
Download
ORIGINAL ARTICLE
Vitamin A–regulated ciliated cells promote airway epithelium repair in an asthma mouse model
Wen Tana, b, Jilei Lina, b, Yaping Wanga, Li Yana, b, Linyan Yinga, b, Jihong Daia, b, Zhou Fua, b, Jingyue Liua, b*
aNational Clinical Research Center for Child Health and Disorders, Ministry of Education Key Laboratory of Child Development and Disorders, Chongqing Key Laboratory of Pediatrics, Children’s Hospital of Chongqing Medical University, Chongqing, China
bDepartment of Respiratory Medicine, Children’s Hospital of Chongqing Medical University, Chongqing, China
Abstract
Background: Asthma is a chronic inflammatory airway disease that causes damage to and exfoliation of the airway epithelium. The continuous damage to the airway epithelium in asthma cannot be repaired quickly and generates irreversible damage, repeated attacks, and aggravation. Vitamin A (VA) has multifarious biological functions that include maintaining membrane stability and integrity of the structure and function of epithelial cells. Our research explored the role of VA in repairing the airway epithelium and provided a novel treatment strategy for asthma.
Methods: A mouse asthma model was established by house dust mite (HDM) and treated with VA by gavage. Human bronchial epithelial (16HBE) cells were treated with HDM and all-trans retinoic acid (ATRA) in vitro. We analyzed the mRNA and protein expression of characteristic markers, such as acetyl-α-tubulin (Ac-TUB) and FOXJ1 in ciliated cells and MUC5AC in secretory cells, mucus secretion, airway inflammation, the morphology of cilia, and the integrity of the airway epithelium.
Results: Findings showed destruction of airway epithelial integrity, damaged cilia, high mucus secretion, increased MUC5AC expression, and decreased Ac-TUB and FOXJ1 expression in asthmatic mice. The VA intervention reversed the effect on Ac-TUB and FOXJ1 and promoted ciliated cells to repair the damage and maintain airway epithelial integrity. In 16HBE cells, we could confirm that ATRA promoted the expression of Ac-TUB and FOXJ1.
Conclusion: These results demonstrated that VA-regulated ciliated cells to repair the damaged airway epithelium caused by asthma and maintain airway epithelial integrity. VA intervention is a potential adjunct to conventional treatment for asthma.
Key words: airway epithelium, asthma, ciliated cell, vitamin A, treatment
*Corresponding author: Jingyue Liu, Department of Respiratory Medicine, Children’s Hospital of Chongqing Medical University, No. 136 Zhongshan 2nd Road, Yuzhong District, Chongqing 400014, China. Email address: [email protected]
Received 14 June 2022; Accepted 13 October 2022; Available online 1 January 2023
Copyright: Tan W, et al.
License: This open access article is licensed under Creative Commons Attribution 4.0 International (CC BY 4.0). http://creativecommons.org/licenses/by/4.0/
Introduction
Asthma is a chronic inflammatory disease that seriously threatens human health; the number of patients worldwide has exceeded 300 million. Asthma is a common chronic airway disease in children, with death rates ranging from 0 to 0.7 per 100,000 people.1 Although first-line treatment with glucocorticoids can control airway inflammation, this does not fundamentally reduce morbidity and mortality. Our group previously confirmed that glucocorticoid treatment partially inhibited repair of the airway epithelium after injury.2 Then, recurrent attacks and aggravation of asthma may be related to persistent airway epithelial injury.
The airway epithelium is a barrier of interaction between the internal and external environment, and the integrity of its structure and function is necessary to maintain the local airway microenvironment. Normal airway epithelial cells are mainly composed of ciliated cells, basal cells, and secretory cells, which account for 50–90%, 6–30%, and 5–30% of the cells in the airway, respectively.3 Ciliated cells, as the main component of mucociliary clearance system, can effectively remove invasive pathogens, microorganisms, and particles. In general, the injured airway epithelium can repair itself to maintain structural integrity.4 The airway epithelium plays a key role in the occurrence and development of asthma.5,6 In one study, endobronchial biopsy in asthma patients revealed varying degrees of airway epithelial damage.7 If the airway barrier sustains long-term damage and cannot be repaired quickly, there occur ciliated cell damage and exfoliation, mucous cell metaplasia, high mucus secretion, inflammatory cell infiltration, and airway remodeling.8,9 Ciliated cell injury and increased mucus secretion lead to a series of abnormal behaviors. Destruction of the airway epithelial structure increases the frequency of attacks and severity of asthma. Therefore, the identification of effective measures to repair the airway epithelium is of great clinical significance.
Vitamin A (VA) is a class of compounds with retinol bioactivity and regulates multifarious biological processes such as cell growth, differentiation, and morphogenesis. VA is also a necessary factor for maintaining the stability of the cell membrane structure.10 VA deficiency affects epithelial cells in various tissues and causes mucosal barrier function damage, resulting in corresponding clinical manifestations. The occurrence and development of asthma is closely related to VA status. When VA is deficient, the ability to fight off infection decreases, airway repair weakens, and airway responsiveness increases, which lead to repeated asthma attacks. Data showed that the level of VA in children with asthma was lower than that in normal children.11 Additionally, VA intake is associated with improved lung function, as well as a lower risk of fixed airflow limitation and incident asthma.12,13 Furthermore, VA maintains the integrity of the structure and function of epithelial cells, which may play an important role in the repair of damaged airway epithelium caused in asthma. The reduction of VA in patients with asthma may hinder the repair process of the airway epithelium, resulting in repeated attacks and aggravation, generating irreversible damage and promoting airway remodeling. Therefore, we speculate that VA supplementation can reduce the damage to the airway epithelium, promote the repair of airway epithelium, and restore the barrier function of the airway epithelium. This study aimed to investigate the repair effect of VA on the damaged airway epithelium caused by asthma and to provide a potential new treatment strategy for treating patients with asthma.
Materials and Methods
Cell culture
Human bronchial epithelial (16HBE) cells were provided by the Respiratory Research Laboratory, Ministry of Education Key Laboratory of Child Development and Disorders, Children’s Hospital of Chongqing Medical University, China. 16HBE cells were maintained in Dulbecco’s modified Eagle medium (DMEM, Gibco, New York, USA) supplemented with 10% fetal bovine serum (BI, Israel) and were incubated at 37°C in a 5% CO2 atmosphere. The cells were treated with house dust mite (HDM, Greer, Los Angeles, USA) (25 μg/mL) and all-trans retinoic acid (ATRA, Sigma-Aldrich, St. Louis, USA) (2 μM) and then collected for testing.
Animals
Six to eight-weeks-old female C57BL/6 mice were purchased from the Experimental Animal Center of Chongqing Medical University, China. The mice were raised under specific pathogen-free conditions. The study was approved by the Ethics Committee of the Chongqing Medical University (Reference number: CHCMU-IACUC20200424005). All procedures were fully in compliance with the strict guidelines of the Chongqing Medical University Policy on Animal Care and Use. The three experimental groups were the control group, asthma group, and asthma + VA group (VA-treated asthma group).
Establishment of the asthma mouse model
Mice were sensitized intranasally with HDM (20 μg) on Days 0 and 14 and then challenged by intranasal HDM (20 μg) on Days 21, 23, 25, 27, and 29.14 VA (44 IU/g) was administered by gavage every 7 days (Days 0, 7, 14, 21, 28). Within 24 h of the last challenge, mice were euthanized for analysis.
Determination of airway hyperresponsiveness (AHR)
AHR was detected within 24 h of the last HDM treatment. Lung function index was measured using whole-body plethysmograph (EMKA Technologies, Paris, France). The mice were exposed to gradient concentrations (0, 3.125, 6.25, 12.5, 25, and 50 mg/mL) of acetyl-β-methylcholine chloride (Sigma-Aldrich, St. Louis, USA). The index of airflow obstruction was recorded according to previously described methods.14,15
Bronchoalveolar lavage fluid (BALF) assays
The tracheae of the mice were cannulated, and the left bronchus was clamped. Then, BALF was collected by three injections of 0.5 mL sterile phosphate-buffered saline into the right lung. After BALF was centrifuged, the number of collected total cells was counted using a cell counter. The cells were stained with Wright–Giemsa stain (Jiancheng Techno Co., Nanjing, China), 200 cells were counted from each sample, and the number of eosinophils was calculated.
Detection of total IgE
Blood samples were obtained by extracting eyeball blood under anesthesia and centrifuged at 2500 rpm for 15 min at room temperature. The supernatants were collected and used to detect total IgE levels using a mouse IgE ELISA kit (NeoBioscience, Shenzhen, China) according to the manufacturer’s protocol.
Lung histopathology
After lung lavage, the paraffin-embedded left lungs were sliced into 4-μm-thick sections and were stained with hematoxylin and eosin (H&E) for analysis. The severity of inflammation in the lung tissues was evaluated on a 0–3 scale defined as: 0, no inflammation; 1, mild inflammation with punctate infiltration of inflammatory cells in the bronchial or vascular wall and alveolar septum; 2, moderate inflammation with patchy infiltration of inflammatory cells in the bronchial or vascular walls and alveolar septum, which involved less than 1/3 of the lung cross--sectional area; 3, severe inflammation with diffuse infiltration of inflammatory cells in the bronchial or vascular walls and alveolar septum, which involved between 1/3 and 2/3 of the lung cross-sectional area.16 The histological assessments were performed by two independent blinded observers.
Vitamin A concentration measurement
Serum retinol concentrations were measured by high-performance liquid chromatography (HPLC).17 Serum (200 μL) was deproteinized with absolute alcohol, and the retinol was extracted with hexane and evaporated to dryness with nitrogen gas. The mobile phase was a double-distilled water mixture (96:4). The mobile phase mixture (100 µL) was added to dilute the precipitate, and a portion (20 µL) of the sample was injected into a bottle that was inserted into a C18 column (5 μm, 4.6 × 150 mm, Waters, Milford, USA) on the HPLC apparatus (Agilent, Palo Alto, USA). The retinol concentrations were determined by spectrophotometry at 315 nm.
Glycogen staining
Lung tissue slices were obtained as described above, dewaxed with xylene and dehydrated using graded ethanol. Slices were stained using Glycogen Periodic Acid-Schiff Staining Kit (LEAGENE, Beijing, China) following the manufacturer's instructions. Then, the severity of mucus secretion was evaluated on a 0–4 scale: 0, no PAS-positive cells; 1, ˂ 10% PAS-positive cells; 2, 10–25% PAS-positive cells; 3, 25–50% PAS-positive cells; and 4, ˃50% PAS-positive cells within a certain visual field.18 The histological parameters of all samples were independently analyzed by two blinded observers.
Scanning electron microscope (SEM)
Tracheae were cut lengthwise, and mucus was removed by ultrasound. Samples were fixed with 2.5% glutaraldehyde, dehydrated in a graded series of alcohol, and dried with tertiary butanol. Then, the dried samples were subjected to ion-spattering coating and observed using a SEM (SU8010, Hitachi, Tokyo, Japan).
Quantitative real-time PCR (qRT-PCR)
Total RNA was extracted from 16HBE cells and mouse lung tissues using the Simply P Total RNA Extraction Kit (BioFlux, Hangzhou, China) and reverse-transcribed into cDNA with the EvoScript Universal cDNA Master (Roche, Mannheim, Germany). The reaction was performed using RealMasterMix SYBR Green (TakaRa, Shiga, Japan). The primer sequences of Ac-TUB/FOXJ1/MUC5AC/GAPDH are shown in Table 1.
Table 1 The primers for quantitative real-time PCR.
| Genes Forward primer Reverse primer |
|---|
| Human Ac-TUB GAGGAGATGACTCCTTCAACACC TGATGAGCTGCTCAGGGTGGAA Human FOXJ1 TGGATCACGGACAACTTCTGCTA CACTTGTTCAGAGACAGGTTGTG Human MUC5AC CAGCCACGTCCCCTTCAATA ACCGCATTTGGGCATCC Human GAPDH GTCTCCTCTGACTTCAACAGCG ACCACCCTGTTGCTGTAGCCAA Mouse Ac-TUB GGTGATGTGGTTCCCAAAGA GTGGGAGGCTGGTAGTTAATG Mouse FOXJ1 AGAGAGTGAGGGCAAGAGAC GCGGGCTTAGAGACCATTTC Mouse MUC5AC ACTGTTACTATGCGATGTGTAGCCA GAGGAAACACATTGCACCGA Mouse GAPDH GACATGCCGCCTGGAGAAAC AGCCCAGGATGCCCTTTAGT |
Western blotting (WB)
Proteins from 16HBE cells and mouse lung tissues were extracted using the Total Protein Extraction Kit (Keygen, Nanjing, China). Proteins were electrophoresed using sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) (Beyotime, Shanghai, China) and were transferred onto polyvinylidene fluoride membranes. Nonspecific binding was blocked using blocking buffer (NCM, Suzhou, China) for 20 min, and membranes were incubated with primary antibodies for acetyl-α-tubulin (Ac-TUB) (1:1000, D20G3, Cell Signaling, Boston, USA), FOXJ1 (1:250, 14-9965-82, Invitrogen, Carlsbad, USA), MUC5AC (mouse: 1:500, NBP2-15196, Novus, Colorado, USA; human: 1:500, 381811, Zen Bio, Chengdu, China), β-actin (1:10000, 700068, Zen Bio, Chengdu, China), and GAPDH (1:10000, 200306-7E4, Zen Bio, Chengdu, China) overnight at 4°C. After the blots were washed, they were immersed in HRP-linked anti-mouse or rabbit secondary antibodies (1:1000, A0216/A0208, Beyotime, Shanghai, China) for 1 h at room temperature. Finally, the blots were visualized using an Ultrasensitive ECL Western HRP Substrate (Zen Bio, Chengdu, China).
Confocal laser scanning
16HBE cells were cultured in 24-well plates, and the glass slides were placed at the bottom. Cells were fixed with 4% paraformaldehyde for 30 min. Slides were incubated with 0.1% TritonX-100 for 20 min, and then 5% bovine serum albumin was used to block the reaction for 30 min. The lung tissue slices were obtained as described above, and antigenic repair was performed with sodium citrate. Slices were incubated with Ac-TUB and MUC5AC antibodies (1:800/1:200) overnight at 4°C. After washing, they were incubated with Alexa Fluor 488–labeled Goat Anti-Mouse secondary antibody (1:500, Beyotime, Shanghai, China) and Cy3-labeled Goat Anti-Rabbit secondary antibody (1:500, Beyotime, Shanghai, China) at room temperature for 1 h. Finally, slices were dyed with DAPI for 15 min and examined by confocal laser scanning using a Leica laser confocal microscope (C2+ system, Nikon, Tokyo, Japan).
Statistical analysis
The data were analyzed using GraphPad Prism 7.0 (Graph-Pad, San Diego, CA, USA) via Student’s t test or two-way ANOVA. The data are expressed as mean ± standard deviation, and P ˂ 0.05 was considered significant.
Results
Identification of the HDM-induced mouse model of asthma
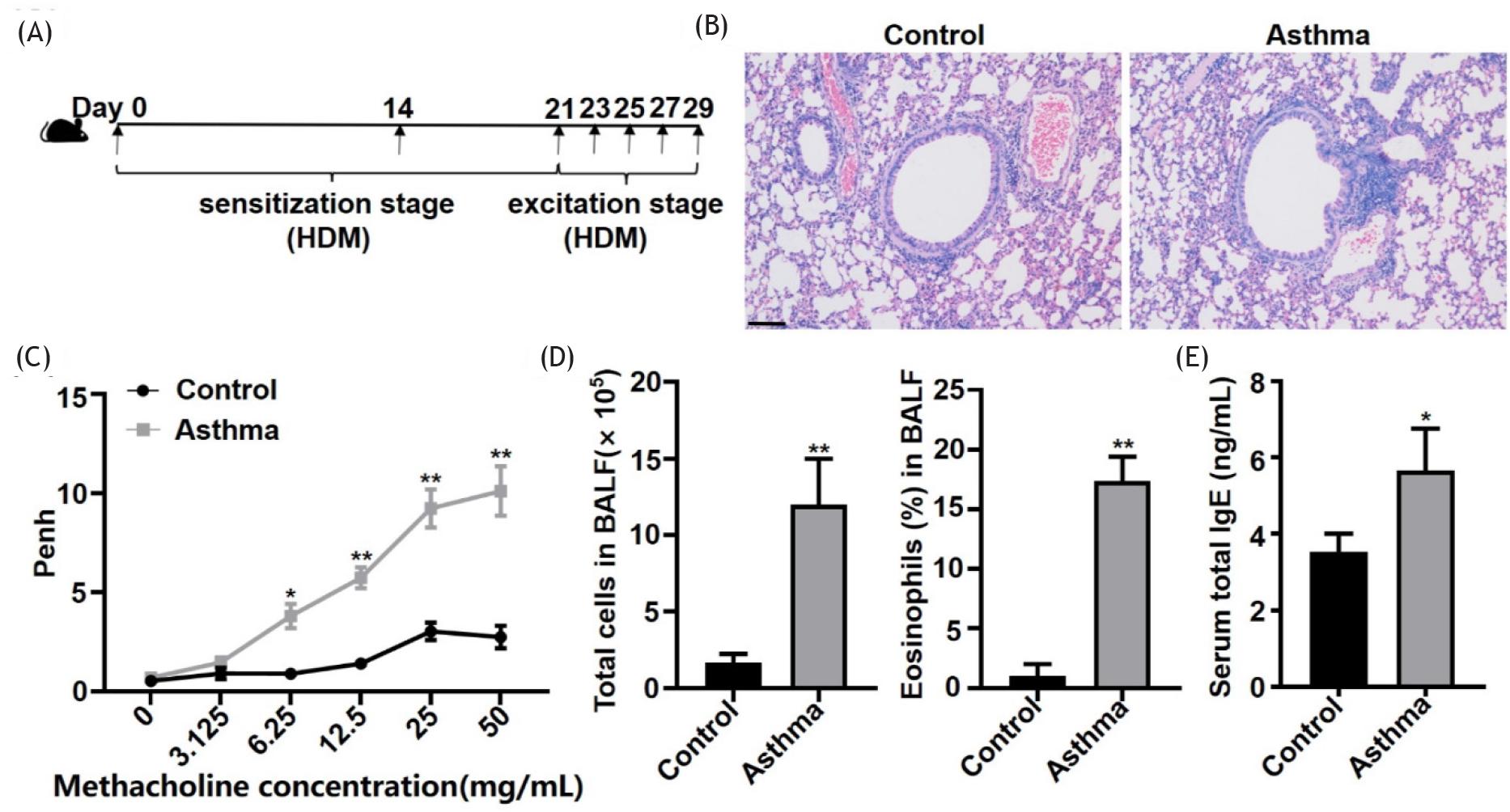
The C57BL/6 mouse model of asthma was established using HDM according to the schematic (Figure 1A). The mice were analyzed by H&E staining, Penh values, total cell and eosinophil count in the BALF, and serum total IgE. In asthmatic mice, the airway epithelium was injured, and morphology was incomplete. Inflammatory cells had infiltrated around the bronchia and vasculature (Figure 1B). Airflow obstruction began to occur in the asthma model at a concentration of 6.25 mg/mL methacholine and continued to exist up to a concentration of 50 mg/mL (Figure 1C). The number of total cells and eosinophils in the BALF and serum total IgE levels were significantly increased (Figures 1D and 1E). Results indicated that the asthma model was established successfully.
Figure 1 HDM-induced asthmatic mice exhibited an impaired airway epithelium, aggravated inflammation, and allergic airway hyperresponsiveness. (A) HDM-induced mouse model of asthma. (B) Representative hematoxylin and eosin (H&E) staining of lung sections of control and asthmatic mice (100X magnification), scale bar ˂ 100 μm. (C) Penh values of inhaled methacholine. (D) The number of total cells and the percentage of eosinophils in 200 cells/slide in BALF. (E) Serum total IgE levels. *P ˂ 0.05; **P ˂ 0.01.
Effect of Vitamin A on airway inflammation, epithelial integrity, and mucus secretion in asthmatic mice
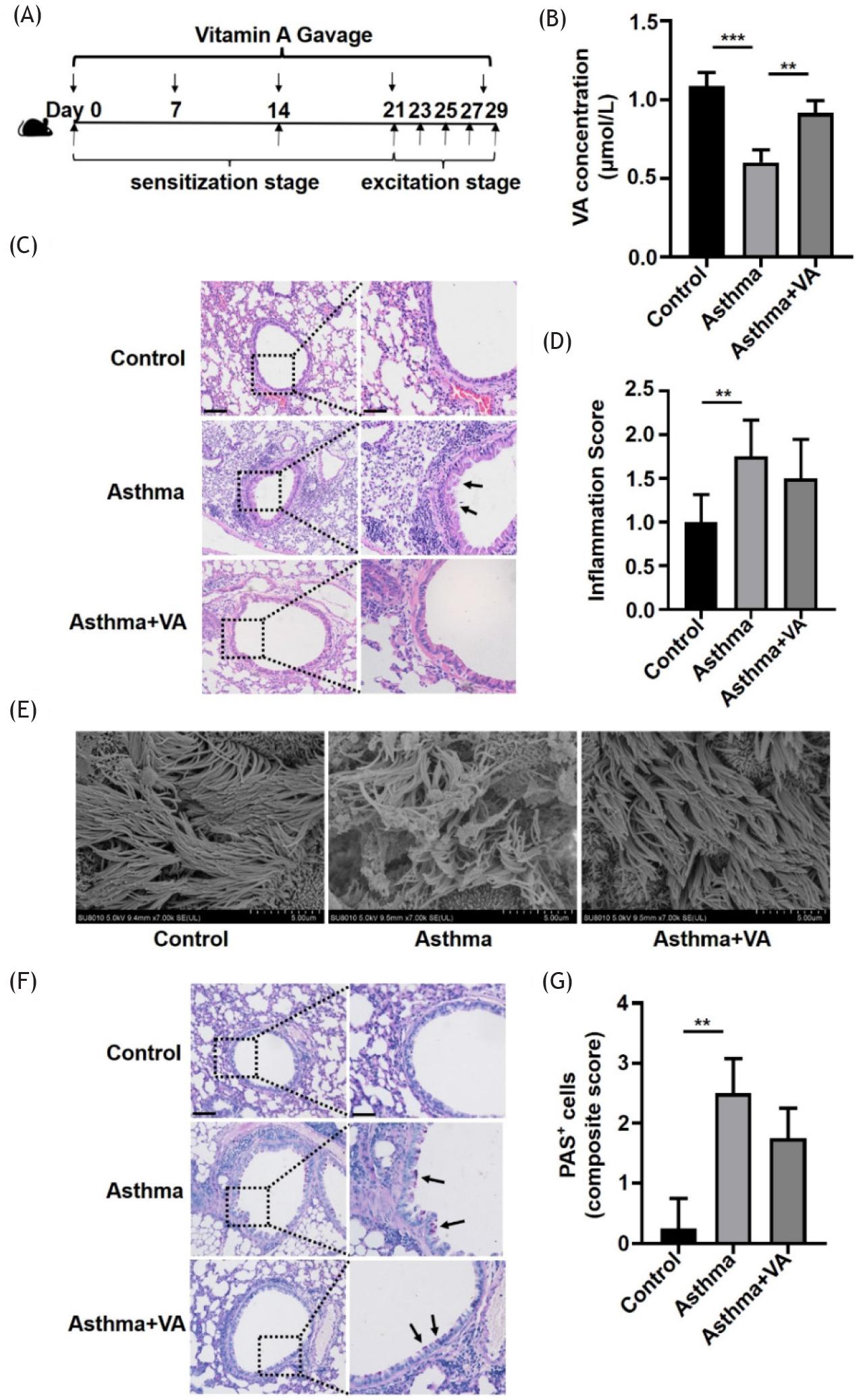
The flow chart shows the VA intervention in asthmatic mice (Figure 2A). The concentration of VA in serum significantly decreased in the asthma group and increased in the VA-treated asthma group, as shown by HPLC (Figure 2B). After VA treatment, H&E staining showed that the airway epithelium was more intact than in the untreated asthmatic mice. In the asthma group, the perivascular and peribronchiolar inflammation was obvious, and VA tended to reduce inflammation, but it was not statistically significant (Figures 2C and 2D). Using SEM, we observed that the cilia of trachea epithelium were orderly arranged with regular morphology and consistent direction in control mice. While the cilia of asthmatic mice were disordered and sparse, some of them had mucus adhesion. After VA treatment, the morphology of cilia recovered obviously (Figure 2E). We detected mucus secretion by glycogen staining and showed that secretion was significantly higher in the asthma group, but there was no significant difference between the VA-treated asthma group and the asthma group (Figures 2F and 2G). This suggested that VA repaired cilia to maintain the integrity of the airway epithelium in asthmatic mice.
Figure 2 VA influenced airway inflammation, epithelial integrity, and mucus secretion in asthmatic mice. (A) VA-treated mouse model of asthma. (B) Serum VA concentrations in mice from the control, asthma, and VA-treated asthma groups. (C) Representative hematoxylin and eosin (H&E) staining of lung sections of mice from the control, asthma, and VA-treated asthma groups (100X and 200X magnifications). Left panel scale bar ˂ 100 μm; right panel scale bar = 200 μm. (D) Inflammation score as shown in (C). (E) SEM images of trachea epithelium of mice from the control, asthma, and VA-treated asthma groups. (F) Glycogen staining of lung sections of mice from control, asthma, and VA-treated asthma groups (100X and 200X magnifications). Left panel scale bar = 100 μm; right panel scale bar = 200 μm. (G) Quantification of mucus secretion as shown in (F). *P ˂ 0.05, **P ˂ 0.01, and ***P ˂ 0.001.
Vitamin A regulated the expression of characteristic makers in ciliated and secretory cells in asthmatic mice
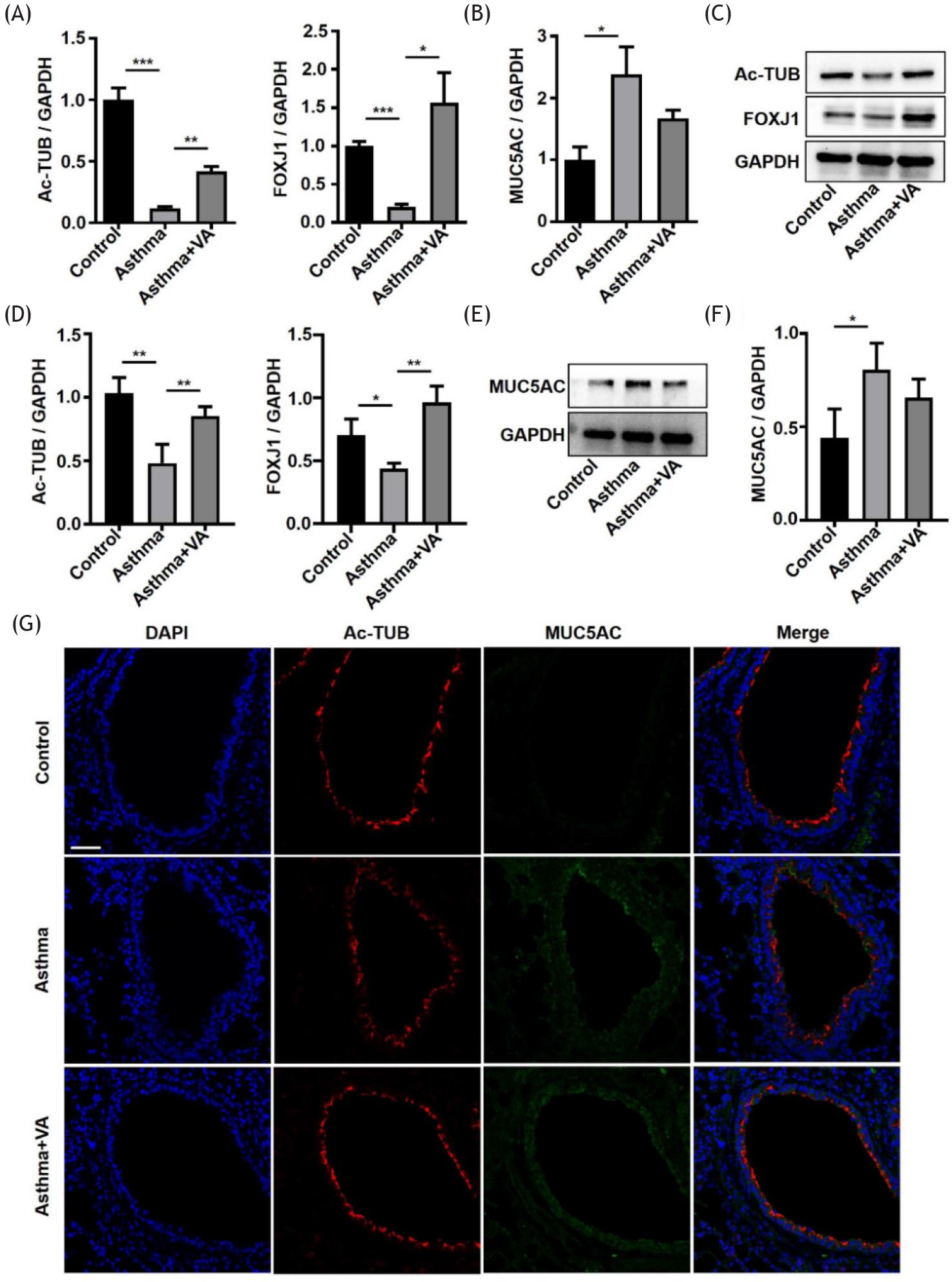
We used qRT-PCR to detect the mRNA expression of characteristic markers in ciliated cells (Ac-TUB and FOXJ1) and secretory cells (MUC5AC) in the lung tissues of mice.19–21 The results showed that the mRNA expression of Ac-TUB and FOXJ1 in the asthma group was significantly lower than in the control group, while the expression of MUC5AC in the asthma group was higher than in the control group. The mRNA expression of Ac-TUB and FOXJ1 in the VA-treated asthma group was significantly higher than in the asthma group, while MUC5AC was not significantly different in the two groups (Figures 3A and 3B). We also verified the protein expression of Ac-TUB, FOXJ1, and MUC5AC by WB. Compared with the control group, Ac-TUB and FOXJ1 decreased and MUC5AC increased in the asthma group. After VA treatment, Ac-TUB and FOXJ1 protein increased significantly (Figures 3C, 3D, 3E, and 3F). We also confirmed this by immunofluorescence and observed that Ac-TUB was disorganized in the airway epithelium of asthmatic mice. After VA intervention, the alignment of Ac-TUB was more orderly, and the integrity of the airway epithelium was restored (Figure 3G). The results showed that asthma caused decreased expression of Ac-TUB and FOXJ1 and increased expression of MUC5AC. VA mostly reversed the effect of asthma on Ac-TUB and FOXJ1. This is of great significance to the repair of airway epithelial integrity.
Figure 3 VA regulated the expression of Ac-TUB, FOXJ1, and MUC5AC in asthmatic mice. (A) Ac-TUB and FOXJ1 mRNA expression levels in lung tissues. (B) MUC5AC mRNA expression level in lung tissues. (C) Representative WB of Ac-TUB and FOXJ1 in lung tissues. (D) Quantification of WB data from (C). (E) Representative WB of MUC5AC in lung tissues. (F) Quantification of WB data from (E). (G) Immunofluorescent staining for Ac-TUB and MUC5AC in lung sections (400X magnification), scale bar = 50 μm. *P ˂ 0.05, **P ˂ 0.01, and ***P ˂ 0.001.
ATRA affected the expression of Ac-TUB, FOXJ1, and MUC5AC in 16HBE cells
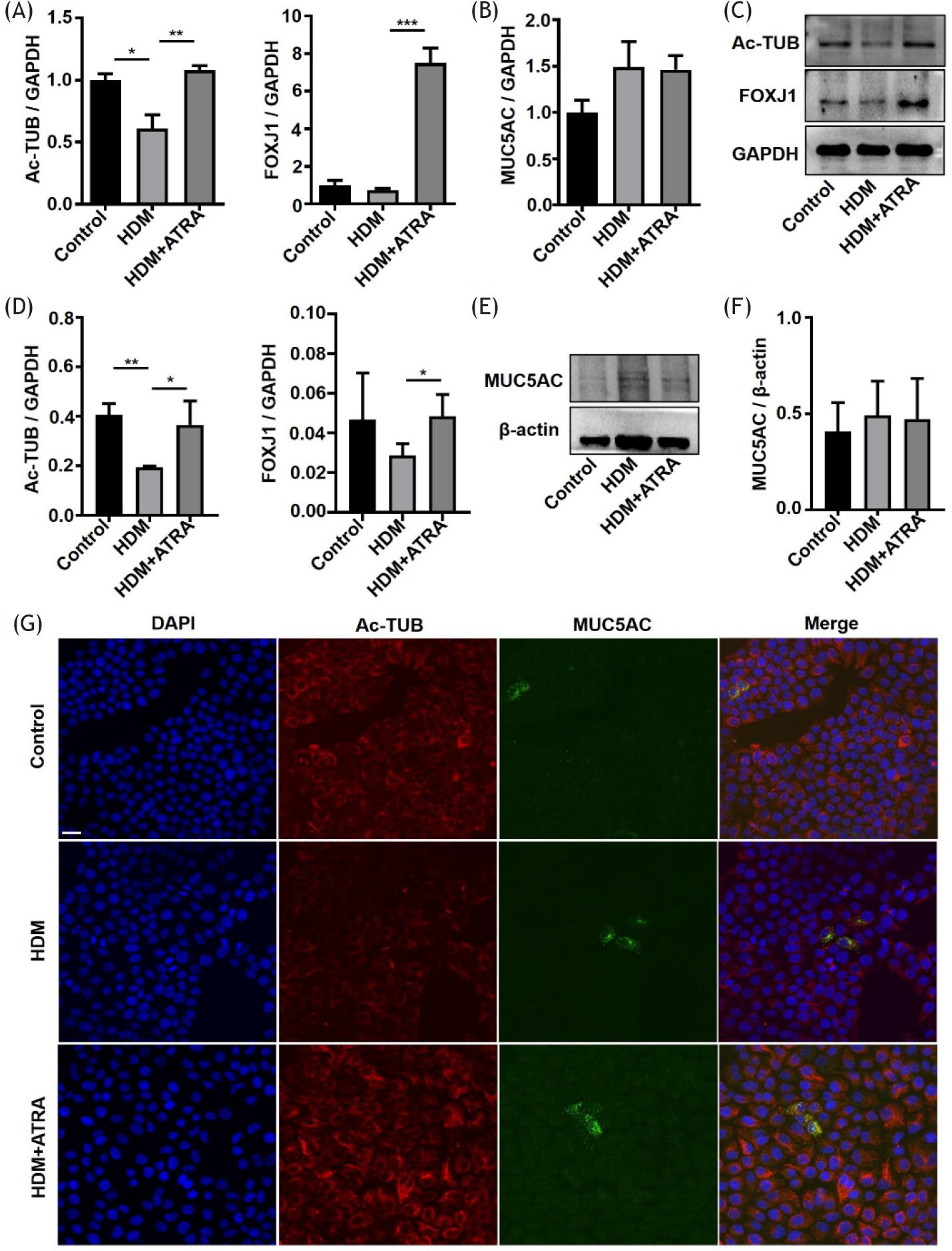
We characterized 16HBE cells by investigating the expression of Ac-TUB, FOXJ1, and MUC5AC by qRT-PCR, WB, and immunofluorescence, respectively. We found that the mRNA expression of Ac-TUB in the HDM group was lower than in the control group, and FOXJ1 showed no statistical difference. The expression of Ac-TUB and FOXJ1 in the HDM + ATRA group was significantly higher than in the HDM group. The mRNA expression of MUC5AC in the three groups was not significantly different in 16HBE cells (Figures 4A and 4B). This suggested that HDM reduced the mRNA expression of Ac-TUB, and ATRA increased the expression of Ac-TUB and FOXJ1. The WB showed that HDM mainly decreased the protein expression of Ac-TUB, and ATRA significantly increased the expression of Ac-TUB and FOXJ1 (Figures 4C and 4D). There was also no significant difference in the protein expression of MUC5AC (Figures 4E and 4F). We observed the positive visual expression of Ac-TUB and MUC5AC by immunofluorescence staining. Ac-TUB protein was also reduced in 16HBE cells treated with HDM compared to the control group; after ATRA intervention, Ac-TUB was increased. Expression of the MUC5AC protein was not significantly changed (Figure 4G). In vitro results also suggested that ATRA mainly affected the expression of characteristic markers in ciliated cells.
Figure 4 ATRA regulated the expression of Ac-TUB, FOXJ1, and MUC5AC in 16HBE cells. (A) Ac-TUB and FOXJ1 mRNA expression levels in 16HBE cells. (B) MUC5AC mRNA expression level in 16HBE cells. (C) Representative WB of Ac-TUB and FOXJ1 in 16HBE cells. (D) Quantification of WB data from (C). (E) Representative WB of MUC5AC in 16HBE cells. (F) Quantification of WB data from (E). (G) Immunofluorescent staining for Ac-TUB and MUC5AC in 16HBE cells (400X magnification), scale bar = 20 μm. *P ˂ 0.05, **P ˂ 0.01, and ***P ˂ 0.001.
Discussion
Asthma is a global public health issue that occurs in all age groups and requires an urgent solution. Asthma endangers the health of adults and children, and imposes a large burden in the form of morbidity, mortality, and economic costs.22,23 Clinically, symptoms of asthma remain uncontrolled in many patients following routine treatment, greatly affecting the quality of life (QoL). Research has revealed that patients exhibiting better symptom control have higher QoL scores; however, in asthma patients, QoL decreased consistently from onset.24
The airway epithelium is a defensive barrier against physical, chemical, and biological stressors. It is the first barrier of the bronchopulmonary tissue to resist the external environment and is essential to maintaining homeostasis. The airway epithelium is mainly involved in host defense, the inflammatory response, immune regulation, and airway remodeling.25,26 Tight junctions between cells maintain the integrity of the airway epithelial structure.27 The airway epithelium expresses and secretes anti-injury factors such as epidermal growth factor receptor, vasoactive intestinal peptide, and lipoxins to reduce the inflammatory response and promote epithelial repair.28–30 Pattern recognition receptors are also involved in immune regulation.31 Airway epithelial cells are mainly composed of ciliated cells, and cilia constitute the most important part of the mucociliary clearance system, as they have a key role in the function of the airway epithelium. Allergens, viruses, and inflammatory response are external factors that impair the airway epithelial barrier. If the structure and function are destroyed, abnormal cilia will induce mucus clearance disorder and a series of damaging effects. Mucus movement and clearance rates are decreased significantly, resulting in mucus accumulation. Local infection of the airway is then aggravated, causing disturbance of the mucociliary clearance system, further aggravating airway stenosis and causing varying degrees of ventilation disorders. Therefore, the airway epithelium is a specialized barrier performing multiple functions, and any modification to this barrier or its components can lead to lung diseases. Persistent chronic airway inflammation leads to continuous injury of the airway epithelium in asthma patients. If the normal repair process is destroyed, disorder may lead to mucociliary clearance dysfunction, ciliated cell damage and exfoliation, goblet cell metaplasia, high mucus secretion, and resulting airway remodeling. In a study of children with asthma, endobronchial biopsies showed that the ciliated cells exhibited a loss of cilia in some cases.32 Our research also found that the cilia of tracheal epithelium in asthmatic mice were destroyed and appeared disorder and sparseness, with mucous attachment. Ac-TUB and FOXJ1 were decreased, while MUC5AC was significantly increased. These results are consistent with the pathophysiological characteristics of asthma. Ciliated cells maintain barrier integrity, and structure destruction inevitably leads to a loss of function. Promoting airway epithelium repair has become a research target in the prevention and treatment of asthma.
VA is necessary for the maintenance of normal metabolism and function of the epithelium. Its metabolites are involved in cell proliferation and differentiation, immune regulation, vision, bone metabolism, reproduction, and other physiological functions; deficiency leads to abnormal physiological functions and pathological changes.33 VA could also promote wound healing, and a study reported that topical VA promoted mucociliary reepithelization during sinonasal wound healing after endoscopic sinus surgery.34 Experiments in mouse models and cells in vitro have also confirmed that VA and its active derivatives promoted wound healing of corneal epithelial cells.35 VA deficiency is more common during infection, and supplementation reduces severe morbidity and mortality from infectious diseases.36 Our study explored the effect of VA on the airway epithelium in asthmatic mice. The results showed that VA repaired cilia, increased expression of characteristic markers Ac-TUB and FOXJ1 in ciliated cells, and maintained airway epithelial integrity. Therefore, the repair effect of VA in the airway epithelium of asthmatic mice was mainly through regulating ciliated cells.
In summary, asthma is a respiratory disease that is caused by inflammation, mucus overproduction, and airway remodeling, leading to bronchial hyperreactivity and obstruction. Currently, the use of corticosteroids cannot completely control the recurrence and aggravation of asthma in some patients, illustrating the need to identify other exploratory treatments. The maintenance of airway epithelial integrity and restoration of function is particularly important. The fate of the airway epithelium determines the severity and prognosis of asthma. Therefore, we investigated the role of VA in the airway epithelium of asthmatic mice and clarified that VA promoted airway barrier repair by acting on ciliated cells to maintain the integrity of the epithelium. These results provide a potential new treatment strategy for asthma.
Conclusion
Damage to the airway epithelium is an important factor that leads to recurrent attacks and aggravation of asthma. Our group previously confirmed that glucocorticoid treatment partially inhibited the repair of airway epithelium after injury. Thus far, there is no pharmacological intervention to repair the airway epithelium in the treatment of asthma. In this study, we confirmed that the role of VA in reversing airway epithelial injury caused by asthma. To the best of our knowledge, this is the first study to demonstrate that VA regulates ciliated cells to repair the damaged airway epithelium and maintain its integrity. Additionally, this study provides a potential new treatment strategy for treating patients with asthma and lays a foundation for further studies on the mechanism.
Acknowledgements
This work was supported by the National Natural Science Foundation of China (No. 81900023).
REFERENCES
1. Serebrisky D, Wiznia A. Pediatric asthma: A global epidemic. Ann Glob Health. 2019;85(1):6. 10.5334/aogh.2416
2. Liu J, Zhang M, Niu C, Luo Z, Dai J, Wang L, et al. Dexamethasone inhibits repair of human airway epithelial cells mediated by glucocorticoid-induced leucine zipper (GILZ). PLoS One. 2013;8(4):e60705. 10.1371/journal.pone.0060705
3. Hupin C, Gohy S, Bouzin C, Lecocq M, Polette M, Pilette C. Features of mesenchymal transition in the airway epithelium from chronic rhinosinusitis. Allergy. 2014;69(11):1540–9. 10.1111/all.12503
4. Whitsett JA. Airway Epithelial differentiation and mucociliary clearance. Ann Am Thorac Soc. 2018;15(Suppl 3):S143–8. 10.1513/AnnalsATS.201802-128AW
5. Gon Y, Hashimoto S. Role of airway epithelial barrier dysfunction in pathogenesis of asthma. Allergol Int. 2018;67(1):12–7. 10.1016/j.alit.2017.08.011
6. Hellings PW, Steelant B. Epithelial barriers in allergy and asthma. J Allergy Clin Immunol. 2020;145(6):1499–509. 10.1016/j.jaci.2020.04.010
7. Xiao C, Puddicombe SM, Field S, Haywood J, Broughton-Head V, Puxeddu I, et al. Defective epithelial barrier function in asthma. J Allergy Clin Immunol. 2011;128(3):549–56.e1–12. 10.1016/j.jaci.2011.05.038
8. Laulajainen-Hongisto A, Toppila-Salmi SK, Luukkainen A, Kern R. Airway epithelial dynamics in allergy and related chronic inflammatory airway diseases. Front Cell Dev Biol. 2020;8:204. 10.3389/fcell.2020.00204
9. Samitas K, Carter A, Kariyawasam HH, Xanthou G. Upper and lower airway remodelling mechanisms in asthma, allergic rhinitis and chronic rhinosinusitis: The one airway concept revisited. Allergy. 2018;73(5):993–1002. 10.1111/all.13373
10. Zhang ML, Tao Y, Zhou WQ, Ma PC, Cao YP, He CD, et al. All-trans retinoic acid induces cell-cycle arrest in human cutaneous squamous carcinoma cells by inhibiting the mitogen-activated protein kinase-activated protein 1 pathway. Clin Exp Dermatol. 2014;39(3):354–60. 10.1111/ced.12227
11. Samarasinghe AE, Penkert RR, Hurwitz JL, Sealy RE, LeMessurier KS, Hammond C, et al. Questioning cause and effect: Children with severe asthma exhibit high levels of inflammatory biomarkers including beta-hexosaminidase, but low levels of vitamin A and immunoglobulins. Biomedicines. 2020;8(10):393. 10.3390/biomedicines8100393
12. Bai YJ, Dai RJ. Serum levels of vitamin A and 25-hydroxyvitamin D3 (25OHD3) as reflectors of pulmonary function and quality of life (QOL) in children with stable asthma: A case-control study. Medicine (Baltimore). 2018;97(7):e9830. 10.1097/MD.0000000000009830
13. Talaei M, Hughes DA, Mahmoud O, Emmett PM, Granell R, Guerra S, Shaheen SO. Dietary intake of vitamin A, lung function and incident asthma in childhood. Eur Respir J. 2021;58(4):2004407. 10.1183/13993003.04407-2020
14. Wang Y, Liao K, Liu B, Niu C, Zou W, Yang L, et al. GITRL on dendritic cells aggravates house dust mite-induced airway inflammation and airway hyperresponsiveness by modulating CD4(+) T cell differentiation. Respir Res. 2021;22(1):46. 10.1186/s12931-020-01583-x
15. Niu C, Liu N, Liu J, Zhang M, Ying L, Wang L, et al. Vitamin A maintains the airway epithelium in a murine model of asthma by suppressing glucocorticoid-induced leucine zipper. Clin Exp Allergy. 2016;46(6):848–60. 10.1111/cea.12646
16. Triantaphyllopoulos K, Hussain F, Pinart M, Zhang M, Li F, Adcock I, et al. A model of chronic inflammation and pulmonary emphysema after multiple ozone exposures in mice. Am J Physiol Lung Cell Mol Physiol. 2011;300(5):L691–700. 10.1152/ajplung.00252.2010
17. Tan M, Yang T, Liu H, Xiao L, Li C, Zhu J, et al. Maternal vitamin A deficiency impairs cholinergic and nitrergic neurons, leading to gastrointestinal dysfunction in rat offspring via RARβ. Life Sci. 2021;264:118688. 10.1016/j.lfs.2020.118688
18. Chen S, Yun F, Yao Y, Cao M, Zhang Y, Wang J, et al. USP38 critically promotes asthmatic pathogenesis by stabilizing JunB protein. J Exp Med. 2018;215(11):2850–67. 10.1084/jem.20172026
19. Giuranno L, Roig EM, Wansleeben C, van den Berg A, Groot AJ, et al. NOTCH inhibition promotes bronchial stem cell renewal and epithelial barrier integrity after irradiation. Stem Cells Transl Med. 2020;9(7):799–812. 10.1002/sctm.19-0278
20. Griggs TF, Bochkov YA, Basnet S, Pasic TR, Brockman-Schneider RA, Palmenberg AC, et al. Rhinovirus C targets ciliated airway epithelial cells. Respir Res. 2017;18(1):84. 10.1186/s12931-017-0567-0
21. You Y, Huang T, Richer EJ, Schmidt JH, Zabner J, Borok Z, et al. Role of f-box factor foxj1 in differentiation of ciliated airway epithelial cells. Am J Physiol Lung Cell Mol Physiol. 2004;286(4):L650–7. 10.1152/ajplung.00170.2003
22. Kauppi P, Linna M, Jantunen J, Martikainen JE, Haahtela T, Pelkonen A, et al. Chronic comorbidities contribute to the burden and costs of persistent asthma. Mediators Inflamm. 2015;2015:819194. 10.1155/2015/819194
23. Wang L, Zhong Y, Wheeler L. Direct and indirect costs of asthma in school-age children. Prev Chronic Dis. 2005;2(1):A11.
24. Uchmanowicz B, Panaszek B, Uchmanowicz I, Rosinczuk J. Clinical factors affecting quality of life of patients with asthma. Patient Prefer Adherence. 2016;10:579–89. 10.2147/PPA.S103043
25. Proud D, Leigh R. Epithelial cells and airway diseases. Immunol Rev. 2011;242(1):186–204. 10.1111/j.1600-065X.2011.01033.x
26. Tam A, Wadsworth S, Dorscheid D, Man SF, Sin DD. The airway epithelium: More than just a structural barrier. Ther Adv Respir Dis 2011;5(4):255–73. 10.1177/1753465810396539
27. Frey A, Lunding LP, Ehlers JC, Weckmann M, Zissler UM, Wegmann M. More than just a barrier: The immune functions of the airway epithelium in asthma pathogenesis. Front Immunol. 2020;11:761. 10.3389/fimmu.2020.00761
28. Davies DE. The role of the epithelium in airway remodeling in asthma. Proc Am Thorac Soc. 2009;6(8):678–82. 10.1513/pats.200907-067DP
29. Szema AM, Hamidi SA, Koller A, Martin DW. Vasoactive intestinal peptide knockout (VIP KO) mouse model of sulfite-sensitive asthma: Up-regulation of novel lung carbonyl reductase. BMC Immunol. 2011;12:66. 10.1186/1471-2172-12-66
30. Grumbach Y, Quynh NV, Chiron R, Urbach V. LXA4 stimulates ZO-1 expression and transepithelial electrical resistance in human airway epithelial (16HBE14o-) cells. Am J Physiol Lung Cell Mol Physiol. 2009;296(1):L101–8. 10.1152/ajplung.00018.2008
31. Lambrecht BN, Hammad H. The airway epithelium in asthma. Nat Med. 2012;18(5):684–92. 10.1038/nm.2737
32. Cokuğraş H, Akçakaya N, Seçkin I, Camcioğlu Y, Sarimurat N, Aksoy F. Ultrastructural examination of bronchial biopsy specimens from children with moderate asthma. Thorax. 2001;56(1):25–9. 10.1136/thorax.56.1.25
33. Bar-El Dadon S, Reifen R. Vitamin A and the epigenome. Crit Rev Food Sci Nutr. 2017;57(11):2404–11. 10.1080/10408398.2015.1060940
34. Fang KM, Wang CT, Chen YW, Huang TW. Reduction of adhesions and antrostomy stenosis with topical vitamin A after endoscopic sinus surgery. Am J Rhinol Allergy. 2015;29(6):430–4. 10.2500/ajra.2015.29.4235
35. Comptour A, Rouzaire M, Belville C, Bonnin N, Daniel E, Chiambaretta F, et al. Lysyl oxidase-like 4 involvement in retinoic acid epithelial wound healing. Sci Rep. 2016;6:32688. 10.1038/srep32688
36. Zinder R, Cooley R, Vlad LG, Molnar JA. Vitamin A and wound healing. Nutr Clin Pract. 2019;34(6):839–49. 10.1002/ncp.10420