Shuai Xu, Department of Otorhinolaryngology Head and Neck Surgery, Shandong Provincial Hospital Affiliated to Shandong First Medical University, No. 324 Jingwu Road, Jinan, Shandong 250021, China. Email addresses: [email protected] and [email protected]
Received 17 September 2022; Accepted 27 November 2022; Available online 1 March 2023
Copyright: Gao K, et al.
This open access article is licensed under Creative Commons Attribution 4.0 International (CC BY 4.0). http://creativecommons.org/licenses/by/4.0/
Introduction
Long-term chronic intermittent hypoxia (CIH) is usually accompanied by abnormality of cardiopulmonary circuit, including pulmonary hypertension, atherosclerosis, and lung inflammation.1–3 CIH-induced lung inflammation is involved in the production of many inflammatory molecules, including tumor necrosis factor α (TNF-α) and interleukins (ILs), such as IL-1β, IL-6, and IL-2.4 During inflammation, CD4+ T lymphocytes could differentiate into T helper 1 (Th1, pro-inflammatory) cells, which specifically release interferon gamma (IFNγ),5 while regulatory T (Treg, anti-inflammatory) cells are suppressed, demonstrating reduction in CD4+ transforming growth factor-beta 1+ (TGF-β1) T cells.6 Therefore, inhibition of lung inflammation is an effective path to prevent lung injury from CIH. Aerobic exercise, resistance exercise, and herbal medicine can successfully target inflammatory cytokines in chronic diseases,7–9 but there is a need to figure out the mechanism of lung inflammation to provide a new insight to CIH treatment.
Long noncoding RNAs (lncRNAs, ˃200 nucleotides [nt]) are a group of transcripts which have no coding potential but have the ability to bind to microRNAs (miRs) through base-pairing interactions, regulating protein expression.10 LncRNAs are dysregulated and involved into the pathophysiological process of CIH.11 miRs are a category of small, conserved RNAs (∼22 nucleotides) that are involved in almost all the pathophysiological process through targeting -protein-coding transcripts.12 miRs play an anti-inflammatory role in many diseases, including CIH and asthma.13–15 For example, lncRNA Mirt1 binds to miR-466i-5p to regulate target gene expression, demonstrating a critical regulator of myocardial infarction aggravated by CIH.16 HOXA cluster antisense RNA 2 (HOXA-AS2), a 1048-bp lncRNA, plays an anti-inflammatory role in different inflammatory -models.17,18 However, no publications has reported the role of HOXA-AS2 in CIH-induced lung inflammation.
Tumor necrosis factor-alpha-induced protein 8-like 2 (tipe2) is a key regulator of inflammation and immune homeostasis,19 and has strong anti-inflammatory properties.20 Loss of tipe2 is associated with the upregulation of pro-inflammatory cytokines, such as IL-6 and TNF-α, and downregulation of anti-inflammatory factors, such as TGF-β1, thereby promoting proliferation and differentiation of T lymphocytes.19 miR-21 has been revealed to promote T cell apoptosis by targeting tipe2.21 It is thus hypothesized that HOXA-AS2/miR-17-5p/tipe2 axis could play a functional role in the progression and development of CIH. However, limited literature has described the role of tipe2 in the progression of CIH. Therefore, this study was aimed to identify the anti-inflammatory role of HOXA-AS2/miR-17-5p/tipe2 in a CIH rat model through a series of experiments.
Methods
Establishment of CIH rat model
Male Sprague Dawley (SD) rats (n = 36, 200–250 g) were purchased from Jiangsu ALF Biotechnology Co. Ltd. (Jiangsu Province, China). Rats were kept under 12-h light/12-h dark cycle at 23±2°C with a humidity of 40±5% and provided standard animal food and drinking water. All experiment procedures were carried out in compliance with the ethical standards under a protocol approved by the Institutional Animal Care and Use Committee of Shandong Provincial Hospital Affiliated to Shandong First Medical University, and were executed conforming to the Care and Use of Laboratory Animals published by the US National Institutes of Health.22
The CIH model was established following the study conducted by Zhang et al.3 Briefly, SD rats were moved into sealed chambers away from the normal environment. Compressed air and nitrogen were infused into the chambers at a cycle of 90 s. To make sure a minimum of 5% oxygen concentration, pure nitrogen was infused into each chamber during the first 30 s. For the remaining 60 s, each chamber was infused with compressed air to make oxygen concentration delivery of 21%. Normal air was forced into the chambers of control group at a cycle of 90 s. Rats were randomly assigned into the following six groups (n = 6 rats/group): sham group: chambers were infused with normal air; CIH group: chambers were infused with compressed air and nitrogen; sham+AAV -HOXA-S2 group: sham rats were injected with adeno-associated virus (AAV) containing HOXA-S2 gene (AAV -HOXA-S2) through tail vein; sham-AAV-NC (negative control) group: sham rats were injected with AAV containing scramble sequence of HOXA-S2 gene (AAV-NC) through tail vein; CIH+AAV-HOXA-S2: CIH rats were injected with AAV-HOXA-S2 vector; CIH+AAV-NC: CIH rats were injected with AAV-NC vector.
Blood gas analysis
After establishing the CIH rat model, arterial partial pressure of oxygen (PaO2) and carbon dioxide (PaCO2) was measured using GEM Premier 4000 & 3500 (Instrumentation Laboratory, Lexington, MA, USA). At the end of the experiments, rats were scarified by an overdose of chloral hydrate (100 mg/kg). Lung tissues, spleen, and serum were collected for further experiments.
Hematoxylin and Eosin (H&E) staining
Lung tissues were fixed in 4% buffered formalin solution for 48 h at 4°C and embedded in paraffin wax. The fixed lung tissues were then cut into 5-µm sections and stained using the H&E staining kit (Powerful, China) according to the manufacturer’s protocols. The stained slides were photographed under a microscope (Nikon, Tokyo, Japan).
Terminal deoxynucleotidyl transferase dUTP nick end labelling (TUNEL) assay
Cell apoptosis was determined using TUNEL assay kit (Abcam, Cambridge, UK). Lung tissues were fixed in 4% buffered formalin solution for 48 h at 4°C and embedded in paraffin wax. The fixed lung tissues were then cut into 5-µm sections. Deparaffinization and dehydration were performed using xylene and graded ethanol, respectively. The tissue sections were incubated with 20-µg/mL recombinant Proteinase K solution (Thermo Fisher Scientific, Waltham, MA, USA) at 37°C for 30 min. The sections were washed for at least three times with phosphate buffered saline (PBS). The sections were incubated with DNA-labeling solution for 1 h at 37°C, followed by incubation with antibody solution for 30 min at room temperature. 4',6-Diamidino-2-phenylindole (DAPI; Abcam), 20 µL, was added to all slides, which were gently covered with cover slips. The slides were photographed under a fluorescent microscope (Nikon).
Flow cytometry
Flow cytometry was used to examine the ratio of CD4+IFN-γ+ T cells and CD4+TGF-β1+ T cells in the spleen. The spleen tissues were cut into small pieces and digested in 400-µ/mL collagenase III (Abcam) for 20 min at 37°C. Tissues were filtered using 200-mesh sieves and centrifuged at 1000 rpm for 5 min. Cells were resuspended in PBS, fixed using 4% buffered formalin solution for 1 h, and treated with 1% Trition X-100 (Merck KGaA, Darmstadt, Germany) for 5 min at room temperature. The fixed cells were stained with the following antibodies for flow cytometry: CD4 (Abcam), IFN-γ (Invitrogen/Thermo Fisher Scientific), and TGF-β1 (Shanghai Yaji Biotechnology Co. Ltd., Shanghai, China). In all, 30,000 stained cells were collected and analyzed using Gallios flow cytometer (Beckman Coulter, Fullerton, CA, USA).
Quantitative real-time polymerase chain reaction (qRT-PCR)
Lung tissues were cut into small pieces and lysed using TRIzol RNA isolation reagents (Invitrogen). Tissues were centrifuged at 5000 rpm for 10 min, the supernatant was collected, and Multiskan SkyHigh microplate spectrophotometer (Thermo Fisher Scientific) was used to determine RNA concentration. QuantiTect reverse transcription kit (QIAGEN GmbH, Hilden, Germany) was used to synthesize complementary DNA (cDNA) from 300-ng total RNA. qRT-PCR was performed using SYBR™ Green PCR Master Mix (Thermo Fisher Scientific) through ProFlex PCR System (Thermo Fisher Scientific). The relative RNA expression was quantified using the 2-ΔΔCT method. Primary sequences (Sigma-Aldrich Corp., Milwaukee, WI, USA) used in this study are listed in Table 1.
Table 1 The primary sequences used in this study.
| Gene name | Forward (5’-3’) | Reverse (5’-3’) |
|---|---|---|
| HOXA-AS2 | AACCCATCTTTGCCTTCTGC | CGGAGGAGTTTGGAGTTGG |
| Tipe2 | GTGACTGACCACATACCCCA | AGTGTTAGTGCCAGGTGAGC |
| miR-17-5p | GGCAAAGTGCTTACAGTGC | GTGCAGGGTCCGAGG |
| U6 | CTCGCTTCGGCAGCACA | AACGCTTCACGAATTTGCGT |
| β-actin | GTCTGCCTTGGTAGTGGATAATG | TCGAGGACGCCCTATCATGG |
Western blotting analysis
The lung tissues were cut into small pieces and homogenized for 10 s for three times in ice-cold radioimmunoprecipitation assay (RIPA) lysis and extraction buffer (Thermo Fisher Scientific). After stewing for 15 min on ice, the tissues were centrifuged at 5000 rpm for 5 min at 4°C, and the supernatant was collected. Protein concentration was tested using bicinchoninic acid (BCA) protein assay kit (Abcam); 30-μg total protein was loaded and separated using 10% ExpressPlus™ polyacrylamide gel electrophoresis (PAGE; GenScript Corporation, Piscataway, NJ, USA) at 100 V. Proteins were transferred from gels to polyvinylidene fluoride (PVDF) membranes (Merck KGaA) at 100 V for 1 h. The membranes were blocked using 5% fat-free milk for 2 h at room temperature. The target protein was probed with the corresponding primary antibodies at 4°C overnight and incubated with secondary antibodies for 2 h at room temperature. The protein signal was developed using Pierce™ ECL Western blotting substrate (Thermo Fisher Scientific) and imaged using GelView 6000Plus Imaging Systems (Bltlux, China). The protein bands were quantified using the ImageJ software (Softonic, Spain). Primary antibodies used were as follows: tipe2 (ab110389, 1:1000 dilution) and β-actin (ab8227, 1:5000 dilution).
Enzyme-linked immunosorbent serologic assay (ELISA)
ELISA was used to measure the serum levels of IFN-γ, TNF-α, IL-2, IL-6, IL-1β, and TGF-β1. ELISA was performed using commercial kits (R&D Systems, Minneapolis, MN., USA) according to the manufacturer’s instruction.
Cell culture and transfection
Normal human bronchial epithelial (NHBE) cells and human embryonic kidney (HEK) 293T cells were obtained from Ningbo Mingzhou Biotechnology Co. Ltd. (Mingzhou, Zhejiang Province, China). NHBE cells were cultured in Bronchial Epithelial Cell Growth Medium (BEGMTM; Lonza G and T AG, Switzerland) supplemented with 10% fetal bovine serum (FBS; Invitrogen), 1% penicillin–streptomycin solution (100×; Procell Life Science & Technology, Wuhan, Hubei Province, China), and SingleQuots® kit (Lonza G and T AG) at 37 C with 5% CO2. In all, 293T cells were cultured in Dulbecco’s modified Eagle’s medium (DMEM; Invitrogen) supplemented with 10% FBS, and 1% penicillin–streptomycin solution at 37°C with 5% CO2.
Short hairpin of HOXA-AS2 (sh HOXA-AS2) and its negative control (sh HOXA-AS2-NC), Adeno-associated virus containing HOXA-S2 gene (AAV-HOXA-S2) and its negative control (AAV-NC), and miR-17-5p mimic and its negative control (NC-mimic) were purchased from Hanbio Biotechnology Co. Ltd. (Shanghai, China). They were dissolved into nano-pure water at a stock concentration of 10 µg/mL and mixed with lipofactamine 2000 (Invitrogen) to form transfection mixture (1 ng/µL) according to manufacturer’s instructions. Cells were cultured with FBS-free medium into a sterile six-well plate; 200-ng transfection mixture was added to each well at 37°C. After 8 h, FBS-free medium was replaced with normal culture medium for another 24 h.
Plasmid and luciferase reporter assay
The full sequence of HOXA-AS2 (HOXA-AS2-WT) and its mutant sequence (HOXA-AS2-MUT), or the full sequence of tipe2 (tipe2-WT) and its mutant sequence (tipe2-MUT) were cloned into the downstream of the firefly luciferase gene in psiCHECK2 (Promega, USA). The constructed psiCHECK2 vector was co-transfected with miR-17-5p mimic or NC-mimic into 293T cells using lipofectamine 2000. After cultured for 24 h, luciferase activity was performed using dual luciferase reporter assay Kit (Vazyme, Jiangsu Province, China).
Statistical analysis
All results were analyzed using GraphPad Prism 8.1.0 (GraphPad, San Diego, CA, USA). All data were performed as mean ± standard error of mean (SEM). Statistical differences between two groups were analyzed using Student’s t-test under non-normal conditions. Statistical differences among more than two groups were performed using one-way ANOVA, followed by the Bonferroni's post hoc tests. Normality of data was assessed using Shapiro–Wilk normality tests. Homogeneity of data was analyzed using Bartlett’s test. P ˂ 0.05 was considered as statistically significant.
Results
HOXA-AS2 was downregulated in lung tissues from CIH rat model
The expression of HOXA-AS2 was determined to investigate the potential role of HOXA-AS2 in the progression of CIH. In the lung tissues of CIH rat model, granulocyte and lymphocyte infiltration, edema, and bleeding were observed, and the thickness of alveolar wall and blood vessels was increased in alveolar chambers, compared to that from sham group (Figure 1A). PaO2 level was reduced and PaCO2 was increased in CIH rat model, compared to that in sham rats (Figures 1B and C). The relative expression of HOXA-AS2 was downregulated in CIH rat model, compared to that in sham rats (Figure 1D).
Figure 1 HOXA-AS2 was downregulated in lung tissues from CIH rat model. (A) H & E staining results demonstrated lung tissue injury in CIH rat model. (B) PaO2 was reduced in CIH rats. (C) PaCO2 was elevated in CIH rats. (D) The expression of HOXA-AS2 was downregulated in CIH rats. **P ˂ 0.01 versus sham; ***P ˂ 0.005 versus sham. CIH: chronic intermittent hypoxia; HOXA-AS2: HOXA cluster antisense RNA 2; H & E staining: hematoxylin and eosin staining; PaO2: arterial partial pressure of oxygen; PaCO2: arterial partial pressure of carbon dioxide.
HOXA-AS2 reduced lung injury in CIH rat model
Overexpression of HOXA-AS2 was established by transfection of AVV-HOXA-AS2 to evaluate the involvement of HOXA-AS2 in lung injury in CIH rat model. The expression of HOXA-AS2 was increased in both sham and CIH rats after transfection of AVV-HOXA-AS2 (Figure 2A). The overexpression of HOXA-AS2 relieved CIH-induced lung tissue injury in rats (Figure 2B). The number of TUNEL-positive cells was also reduced after being transfected with AVV-HOXA-AS2 in CIH rats (Figure 2C). Cell apoptosis was significantly induced in CIH rats (Figure 2D). Although the overexpression of HOXA-AS2 caused no significant alterations in cell apoptosis in sham rats, cell apoptosis in CIH group was reduced (Figure 2D). The overexpression of HOXA-AS2 influenced no significant alterations in PaO2 and PaCO2 levels in sham rats, but significantly increased PaO2 and decreased PaCO2 levels in CIH rats (Figures 2E and F). These data demonstrated that HOXA-AS2 reduced both lung injury and cell apoptosis in CIH rats.
Figure 2 HOXA-AS2 reduced lung injury in CIH rat model. (A) AAV-HOXA-AS2 transfection increased the expression of HOXA-AS2 in CIH rats. (B) AAV-HOXA-AS2 ameliorated lung injury in CIH rats. (C) AAV-HOXA-AS2 reduced the number of TUMEL-positive cells in CIH rats. **P ˂ 0.01 versus sham + AVV-NC; ***P ˂ 0.005 versus sham + AVV-NC; #P ˂ 0.05 versus sham + AVV-HOXA-AS2; ###P ˂ 0.005 versus sham + AVV-HOXA-AS2; &P ˂ 0.05 versus CIH + AVV-NC; &&P ˂ 0.01 versus CIH + AVV-NC; &&&P ˂ 0.005 versus CIH + AVV-NC. AVV-HOXA-AS2: adeno-associated virus containing HOXA-S2 gene; AVV-NC: negative control of AVV-HOXA-AS2.
HOXA-AS2 reduced inflammatory response in CIH rat model
T cell differentiation and cytokine production were determined using flow cytometry to investigate the role of HOXA-AS2 to inflammatory response in CIH rats. The number of CD4+IFNγ+ T cells was elevated in the lung tissues of CIH rats, compared to that of sham rats, while this elevation was ameliorated by the overexpression of HOXA-AS2 (Figure 3A). The number of CD4+TGF-β1+ T cells was lower in CIH rats than that in sham rats, and this reduction was increased in CIH rats injected with HOXA-AS2 plasmid (Figure 3B). Serum levels of IFNγ and IL-2 were increased, and serum TGF-β1 was reduced in CIH rats, and these alterations were reversed by the overexpression of HOXA-AS2 (Figure 3C). Serums levels of TNF-α, IL-6, and IL-1β were significantly stimulated in CIH rats, and these enhancements were repressed by the overexpression of HOXA-AS2 (Figure 3D). It is deduced that HOXA-AS2 could reduce inflammatory response in CIH rats.
Figure 3 HOXA-AS2 reduced inflammatory response in CIH rat model. (A) AVV-HOXA-AS2 inhibited the induction of CD4+ IFNγ+ T cells in CIH rats. (B) AVV-HOXA-AS2 reversed the suppression of CD4+TGF-β1+ T cells in CIH rats. (C) AVV-HOXA-AS2 inhibited the induction of IFNγ and IL-2, and the reduction of TGF-β1 in CIH rats. (D) AVV-HOXA-AS2 ameliorated the induction of TNF-α, IL-6, and IL-1β in CIH rats. ***P ˂ 0.005 versus sham + AVV-NC; ##P ˂ 0.01 versus sham + AVV-HOXA-AS2; ###P ˂ 0.005 versus sham + AVV-HOXA-AS2; &&&P ˂ 0.005 versus CIH + AVV-NC. IFNγ: interferon gamma; TGF-β1: transforming growth factor beta1; IL-2: interleukin-2; IL-6: interleukin-6; IL-1β: interleukin-1β; TNF-α: tumor necrosis factor alpha.
HOXA-AS2 regulated the expressions of miR-17-5p and tipe2 in CIH rat model
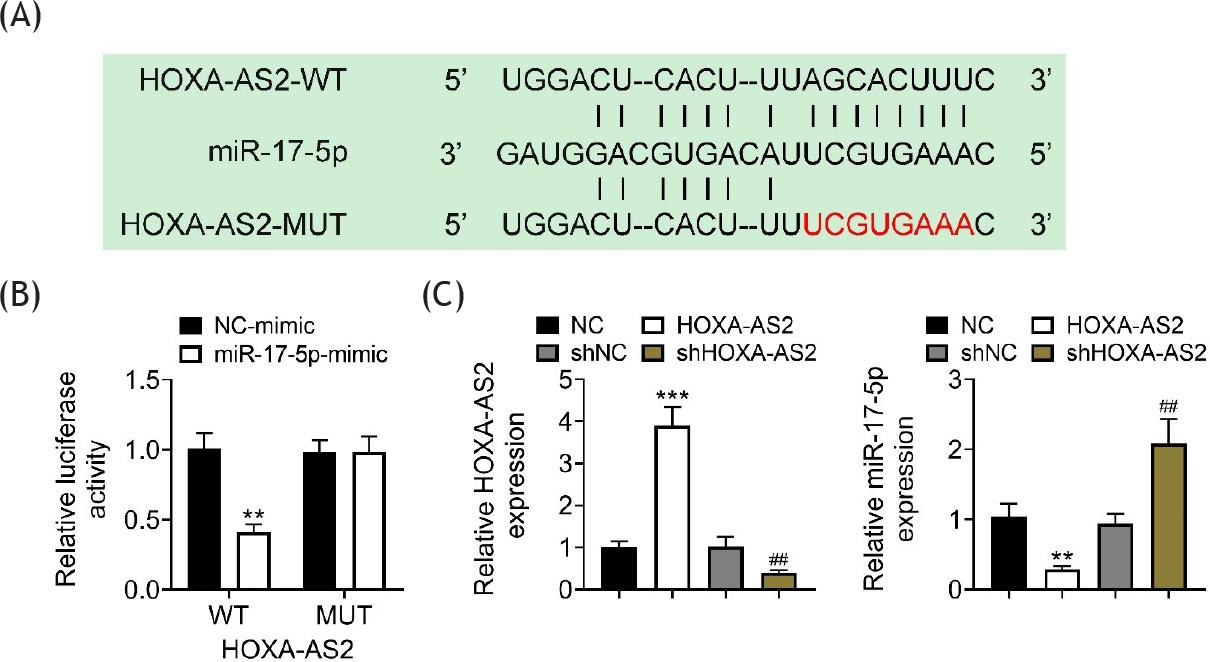
In order to confirm the relationship between HOXA-AS2, miR-17-5p, and tipe2, expressions of miR-17-5p and tipe2 were determined after the overexpression of HOXA-AS2 in 293T cells. The overexpression of HOXA-AS2 suppressed the mRNA expression of miR-17-5p but increased the mRNA expression of tipe2 in both sham and CIH rats (Figure 4A). The protein expression of tipe2 was upregulated by the overexpression of HOXA-AS2 in both sham and CIH rats (Figure 4B). The predictive results from the Encyclopedia of RNA Interactomes (ENCORI; https://starbase.sysu.edu.cn/) demonstrated that there was a complementary sequence between HOXA-AS2 and miR-17-5p (Figure 5A). Luciferase activity was decreased in cells co-transfected with HOXA-AS2-WT and miR-17-5p mimic, compared to that with HOXA-AS2-WT and NC mimic (Figure 5B). The expression of HOXA-AS2 was increased by HOXA-AS2 plasmid and deceased by sh HOXA-AS2 (Figure 5C), while the expression of miR-17-5p was downregulated by HOXA-AS2 and upregulated by sh HOXA-AS2 (Figure 5D) in NHBE cells. These data supported the hypothesis that miR-17-5p was the direct target of HOXA-AS2.
Figure 4 HOXA-AS2 regulated the expressions of miR-17-5p and tipe2 in CIH rat model. (A) AVV-HOXA-AS2 downregulated the expression of miR-17-5p and upregulated the mRNA of tipe2 in CIH rats. (B) AVV-HOXA-AS2 upregulated the protein expression of tipe2 in CIH rats. **P ˂ 0.01 versus sham + AVV-NC; ***P ˂ 0.005 versus sham + AVV-NC; ###P ˂ 0.005 versus sham + AVV-HOXA-AS2; &P ˂ 0.05 versus CIH + AVV-NC; &&&P ˂ 0.005 versus CIH + AVV-NC. Tipe2: tumor necrosis factor-alpha-induced protein 8-like 2.
Figure 5 miR-17-5p was the direct target of HOXA-AS2. (A) There was a complementary sequence between HOXA-AS2 and miR-17-5p. (B) Luciferase activity was decreased in 293T cells co-transfected with HOXA-AS2-WT and miR-17-5p mimic, compared to that with HOXA-AS2-WT and NC mimic. (C) Repression of HOXA-AS2 reduced the expression of HOXA-AS2 but increased the expression of miR-17-5p normal human bronchial epithelial (NHBE) cells. **P ˂ 0.01 versus NC; ***P ˂ 0.005 versus NC; ##P ˂ 0.01 versus shNC. HOXA-AS2-WT: psiCHECK2 contained full sequence of HOXA-AS2; HOXA-AS2-MUT: psiCHECK2 contained mutant sequence of HOXA-AS2; HOXA-AS2: adeno-associated virus containing HOXA-S2 gene; NC: negative control of HOXA-AS2; shHOXA-AS2: short hairpin of HOXA-AS2; shNC: negative control of shHOXA-AS2.
HOXA-AS2 positively regulated tipe2 by sponging miR-17-5p
ENCORI and luciferase assay were used to confirm the direct target of HOXA-AS2 and miR-17-5p. The predictive results from ENCORI showed that there was a complementary sequence between miR-17-5p and 3’-UTR of tipe2 (Figure 6A). Luciferase activity was reduced in 293T cells co-transfected with miR-17-5p mimic and tipe2-WT, compared to that with NC mimic and tipe2-WT (Figure 6B). The expression of miR-17-5p was elicited by miR-17-5p mimic and suppressed by miR-17-5p inhibitor, while the mRNA expression of tipe2 was decreased by miR-17-5p mimic and increased by miR-17-5p inhibitor in NHBE cells (Figure 6C). The protein expression of tipe2 was downregulated by miR-17-5p mimic and upregulated by miR-17-5p inhibitor in NHBE cells (Figure 6D). The mRNA and protein expressions of tipe2 were overexpressed by upregulation of HOXA-AS2 and ameliorated after cotransfection of HOXA-AS2 and miR-17-5p mimic in NHBE cells (Figures 6E and F).
Figure 6 Tipe2 was the direct target of miR-17-5p. (A) There was a complementary sequence between 3’-UTR of tipe2 and miR-17-5p. (B) Luciferase activity was decreased in 293T cells co-transfected with tipe2-WT and miR-17-5p mimic, compared to that with tipe2-WT and NC mimic. (C) miR-17-5p mimic increased the expression of miR-17-5p and decreased the expression of tipe2 mRNA in NHBE cells. (D) The protein expression was decreased by miR-17-5p mimic and increased by miR-17-5p inhibitor in NHBE cells. (E) miR-17-5p mimic reversed HOXA-AS2-induced upregulation of tipe2 mRNA in NHBE cells. (F) miR-17-5p mimic reversed HOXA-AS2-induced upregulation of tipe2 protein in NHBE cells. *P ˂ 0.05 versus NC mimic; **P ˂ 0.01 versus NC mimic; ***P ˂ 0.005 versus NC mimic; ##P ˂ 0.01 versus NC inhibitor or HOXA-AS2; ###P ˂ 0.005 versus NC inhibitor or HOXA-AS2. NC: negative control.
Discussion
Chronic intermittent hypoxia is the main feature of obstructive sleep apnea (OSA).16 CIH could induce lung injury, causing lung fibrosis and lung cancer.23,24 Therefore, it is necessary to prevent CIH-induced lung injury in the early stage. In this study, lung tissue injury was observed in CIH rat model accompanying the downregulation of HOXA-AS2. The overexpression of HOXA-AS2-inhibited cell apoptosis increased the number of anti-inflammatory T lymphocytes and enhanced serum levels of anti-inflammatory molecules, thereby ameliorating lung tissue injury. Further experiments proved that HOXA-AS2 acted as an anti-inflammatory regulator through sponging miR-17-5p to upregulate the expression of tipe2, providing a new mechanism for CIH, and building up a new insight on the prevention and treatment for CIH.
Consistent with the previous studies,25,26 lung injury and increased levels of PaO2 were observed in the present study also. The overexpression of HOXA-AS2 reversed CIH-induced lung injury, demonstrating a protective effect on CIH. T cell differentiation was a key procedure to defense the inflammatory injury.27,28 During CIH, CD4+ T cell differentiated into pro-inflammatory subtype, CD4+IFNγ+ T cells, but not anti-inflammatory subtype, CD4+TGF-β1+ T cells, which were ameliorated by the overexpression of HOXA-AS2; this demonstrated that HOXA-AS2 played an anti-inflammatory role in CIH. Upregulation of pro--inflammatory molecules and downregulation of anti--inflammatory molecules were also observed in the present study, and HOXA-AS2 reversed inflammatory response in the lung tissues of CIH rat model. This was the first study to describe that HOXA-AS2 inhibited lung inflammatory response in CIH rat model. The anti-inflammatory effects of HOXA-AS2 were consistent with the conclusions of previous findings.17,29
LncRNAs can regulate transcriptional process through two different ways: (1) recruiting regulatory factors to modulate the expression of neighboring genes; and (2) acting as competing endogenous RNAs (ceRNAs) to upregulate target proteins through directly sponging miRNAs.10 In this study, ENCORI prediction and luciferase assay proved that miR-17-5p was the target miRNA of HOXA-AS2. It is well known that miRNAs suppressed translational process by binding to 3’-untranslated regions (3’-UTR) of target mRNA,12 and some studies have reported that miRNA played a crucial role in lung diseases.30,31 ENCORI predictive assay demonstrated that there was a complementary sequence between miR-17-5p and 3’UTR of tipe2, suggesting that tipe2 was the direct target of miR-17-5p. Results of luciferase assay, qRT-PCR, and Western blotting analysis further confirmed this conclusion.
Tipe2 is an anti-inflammatory regulator in many biological processes.20 In the lungs, tipe2 can regulate inflammation and maintain immune homeostasis in asthma.32 The functional role of tipe2 in CIH-induced lung inflammation has been described for the first time. The mRNA and protein expression of tipe2 were elevated by HOXA-AS2. The expression level of tipe2 was synchronized with HOXA-AS2 but negatively correlated with miR-17-5p. Therefore, it can be summarized that HOXA-AS2 exerted its anti-inflammatory effect through HOXA-AS2/miR-17-5p/tipe2 axis, demonstrating a new mechanism of CIH and providing new therapeutic targets for CIH.
Conclusion
This study manifested that HOXA-AS2 acted as an anti--inflammatory regulator and protected lung tissue injury from CIH in a rat model. By directly targeting miR-17-5p, HOXA-AS2 positively regulated the expression of tipe2, which played an anti-inflammatory role in CIH, thus preventing lung injury from CIH. The findings of this study provided new understanding and therapeutic targets for CIH to facilitate a new drug discovery for CIH treatment.
Availability of Data and Materials
All data generated and analyzed during this study are included in this article.
Competing interests
The authors stated that there was no conflict of interest to declare.
Author Contributions
Kun Gao and Aiai Lv did conceptualization, methodology, and writing of the original draft of this paper. Formal analysis, resources, and investigation were done by Qiang Zhang. Formal analysis, visualization, and data curation were performed by Yanzhong Li. Project administration, supervision, and validation was executed by Zhiyong Yue. Validation, supervision, and writing, including review and editing, was done by Shuai Xu. All authors read and approved the final manuscript.
REFERENCES
1. Brito J, Siques P, Pena E. Long-term chronic intermittent hypoxia: A particular form of chronic high-altitude pulmonary hypertension. Pulm Circ. 2020;10(1 Suppl):5–12. 10.1177/2045894020934625
2. Song D, Fang G, Greenberg H, Liu SF. Chronic intermittent hypoxia exposure-induced atherosclerosis: A brief review. Immunol Res. 2015;63(1–3):121–30. 10.1007/s12026-015-8703-8
3. Zhang X, Rui L, Wang M, Lian H, Cai L. Sinomenine attenuates chronic intermittent hypoxia-induced lung injury by inhibiting inflammation and oxidative stress. Med Sci Monit. 2018;24:1574–80. 10.12659/msm.906577
4. Kiernan EA, Smith SM, Mitchell GS, Watters JJ. Mechanisms of microglial activation in models of inflammation and hypoxia: Implications for chronic intermittent hypoxia. J Physiol. 2016;594(6):1563–77. 10.1113/JP271502
5. Liew FY. Induction and regulation of CD4+ T cell subsets. Ciba Found Symp. 1994;187:170–5; Discussion, 176–8. 10.1002/9780470514672.ch11
6. Becker C, Fantini MC, Neurath MF. TGF-beta as a T cell regulator in colitis and colon cancer. Cytokine Growth Factor Rev. 2006;17(1–2):97–106. 10.1016/j.cytogfr.2005.09.004
7. Rahmati M, Keshvari M, Xie W, Yang G, Jin H, Li H, et al. Resistance training and Urtica dioica increase neurotrophin levels and improve cognitive function by increasing age in the hippocampus of rats. Biomed Pharmacother. 2022;153:113306. 10.1016/j.biopha.2022.113306
8. Rahmati M, Keshvari M, Mirnasouri R, Chehelcheraghi F. Exercise and Urtica dioica extract ameliorate hippocampal insulin signaling, oxidative stress, neuroinflammation, and cognitive function in STZ-induced diabetic rats. Biomed Pharmacother. 2021;139:111577. 10.1016/j.biopha.2021.111577
9. Keshvari M, Rahmati M, Mirnasouri R, Chehelcheraghi F. Effects of endurance exercise and Urtica dioica on the functional, histological and molecular aspects of the hippocampus in STZ-induced diabetic rats. J Ethnopharmacol. 2020;256:112801. 10.1016/j.jep.2020.112801.
10. Kopp F, Mendell JT. Functional classification and experimental dissection of long noncoding RNAs. Cell. 2018;172(3):393–407. 10.1016/j.cell.2018.01.011
11. Hu C, Li J, Du Y, Li J, Yang Y, Jia Y, et al. Impact of chronic intermittent hypoxia on the long non-coding RNA and mRNA expression profiles in myocardial infarction. J Cell Mol Med. 2021;25(1):421–33. 10.1111/jcmm.16097
12. Zhang Y, Sun X, Icli B, Feinberg MW. Emerging roles for microRNAs in diabetic microvascular disease: Novel targets for therapy. Endocr Rev. 2017;38(2):145–68. 10.1210/er.2016-1122
13. Cai Y, Zhang Y, Chen H, Sun XH, Zhang P, Zhang L, et al. MicroRNA-17-3p suppresses NF-κB-mediated endothelial inflammation by targeting NIK and IKKβ binding protein. Acta Pharmacol Sin. 2021;42(12):2046–57. 10.1038/s41401-021-00611-w
14. Alashkar Alhamwe B, Miethe S, Pogge von Strandmann E, Potaczek DP, Garn H. Epigenetic regulation of airway epithelium immune functions in asthma. Front Immunol. 2020;11:1747. 10.3389/fimmu.2020.01747
15. Boateng E, Krauss-Etschmann S. miRNAs in lung development and diseases. Int J Mol Sci. 2020;21(8):2765. 10.3390/ijms21082765..
16. Wang X, Li Z, Du Y, Xing Y, Guo Y, Zhang Y, et al. lncRNA Mirt1: A critical regulatory factor in chronic intermittent hypoxia exaggerated post-MI cardiac remodeling. Front Genet. 2022;13:818823. 10.3389/fgene.2022.818823
17. Zhu X, Liu Y, Yu J, Du J, Guo R, Feng Y, et al. LncRNA HOXA-AS2 represses endothelium inflammation by regulating the activity of NF-κB signaling. Atherosclerosis. 2019;281:38–46. 10.1016/j.atherosclerosis.2018.12.012
18. Li X, Yu HM. Overexpression of HOXA-AS2 inhibits inflammation and apoptosis in podocytes via sponging miRNA-302b-3p to upregulate TIMP3. Eur Rev Med Pharmacol Sci. 2020;24(9):4963–70. 10.26355/eurrev_202005_21187
19. Gu Z, Cui X, Sun P, Wang X. Regulatory roles of tumor necrosis factor-α-induced protein 8 like-protein 2 in inflammation, immunity and cancers: A review. Cancer Manag Res. 2020;12:12735–46. 10.2147/CMAR.S283877
20. Wu X, Kong Q, Zhan L, Qiu Z, Huang Q, Song X. TIPE2 ameliorates lipopolysaccharide-induced apoptosis and inflammation in acute lung injury. Inflamm Res. 2019;68(11):981–92. 10.1007/s00011-019-01280-6
21. Ruan Q, Wang P, Wang T, Qi J, Wei M, Wang S, et al. MicroRNA-21 regulates T-cell apoptosis by directly targeting the tumor suppressor gene tipe2. Cell Death Dis. 2014;5(2):e1095. 10.1038/cddis.2014.47
22. National Research Council (US) Committee for the Update of the Guide for the Care and Use of Laboratory Animals. Guide for the Care and Use of Laboratory Animals, 8th ed. The National Academies Collection: Reports funded by National Institutes of Health. Washington DC: National Academies Press; 2011.
23. Braun RK, Broytman O, Braun FM, Brinkman JA, Clithero A, Modi D, et al. Chronic intermittent hypoxia worsens bleomycin-induced lung fibrosis in rats. Respir Physiol Neurobiol. 2018;256:97–108. 10.1016/j.resp.2017.04.010
24. Gu X, Zhang J, Shi Y, Shen H, Li Y, Chen Y, et al. ESM1/HIF-1α pathway modulates chronic intermittent hypoxia-induced non-small-cell lung cancer proliferation, stemness and epithelial-mesenchymal transition. Oncol Rep. 2021;45(3):1226–34. 10.3892/or.2020.7913
25. Ge L, Ming T, Hou J, Yan J, Wang Q, Qiao F. Role of NF-κB p65 and p38 MAPK in lung injury in rats suffered chronic intermittent hypoxia. Zhong Nan Da Xue Xue Bao Yi Xue Ban. 2015;40(12):1313–9. 10.11817/j.issn.1672-7347.2015.12.005
26. Hou Y, Xu N, Li S, Zhang N, Ren W, Hua Z, et al. Mechanism of SMND-309 against lung injury induced by chronic intermittent hypoxia. Int Immunopharmacol. 2022;105:108576. 10.1016/j.intimp.2022.108576
27. Potaczek DP, Harb H, Michel S, Alhamwe BA, Renz H, Tost J. Epigenetics and allergy: From basic mechanisms to clinical applications. Epigenomics. 2017;9(4):539–71. 10.2217/epi-2016-0162
28. Suárez-Álvarez B, Baragaño Raneros A, Ortega F, López-Larrea C. Epigenetic modulation of the immune function: A potential target for tolerance. Epigenetics. 2013;8(7):694–702. 10.4161/epi.25201
29. Wu H, Wang J, Ma Z. Long noncoding RNA HOXA-AS2 mediates microRNA-106b-5p to repress sepsis-engendered acute kidney injury. J Biochem Mol Toxicol. 2020;34(4):e22453. 10.1002/jbt.22453
30. Alashkar Alhamwe B, Potaczek DP, Miethe S, Alhamdan F, Hintz L, Magomedov A, et al. Extracellular vesicles and asthma—More than just a co-existence. Int J Mol Sci. 2021;22(9):4984. 10.3390/ijms22094984
31. Cañas JA, Rodrigo-Muñoz JM, Sastre B, Gil-Martinez M, Redondo N, Del Pozo V. MicroRNAs as potential regulators of immune response networks in asthma and chronic obstructive pulmonary disease. Front Immunol. 2020;11:608666. 10.3389/fimmu.2020.608666
32. Shi B, Hao Y, Li W, Dong H, Xu M, Gao P. The enigmatic role of tipe2 in asthma. Am J Physiol Lung Cell Mol Physiol. 2020;319(1):L163–72. 10.1152/ajplung.00069.2020