
Download
REVIEW ARTICLE
MicroRNA-155 is a main part of proinflammatory puzzle during severe coronavirus disease 2019 (COVID-19)
Asieh Asadpour-Behzadia, Ashraf Kariminika*, Babak Kheirkhahb
aDepartment of Microbiology, Kerman Branch, Islamic Azad University, Kerman, Iran
bDepartment of Veterinary Medicine, Baft Branch, Islamic Azad University, Baft, Iran
Abstract
Genetic and epigenetic parameters play critical roles in determining the outcomes of the severe acute respiratory syndrome coronavirus type 19 (SARS-CoV-2) infection. MicroRNAs (miRNAs) are an important part of the epigenetic factors that regulate several functions of the immune cells and also viruses. Accordingly, the molecules can regulate expression of the immune cell proteins and virus in the host cells. Among the miRNAs, miRNA-155 (miR-155) is well-studied in patients suffering from severe coronavirus disease 2019 (COVID-19). It has been reported that the SARS-CoV-2 infected patients may be directed to induce a cytokine storm or severe proinflammatory responses. This review article discusses the pathological roles of miR-155 during COVID-19 infection.
Key words: SARS-CoV-2, Coronavirus Disease 2019, MiR-155, Inflammation
*Corresponding author: Ashraf Kariminik, Department of Microbiology, Kerman Branch, Islamic Azad University, Kerman, Iran. Email address: [email protected]
Received 12 June 2022; Accepted 28 August 2022; Available online 1 March 2023
Copyright: Asadpour-Behzadi A, et al.
This open access article is licensed under Creative Commons Attribution 4.0 International (CC BY 4.0). http://creativecommons.org/licenses/by/4.0/
Introduction
Coronavirus disease 2019 (COVID-19) is the name of a disorder, which is induced by SARS-CoV2 virus.1 Immune responses against COVID-19 either control the virus replication or induce severe inflammation and cytokine storm in the infected patients.2 Accordingly, immune response severities against the virus are considered as the main factor to determine COVID-19 outcome.2 Immune cell functions are multifactorial and are dependent on the host genetic and epigenetic conditions and also environmental factors.3 Therefore, the immune cell regulators that influence immune cell functions may alter specific immune responses to COVID-19, and can be considered as the risk factors for modulation of the cytokine storm. Additionally, lung and heart failure are the most lethal complications of COVID-19, which occur after acute forms of the disorder.4 The complications can be induced by SARS-CoV2 virus and also by the severe immune responses to the virus.4 Thus, the genetic and epigenetic parameters that alter immune responses to the SARS-CoV2 may participate in the tissue-related failures during COVID-19.
MicroRNAs (miRNAs) are important epigenetic factors that affect post transcription of several proteins.5 Accordingly, the molecules participate in several cell functions, including activation, proliferation, differentiation, apoptosis, and growth.6,7 Thus, it has been hypothesized that malregulation of miRNAs is a critical factor in the pathogenesis of several immune system-related disorders.8–11 Therefore, it is plausible to hypothesize that miRNAs may play crucial roles in the viral infection pathogenesis. MiRNA-155 (miR-155) is a candidate for regulation of immune responses against viral infections and their related complications.12–14 Due to the various immune responses against SARS-CoV2 in patients it has been hypothesized that the expression and functions of miR-155 may be altered during COVID-19. Interestingly, it has been documented that miR-155 is expressed differently in the hospitalized SARS-CoV2-infected patients.15,16 So, it appears that miR-155 may participate in several aspects of immune responses against SARS-CoV2, including adaptive and innate immunity. Therefore, this article discusses the recent information regarding the plausible roles played by miR-155 in the pathogenesis of COVID-19.
MicroRNA-155
MiRNAs are the noncoding small (21–25 nucleotides) RNAs with various biological functions. The molecules are created from a pri-miRNA, and some miRNAs may be clustered on a pri-miRNA.17 Pri-miRNA maturation by Drosha, Dicer, and RNA-induced silencing complex (RISC) lead to the pre-miRNAs production, miRNA duplex, and mature forms that are the secondary, tertiary, and last versions of the miRNAs, respectively.18 The mature miRNA plus RISC complex have the biological functions to regulate translation from several genes.19
MIRHG155 is the responsible gene for coding miR-155, which is located in the human chromosome 21 (Hsa21) and has three exons.20,21 MiR-155 expression is significantly different in the human cell types and can be regulated by several intracellular signaling pathways. Interestingly, there are binding sites for nuclear factor-kappa B (NF-κB), mothers against decapentaplegic homolog 4 (SMAD4), interferon- sensitive response element (ISRE), interferon regulatory factors (IRF), hypoxia-inducible factor-1 alpha, and activator protein 1 (AP-1) at upstream region of B-cell integration cluster (BIC) gene, the location for miR-155, which confirmed the roles played by the factors for expression of miR-155.22 As mentioned previously, miRNAs need to undergo sequential processing to be pri-miRNA, pre-miRNAs, and mature miRNAs. Pri-miR-155 is made from 65 nucleotides (nts)–long stem-loop precursor, and pre-miR-155 from ~22-nts duplexes.23 Mature miR-155 can be produced in two forms, including miR-155-5p, the most abundant and functionally dominant form, and miR-155-3p.23,24 In this article, all the miR-155 are referred to miR-155-5p, and it is declared in the case of miR-155-3p in the reports.
Micro-RNA-155 and COVID-19
Previous investigations demonstrated that miR-155 is a key factor that participates in the induction of inflammation in several human disorders.25–27 Additionally, its critical roles during viral infection–related inflammation have also been documented by several studies, which confirmed that miR-155 can direct immune responses to acute inflammation, and in some cases the molecule modulates immune responses.28–30 Therefore, the molecule may play different roles during COVID-19. Most studies have demonstrated that miR-155 is upregulated in the SARS-CoV-2-infected patients who suffer from excess inflammation. For example, a study by Garg et al. revealed that the patients that suffered from severe COVID-19 had higher serum levels of miR-155 when compared to healthy controls and patients with influenza-associated acute respiratory distress syndrome (Influenza-ARDS).31 Thus, it appears that SARS-CoV-2 significantly induced inflammation independent of miR-155. A study on the Iranian population revealed that the patients with acute phase of COVID-19 had higher levels of miR-155 when compared with a healthy control group.32 Therefore, it seems that the ethnicity had no effects on the expression of miR-155 in the SARS-CoV-2-infected patients. Additionally, it has been hypothesized that miR-155 may participate in the deterioration of the disease complications. A study on the Spanish population approved the hypothesis and revealed that serum levels of miR-155 were increased in COVID-19 patients, and the elevation had a positive correlation with increase of the disease grade.33 The study confirmed that expression of the anti-inflammatory molecules, including peroxisome proliferator-activated receptors (PPARS), cytokine signaling 1 (SOCS1) suppressor, and CCAAT enhancer-binding protein alpha (CEBPA), which are targeted by miR-155, were decreased in COVID-19 patients.33 Therefore, it seems that miR-155 via downregulation of anti-inflammatory molecules induces inflammation during COVID-19. Some investigations revealed that miR-155 level is a main factor for the upregulation of inflammatory molecules, in parallel with decrease of anti-inflammatory molecules. For instance, Ahmed et al. by using the protein–protein interaction (PPI) network technique analyzed 138 differentially expressed genes (DEGs) between COVID-19 patients and healthy controls.34 The analysis revealed 11 key DEGs, including receptor-interacting serine or threonine protein kinase 1 (RIPK1), SMAD4, glycogen synthase kinase-3 beta (GSK3B), ataxia-telangiectasia mutated (ATM), Sirtuin 1 (SIRT1), protein kinase CAMP-activated catalytic subunit beta (PRKACB), mediator complex subunit 17 (MED17), chaperonin containing TCP1 subunit 2 (CCT2), baculoviral IAP repeat containing 3 (BIRC3), erythroblast transformation-specific (ETS) proto-oncogene 1 (ETS1), and thioredoxin (TXN).34 Interestingly, the investigators showed that miR-155 can be considered as the key post-transcriptional regulator of DEGs.34 Therefore, it appears that miR-155 targets several molecules and regulates a molecular network to induce inflammation in the SARS-CoV-2-infected patients. Accordingly, miR-155 can participate in the induction of the target tissues in the SARS-CoV-2-infected patients. Interestingly, a case report study confirmed the hypothesis and reported that expression of miR-155 and its proinflammatory target, IL-6, is associated with vascular inflammation in the COVID-19 patients.35 Molinero et al. confirmed the proinflammatory roles played by miR-155 during pathological conditions in lung tissues.36 Based on the roles played by miR-155 in the induction of inflammation in COVID-19, it has been hypothesized that miR-155 expression can be associated with the complications of COVID-19. Haroun et al. proved the hypothesis and reported that the expression level of miR-155 had significant correlation with clinicopathological characteristics of COVID-19 patients, such as chest CT findings, CRP, ferritin, mortality, D-dimer, WBC count, and percentage of lymphocytes and neutrophils.16 The study also showed that miR-155 plasma levels significantly increased in COVID-19 patients, during severe symptoms, and in deceased patients when compared to healthy controls, moderate symptoms and surviving COVID-19 patients, respectively.16 Donyavi et al. demonstrated that serum levels of miR-155 not only increased in the patients suffering from acute phases of COVID-19 but also remained high in the post-acute phases of the disease.37 Thus, miR-155 may participate in some post-COVID-19 complications, which need further investigations.
Due to the results, it seems that miR-155 is an important factor to determine the outcome of COVID-19 via increased inflammation in the patients. Thus, it may be hypothesized that targeting miR-155 may be considered as a critical mechanism to modulate excess inflammation in the hospitalized patients. In addition to targeting miR-155 directly, it may be proposed to regulate the mechanisms that lead to upregulation of miR-155. It has been reported that long noncoding RNAs are the main factors for upregulation of miR-155 in the SARS-CoV-2-infected patients.38 Several inflammatory stimuli, such as pathogen-associated molecular patterns (PAMPs), proinflammatory cytokines, and damage- associated molecular patterns (DAMPs) can also increase the expression of miR155 and then, miR-155 could stimulate the inflammatory response in a positive feedback manner.39,40 Additionally, it has been demonstrated that miR-155 can target SARS-CoV-2 genome,39 hence, increased copy number of the virus can induce expression of miR-155 directly. However, the expression of miR-155 is multifactorial and needs to be explored by further investigations.
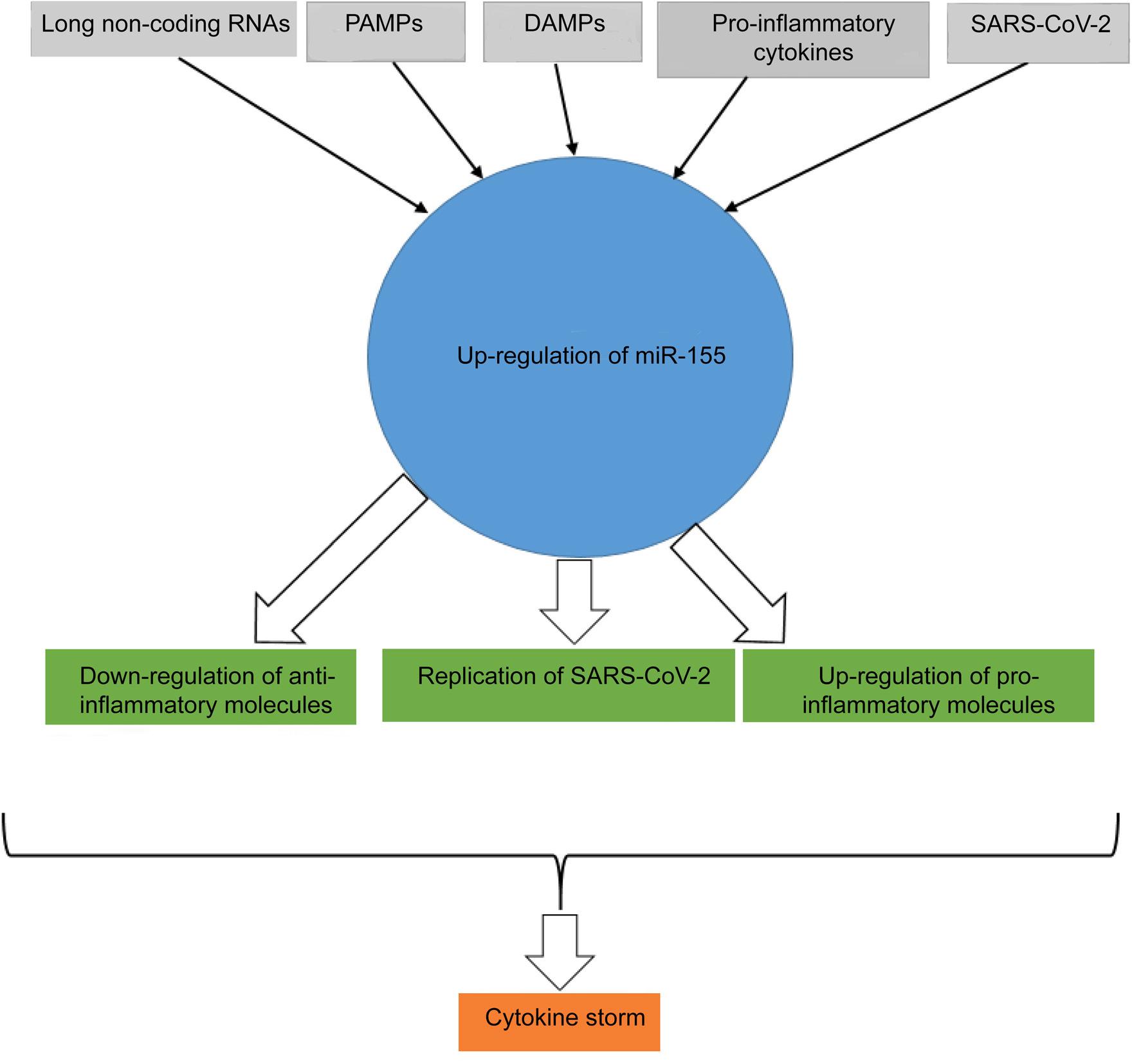
It is worthy to note that the roles played by miR-155 in the pathogenesis of COVID-19 may be related to several interfered factors, such as type of vaccination. It has been reported that miR-155 expression may be decreased in COVID-19 in some situations. For example, Li et al. revealed that miR-155 is upregulated in the COVID-19 patients when compared to healthy controls, however, it was downregulated in the patients with severe or critical (SC) when compared to mild or moderate (MM) symptoms.41 A study by Kassif-Lerner et al. demonstrated that the COVID-19 patients with mild and severe symptoms had 2.5 and 5-fold less circulating miR-155, respectively, than healthy people.15 This is a unique report regarding downregulation of miR-155 in the COVID-19 patients.The study was performed on a low sample size (37 patients and 15 healthy controls), hence, it seems that their result cannot be considered as significant data. Moreover, based on several investigations with higher sample sizes that proved upregulation of miR-155 in COVID-19 patients, it appears that upregulation of miR-155 and its related inflammation is reliable. Additionally, Oh et al. suggested that miR-155 may be considered as an important factor for induction of myocarditis during vaccination against SARS-CoV-2.42 Figure 1 illustrates the factors involved in upregulation of miR-155 and the roles played by the molecule on the increased inflammation during COVID-19 disease.
Figure 1 The known inducers of miR-155 in COVID-19 and its complications. Long noncoding RNAs, PAMPs, DAMPs, proinflammatory cytokines, and SARS-CoV-2 genome are the main inducers of miR-155. Upregulation of miR-155 is associated with down- and upregulation of anti- and proinflammatory molecules, respectively, and proliferation of SARS-CoV-2. The phenomenon leads to COVID-19 compilations, such as cytokine storm.
Future Perspective
Based on the fact that the report is unique and the researchers from other countries with different vaccination had no reports, it seems that the type of vaccination may alter the expression and function of miR-155.
Conclusion
Authors propose to explore the effects of miR-155 silencers to overcome the inflammation and increase survival in clinical trial studies.
Acknowledgments
This project was funded by the Islamic Azad University of Kerman, Iran.
Conflict of Interest
The authors declare no potential conflicts of interest with respect to research, authorship, and/or publication of this article.
REFERENCES
1. Yüce M, Filiztekin E, Özkaya KG. COVID-19 diagnosis—A review of current methods. Biosens Bioelectron. 2021;172:112752. 10.1016/j.bios.2020.112752
2. Hu B, Huang S, Yin L. The cytokine storm and COVID-19. J Med Virol. 2021;93(1):250–6. 10.1002/jmv.26232
3. Abbas AK, Lichtman AH, Pillai S. Cellular and molecular immunology. Elsevier Health Sciences: Philadelphia, PA; 2012.pp.52.
4. Aleksova A, Gagno G, Sinagra G, Beltrami AP, Janjusevic M, Ippolito G, et al. Effects of SARS-CoV-2 on cardiovascular system: The dual role of angiotensin-converting enzyme 2 (ACE2) as the virus receptor and homeostasis regulator—Review. Int J Mol Sci. 2021;22(9):4526. 10.3390/ijms22094526
5. Humphreys DT, Westman BJ, Martin DI, Preiss T. MicroRNAs control translation initiation by inhibiting eukaryotic initiation factor 4E/cap and poly(A) tail function. Proc Natl Acad Sci U S A. 2005;102(47):16961–6. 10.1073/pnas.0506482102
6. Abe M, Bonini NM. MicroRNAs and neurodegeneration: Role and impact. Trends Cell Biol. 2013;23(1):30–6. 10.1016/j.tcb.2012.08.013
7. Poy MN, Eliasson L, Krutzfeldt J, Kuwajima S, Ma X, Macdonald PE, et al. A pancreatic islet-specific microRNA regulates insulin secretion. Nature. 2004;432(7014):226-30. 10.1038/nature03076
8. Jia Y, Wei Y. Modulators of microRNA function in the immune system. Int J Mol Sci. 2020;21(7):2357. 10.3390/ijms21072357
9. Yu HR, Huang LH, Li SC. Roles of microRNA in the immature immune system of neonates. Cancer Lett. 2018;433:99–106. 10.1016/j.canlet.2018.06.014
10. Rose SA, Wroblewska A, Dhainaut M, Yoshida H, Shaffer JM, Bektesevic A, et al. A microRNA expression and regulatory element activity atlas of the mouse immune system. Nat Immunol. 2021;22(7):914–27. 10.1038/s41590-021-00944-y
11. Lu LF, Liston A. MicroRNA in the immune system, microRNA as an immune system. Immunology. 2009;127(3):291–8. 10.1111/j.1365-2567.2009.03092.x
12. Vigorito E, Kohlhaas S, Lu D, Leyland R. miR-155: An ancient regulator of the immune system. Immunol Rev. 2013;253(1):146–57. 10.1111/imr.12057
13. Testa U, Pelosi E, Castelli G, Labbaye C. miR-146 and miR-155: Two key modulators of immune response and tumor development. Noncoding RNA. 2017;3(3):22. 10.3390/ncrna3030022
14. Ibrahim SA, Afify AY, Fawzy IO, El-Ekiaby N, Abdelaziz AI. The curious case of miR-155 in SLE. Expert Rev Mol Med. 2021;23:e11. 10.1017/erm.2021.11
15. Kassif-Lerner R, Zloto K, Rubin N, Asraf K, Doolman R, Paret G, et al. miR-155: A potential biomarker for predicting mortality in COVID-19 patients. J Pers Med. 2022;12(2):324. 10.3390/jpm12020324
16. Haroun RA, Osman WH, Amin RE, Hassan AK, Abo-Shanab WS, Eessa AM. Circulating plasma miR-155 is a potential biomarker for the detection of SARS-CoV-2 infection. Pathology. 2022;54(1):104–10. 10.1016/j.pathol.2021.09.006
17. Altuvia Y, Landgraf P, Lithwick G, Elefant N, Pfeffer S, Aravin A, et al. Clustering and conservation patterns of human microRNAs. Nucleic Acids Res. 2005;33(8):2697–706. 10.1093/nar/gki567
18. Banzhaf-Strathmann J, Edbauer D. Good guy or bad guy: The opposing roles of microRNA 125b in cancer. Cell Commun Signal. 2014;12:30. 10.1186/1478-811X-12-30
19. Momeni M, Hassanshahi G, Arababadi MK, Kennedy D. Ectopic expression of micro-RNA-1, 21 and 125a in peripheral blood immune cells is associated with chronic HBV infection. Mol Biol Rep. 2014;41(7):4833–7. 10.1007/s11033-014-3355-7
20. Kim VN, Han J, Siomi MC. Biogenesis of small RNAs in animals. Nat Rev Mol Cell Biol. 2009;10(2):126–39. 10.1038/nrm2632
21. Tam W. Identification and characterization of human BIC, a gene on chromosome 21 that encodes a noncoding RNA. Gene. 2001;274(1–2):157–67. 10.1016/s0378-1119(01)00612-6
22. Bruning U, Cerone L, Neufeld Z, Fitzpatrick SF, Cheong A, Scholz CC, et al. MicroRNA-155 promotes resolution of hypoxia-inducible factor 1alpha activity during prolonged hypoxia. Mol Cell Biol. 2011;31(19):4087–96. 10.1128/MCB.01276-10
23. Elton TS, Selemon H, Elton SM, Parinandi NL. Regulation of the MIR155 host gene in physiological and pathological processes. Gene. 2013;532(1):1–12. 10.1016/j.gene.2012.12.009
24. Chiang HR, Schoenfeld LW, Ruby JG, Auyeung VC, Spies N, Baek D, et al. Mammalian microRNAs: Experimental evaluation of novel and previously annotated genes. Genes Dev. 2010;24(10):992–1009. 10.1101/gad.1884710
25. Barker KR, Lu Z, Kim H, Zheng Y, Chen J, Conroy AL, et al. miR-155 modifies inflammation, endothelial activation and blood–brain barrier dysfunction in cerebral malaria. Mol Med. 2017;23:24–33. 10.2119/molmed.2016.00139
26. Chen S, Shan J, Niu W, Lin F, Liu S, Wu P, et al. Micro RNA-155 inhibitor as a potential therapeutic strategy for the treatment of acute kidney injury (AKI): A nanomedicine perspective. RSC Adv. 2018;8(29):15890–6. 10.1039/c7ra13440a
27. Butovsky O, Jedrychowski MP, Cialic R, Krasemann S, Murugaiyan G, Fanek Z, et al. Targeting miR-155 restores abnormal microglia and attenuates disease in SOD1 mice. Ann Neurol. 2015;77(1):75–99. 10.1002/ana.24304
28. Zhang Y, Zhang M, Li X, Tang Z, Wang X, Zhong M, et al. Silencing microRNA-155 attenuates cardiac injury and dysfunction in viral myocarditis via promotion of M2 phenotype polarization of macrophages. Sci Rep. 2016;6:22613. 10.1038/srep22613
29. Wang W, Bian H, Li F, Li X, Zhang D, Sun S, et al. HBeAg induces the expression of macrophage miR-155 to accelerate liver injury via promoting production of inflammatory cytokines. Cell Mol Life Sci. 2018;75(14):2627–41. 10.1007/s00018-018-2753-8
30. Goncalves-Alves E, Saferding V, Schliehe C, Benson R, Kurowska-Stolarska M, Brunner JS, et al. MicroRNA-155 controls T helper cell activation during viral infection. Front Immunol. 2019;10:1367. 10.3389/fimmu.2019.01367
31. Garg A, Seeliger B, Derda AA, Xiao K, Gietz A, Scherf K, et al. Circulating cardiovascular microRNAs in critically ill COVID-19 patients. Eur J Heart Fail. 2021;23(3):468–75. 10.1002/ejhf.2096
32. Abbasi-Kolli M, Sadri Nahand J, Kiani SJ, Khanaliha K, Khatami A, Taghizadieh M, et al. The expression patterns of MALAT-1, NEAT-1, THRIL, and miR-155-5p in the acute to the post-acute phase of COVID-19 disease. Braz J Infect Dis. 2022;26(3):102354. 10.1016/j.bjid.2022.102354
33. Keikha R, Jebali A. [The miRNA neuroinflammatory biomarkers in COVID-19 patients with different severity of illness]. Neurologia (Engl Ed). 2021,1-10. 10.1016/j.nrl.2021.06.005
34. Ahmed FF, Reza MS, Sarker MS, Islam MS, Mosharaf MP, Hasan S, et al. Identification of host transcriptome-guided repurposable drugs for SARS-CoV-1 infections and their validation with SARS-CoV-2 infections by using the integrated bioinformatics approaches. PLoS One. 2022;17(4):e0266124. 10.1371/journal.pone.0266124
35. Vasuri F, Ciavarella C, Collura S, Mascoli C, Valente S, Degiovanni A, et al. Adventitial microcirculation is a major target of SARS-CoV-2-mediated vascular inflammation. Biomolecules. 2021;11(7):1063. 10.3390/biom11071063
36. Molinero M, Benítez ID, González J, Gort-Paniello C, Moncusí-Moix A, Rodríguez-Jara F, et al. Bronchial aspirate-based profiling identifies microRNA signatures associated with COVID-19 and fatal disease in critically ill patients. Front Med (Lausanne). 2021;8:756517. 10.3389/fmed.2021.756517
37. Donyavi T, Bokharaei-Salim F, Baghi HB, Khanaliha K, Alaei Janat-Makan M, Karimi B, et al. Acute and post-acute phase of COVID-19: Analyzing expression patterns of miRNA-29a-3p, 146a-3p, 155-5p, and let-7b-3p in PBMC. Int Immunopharmacol. 2021;97:107641. 10.1016/j.intimp.2021.107641
38. Qi M, Liu B, Li S, Ni Z, Li F. Construction and investigation of competing endogenous RNA networks and candidate genes involved in SARS-CoV-2 infection. Int J Gen Med. 2021;14:6647–59. 10.2147/IJGM.S335162
39. Chow JT, Salmena L. Prediction and analysis of SARS-CoV-2-targeting microRNA in human lung epithelium. Genes (Basel). 2020;11(9):1002. 10.3390/genes11091002
40. Mahesh G, Biswas R. MicroRNA-155: A master regulator of inflammation. J Interferon Cytokine Res. 2019;39(6):321–30. 10.1089/jir.2018.0155
41. Li S, Duan X, Li Y, Li M, Gao Y, Li T, et al. Differentially expressed immune response genes in COVID-19 patients based on disease severity. Aging (Albany NY). 2021;13(7):9265–76. 10.18632/aging.202877
42. Oh JH, Kim GB, Seok H. Implication of microRNA as a potential biomarker of myocarditis. Clin Exp Pediatr. 2022;65(5):230–8. 10.3345/cep.2021.01802