
Download


ORIGINAL ARTICLE
Decursin alleviates LPS-induced lung epithelial cell injury by inhibiting NF-κB pathway activation
Jiangfeng Zhu, Xiaoping Dong*
General Medicine Ward, The First People’s Hospital of Fuyang Hangzhou, Hangzhou, Zhejiang Province, China
Abstract
Objective: To reveal the possible effects of decursin on viability, oxidative stress, and inflammatory response in lipopolysaccharide (LPS)-treated human bronchial epithelial cells-2B (BEAS-2B) and human pulmonary artery endothelial cells (HPAEC) cells, and revealed the potential mechanisms.
Methods: LPS was used to induce acute lung injury (ALI) in normal human lung epithelial cells, including BEAS-2B and HPAEC cells. Cell viability and apoptosis in response to LPS and decursin in BEAS-2B and HPAEC cells were, respectively, evaluated by MTT colorimetric and terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) assays. The oxidative stress and inflammatory response in LPS-treated BEAS-2B and HPAEC cells were detected by enzyme-linked-immunosorbent serologic assay. In addition, the role of decursin in nuclear -factor-kappa B (NF-κB) activation was analyzed by immunoblot and immunofluorescence assays.
Results: Our data revealed that decursin could alleviate the viability of LPS-induced BEAS-2B and HPAEC cells. Decursin could also reduce LPS-induced oxidative stress in BEAS-2B and HPAEC cells. In addition, it could reduce LPS-induced inflammation in BEAS-2B and HPAEC cells. Mechanically, decursin suppressed the activation of NF-κB pathway.
Conclusion: Decursin suppressed NF-κB pathway, and therefore alleviated ALI.
Key words: acute lung injury (ALI), decursin, oxidative stress, pneumonia, NF-κB pathway
*Corresponding author: Xiaoping Dong, General Medicine Ward, The First People’s Hospital of Fuyang Hangzhou, No. 29 Beihuan Road, Fuchun Street, Fuyang District, Hangzhou City, Zhejiang Province 311400, China. Email address: [email protected]
Received 1 June 2022; Accepted 20 June 2022; Available online 1 January 2023
Copyright: Zhu J and Dong X
License: This open access article is licensed under Creative Commons Attribution 4.0 International (CC BY 4.0). http://creativecommons.org/licenses/by/4.0/
Introduction
Acute lung injury (ALI), a serious threat to human health, is a disease with high morbidity and mortality.1 ALI has been provided more and more attention in previous studies.2 The main manifestation of ALI is injury to the alveolar capillary system.3 Previous evidences suggest that inflammation and oxidative stress are associated with ALI.4 Activated neutrophils spread inflammation and injury by producing reactive oxygen species (ROS) and proteolytic enzymes. Lipopolysaccharide (LPS) is the main component of endotoxin, and is considered to be the most important pathogen leading to lung inflammation and sepsis.5 LPS activates a large number of inflammatory cells and induces pneumonia through a variety of signaling pathways.
Decursin, an active component of Angelica gigas, has a variety of biological effects, including relieving antioxidative stress and anti-nerve pain, and anticancer activities.6–8 In terms of mechanism, imperatorin, a furocoumarin and phytochemical, inhibits osteoclast formation and bone loss in vitro and in vivo by blocking receptor activator of nuclear factor kappa-Β ligand (RANKL)-triggered mitogen-activated protein kinase (MAPK) phosphorylation and inhibiting expression of c-FOS and nuclear factor of activated T-cells, cytoplasmic 1 (NFATC1) proteins.9 Furthermore, decursin attenuates inflammatory response by regulating MAPK and nuclear factor-kappa B (NF-κB) pathways.10 There are some reports mentioning beneficial anti-inflammatory functions of other herbs, such as Urtica dioica, and their extracts which have been widely used for treating chronic diseases.11,12 Further studies are required to explore the therapeutic effects of decursin in LPS-induced lung epithelial cell injury.
In recent years, studies on the pathogenesis of ALI have shifted to the occurrence, development, and regulation of inflammation.13 Stimulation, such as trauma and microbial infection, can abnormally increase its activity, leading to excessive release of a large number of proinflammatory cytokines.14 In ALI, the activation of NF-κB signal transduction pathway participates in the expression of various inflammatory factors, and regulates the development of ALI.
In this study, the possible effects of decursin on the progression of ALI were investigated. The data revealed that decursin alleviated LPS-induced pneumonia cell viability, decreased oxidative stress and inflammatory cytokine secretion, and inhibited activation of NF-κB pathway. Hence, it is believed that decursin could serve as a drug of choice for treating ALI.
Materials and Methods
Cell culture
The human bronchial epithelial cells (BEAS-2B), and human pulmonary artery endothelial cells (HPAEC) were obtained from American Type Culture Collection (ATCC, Manassas, VA, USA) and cultured in Dulbecco's Modified Eagle Medium (DMEM; Sigma-Aldrich, St. Louis, MO, USA) supplemented with 10% fetal bovine serum (FBS; HyClone, Logan, UT, USA), 2-mM glutamine, and 100-U/mL penicillin/streptomycin (Sigma-Aldrich) in 5% CO2 humidified atmosphere at 37°C. For LPS and decursin (Sigma-Aldrich) stimulation, cells were treated with 5-μg/mL LPS and 10- and 20-μM decursin.
Cell viability
BEAS-2B and HPAEC cells were plated at a density of 3 × 103 cells/well into 96-well plates. After respective treatment, the cell viability was assessed by adding MTT colorimetric solution. Cells were incubated for another 4 h, and insoluble pellets were dissolved with dimethyl sulfoxide (DMSO) before measuring optical density (OD) at 450-nm wavelength.
Terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) Staining
After indicated treatment, BEAS-2B and HPAEC cells were fixed with formaldehyde, rinsed with phosphate--buffered saline (PBS) solution, and stained with cell apoptosis detection kit (Roche Molecular Biochemicals, Mannheim, Germany). The cell apoptosis was measured using microscope (Olympus, Tokyo, Japan). The apoptotic cells were counted manually.
Detection of superoxide dismutase (SOD), -malondialdehyde (MDA), glutathione (GSH), and myeloperoxidase (MPO)
After indicated treatment, BEAS-2B and HPAEC cells were collected for detecting MDA, SOD, GSH, and MPO with relevant commercial kits (Jiancheng Bioengineering Institute of Nanjing, Jiangsu Province, China). Cells were homogenized and centrifuged (1000 g) for 20 min, and the supernatant was collected. Then the samples were added. The sample was gently shaken, mixed, and covered for reaction to complete at 37°C for 2 h. Immediately, a microplate reader was used to detect the OD of each well at a wavelength of 450 nm. The experiment was repeated for three times.
Enzyme-linked-immunosorbent serologic assay (ELISA)
Concentration of tumor necrosis factor-α (TNF-α), interleukin-1β (IL-1β), and interleukin-6 (IL-6) in cell lysates was measured with ELISA kit following the protocols.
Real-time polymerase chain reaction (PCR)
Cellular total RNA was extracted with TRIzol reagents (Thermo, Rockford, USA). Total RNA was reverse-transcribed into complementary DNA (cDNA) using Moloney murine leukemia virus (M-MLV) reverse transcriptase (RT) (Promega Corporation, Madison, WI, USA). cDNA was amplified using the following primers: TNF-α:GGTGCCTATGTCTCAGCCTCTT, GCCATAGAACTGATGAGAGGGAG; IL-1β: ACAAGGAGAAGAA- AGTAATGAC, GCTGTAGAGTGGGCTTAT; IL-6: AGACAGCCA- CTCACC, TTCTGCCAGTGCCTCTT; caspase-1: ATGGCCGACA- AGGTCCTGA, TTTAATG TCCTGGGAAGAGGTAGA; and glyceraldehyde 3-phosphate dehydrogenase (GAPDH): AGAAGGCTGGGGCTCATTTG, AGGGGCCATCCACAGTCTTC. The PCR reactions were performed on an ABI 7500 Fast system (Life Technologies, Carlsbad, CA, USA). The reaction procedures were as follows: In 40 cycles, initial denaturation at 95°C for 3 min, denaturation at 95°C for 15 s, annealing and extension at 60°C for 30 s. The relative expression was calculated with the 2-ΔΔCq method.
Western blot analysis
Proteins were extracted with radioimmunoprecipitation assay (RIPA) buffer (Beyotime, Shanghai, China). Then the samples were collected and electrophoresed by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE), transferring onto polyvinylidene fluoride (PVDF) membranes, and blocked with 5% fat-free milk. Subsequently, membranes were incubated with primary antibodies targeting nuclear factor erythroid 2-related factor 2 (Nrf2) (mouse, 1:1000; Abcam, Cambridge, MA, USA), NQO-1 (mouse, 1:1000, Abcam), HO-1 (mouse, 1:1000, Abcam), p-p65 (mouse, 1:1000, Abcam), p65 (mouse, 1:1000, Abcam), p-IκBα (rabbit, 1:1000, Abcam), IκBα (rabbit, 1:1000, Abcam), and β-actin (mouse, 1:10,000, Abcam) for 1 h. Ultimately, the membranes were conjugated with anti-mouse and anti-rabbit immunoglobulin G (IgG) (Abcam) for 1 h. Specific proteins were visualized with enhanced chemiluminescence detection kit (ECL, Thermo, Rockford, USA). Finally, immunoblots were quantified by the ImageJ software.
Immunofluorescence
After indicated treatment, cells were fixed and permeabilized with phosphate-buffered saline solution with a low-concentration detergent solution (PBST). Then the cells were cultured with 2% bovine serum albumin (BSA) in PBST for 1 h, followed by incubation with p65 antibody (1:200 dilution) at 4°C overnight. After washing, cells were cultured with fluorescent secondary antibody at room temperature for 1 h. Finally, slices were counterstained with 4β,6-diamidino-2-phenylindole (DAPI) and mounted with anti-quenching agents before observed with a confocal fluorescent microscope.
Statistics
GraphPad 6.0 was used for statistical analysis. Three replicates were performed for each experiment. One-way ANOVA and Turkey’s post-hoc analysis were used for statistical comparisons; P ˂ 0.05 was considered as significant.
Results
Decursin promotes the viability of BEAS-2B and HPAEC cells exposed to LPS
In order to evaluate the role of decursin in the viability of cells exposed to LPS, MTT assay was performed. BEAS-2B and HPAEC cells were stimulated with 5-μg/mL LPS or decursin alone, and cell viability decreased and increased, respectively (Figures 1A and B). Cells stimulated with LPS were then cotreated with increasing doses of decursin. The results demonstrated that addition of decursin improved the viability of cells decreased by LPS (Figure 1B). In addition, LPS significantly induced the percentage of apoptotic cells. Cotreatment of decursin relieved the extent of cell apoptosis caused by LPS (Figure 1C). Collectively, decursin promotes the viability of BEAS-2B and HPAEC cells exposed to LPS.
Figure 1 Decursin promotes the viability of BEAS-2B and HPAEC cells exposed to LPS. (A) Structure of decursin. (B) Cell viability of BEAS-2B and HPAEC cells exposed to LPS or/and decursin. (C) Cell apoptosis as detected by TUNEL staining exposed to LPS or/and decursin. *P ˂ 0.05, **P ˂ 0.01, and ***P ˂ 0.001.
Decursin relieves oxidative stress in BEAS-2B and HPAEC cells exposed to LPS
In order to further reveal the role of decursin in oxidative stress, levels of SOD, MDA, GSH, and MPO were analyzed in BEAS-2B and HPAEC cells exposed to LPS. Induction of MDA and MPO, and reduction of SOD and GSH in LPS group were observed. However, treatment of decursin reversed the altered levels of SOD, MDA, GSH, and MPO in both BEAS-2B and HPAEC cells exposed to LPS (Figure 2A). Moreover, levels of Nrf2, NQO-1, and HO-1 were inhibited by LPS, but enhanced by decursin treatment (Figure 2B). These results suggested that decursin reduced oxidative stress in BEAS-2B and HPAEC cells exposed to LPS.
Figure 2 Decursin relieves oxidative stress in BEAS-2B and HPAEC cells exposed to LPS. (A) Levels of SOD, GSH, MDA, and MPO in control, LPS, decursin + LPS in BEAS-2B and HPAEC cells. (B) Protein levels of Nrf2, NQO-1, and HO-1 in control, LPS, decursin + LPS in BEAS-2B and HPAEC cells. *P ˂ 0.05, **P ˂ 0.01, and ***P ˂ 0.001.
Decursin inhibits LPS-induced inflammation in BEAS-2B and HPAEC cells
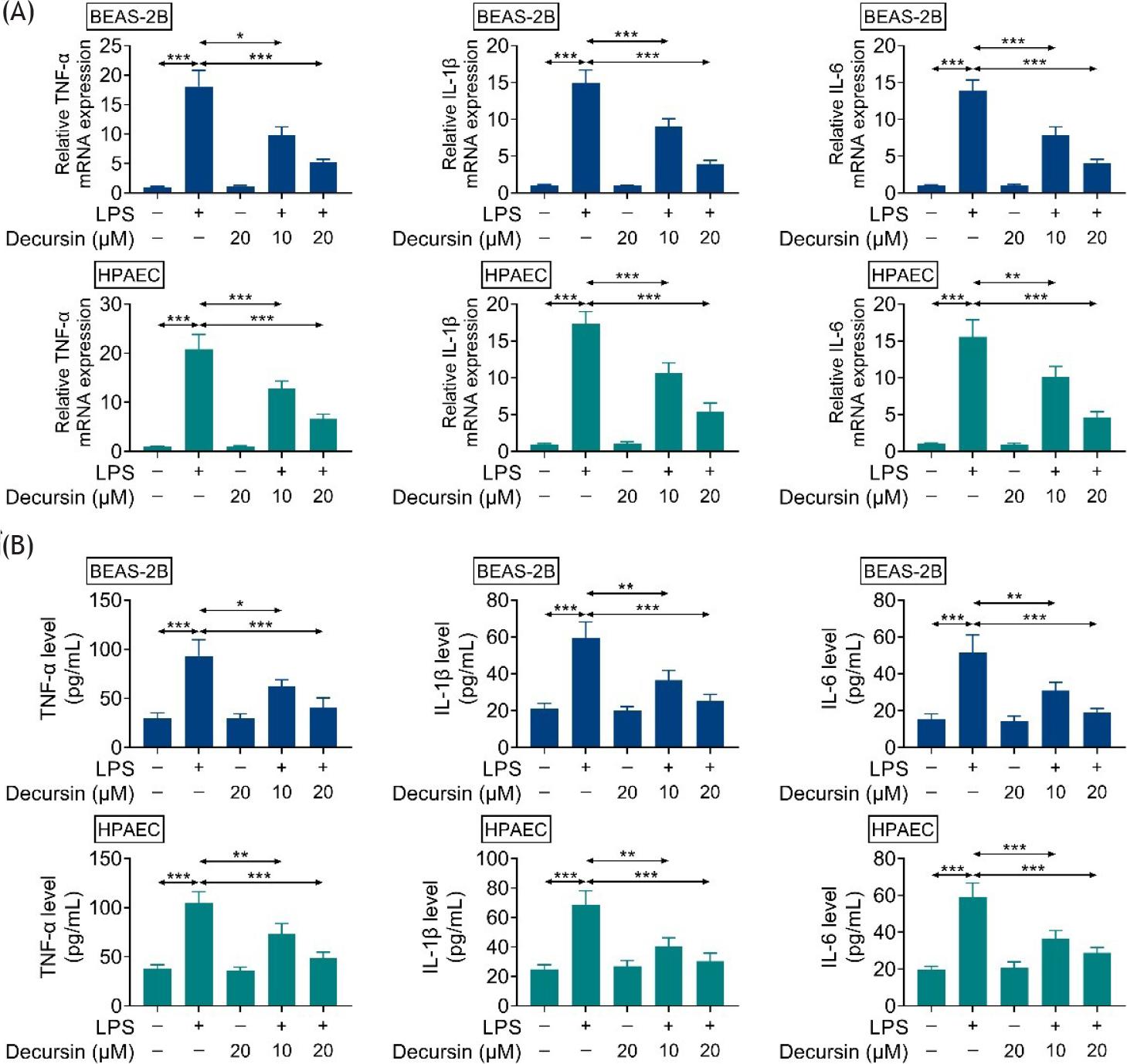
In order to explore inflammatory response in LPS-treated cells, the mRNA levels of IL-6, IL-1β, and TNF-α were monitored. LPS significantly induced the mRNA levels of IL-6, IL-1β, and TNF-α (Figure 3A). Administration of decursin relieved the cellular inflammation as identified by the reduction of these cytokines (Figure 3A). Similar results were found by observing the protein levels of IL-6, IL-1β, and TNF-α after treatment with decursin (Figure 3B). Therefore, it is speculated that decursin inhibits LPS-induced inflammation in BEAS-2B and HPAEC cells.
Figure 3 Decursin inhibits LPS-induced inflammation in BEAS-2B and HPAEC cells. (A) The mRNA levels of TNF-N, IL-1β, and IL-6 in BEAS-2B and HPAEC cells of control, LPS, and decursin + LPS groups. (B) Levels of TNF-N, IL-1β, and IL-6 in BEAS-2B and HPAEC cells of control, LPS, and decursin + LPS cells. *P ˂ 0.05, **P ˂ 0.01, and ***P ˂ 0.001.
Decursin inactivates NF-κB pathway in BEAS-2B and HPAEC cells exposed to LPS
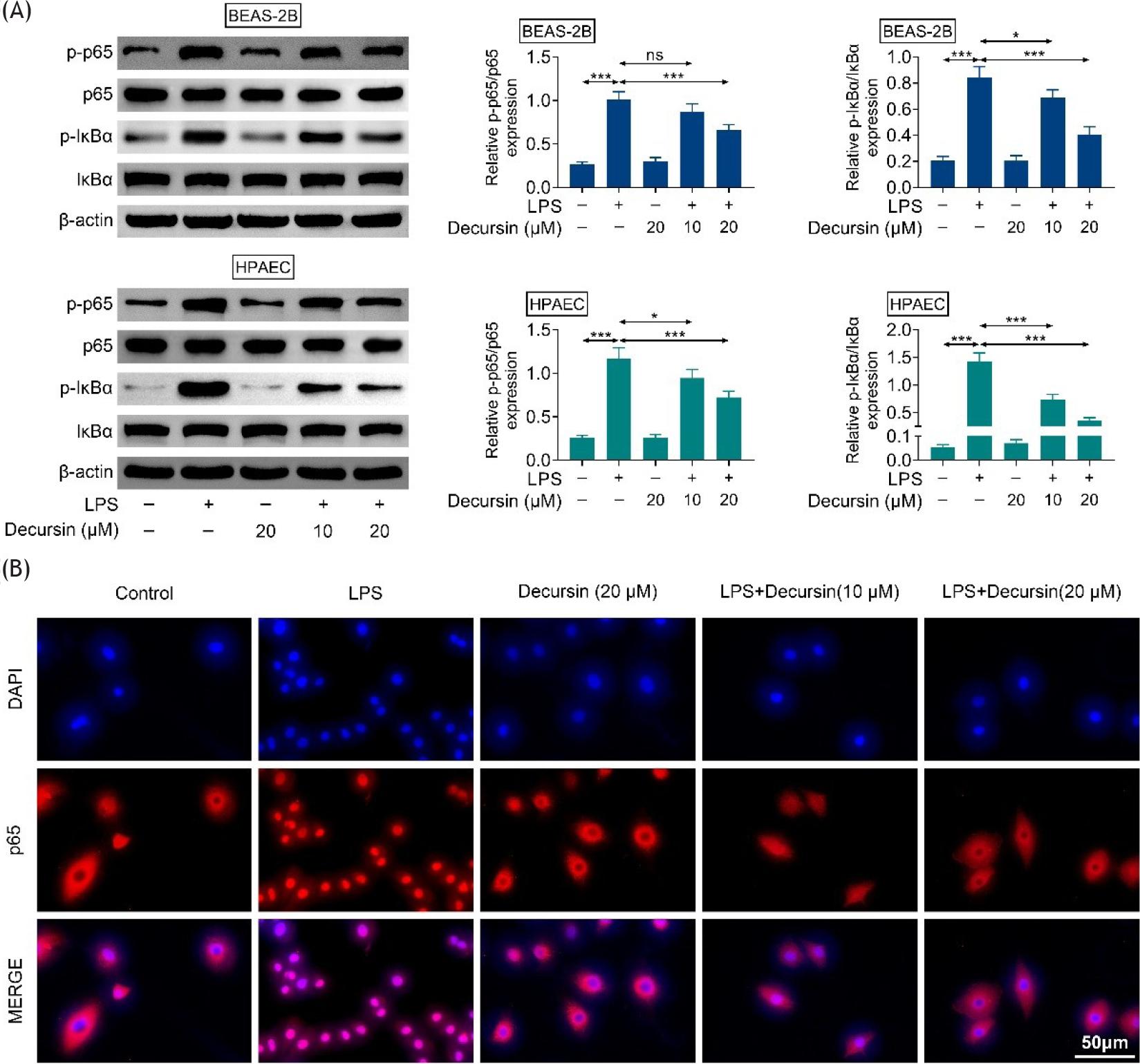
The following study intends to validate the involvement of NF-κB pathway in the functioning of decursin in mediating cell inflammation. LPS increased the levels of p-p65 and p-IκBα, and these alterations were abrogated by decursin treatment (Figure 4A). Moreover, the cytoplasm and nuclear translocation of p65 were changed in different stimuli. LPS significantly induced nuclear accumulation of p65, while decursin repressed the nuclear translocation of p65, suggesting the inactivation of NF-κB pathway (Figure 4B). Therefore, decursin inactivates NF-κB pathway in BEAS-2B and HPAEC cells exposed to LPS.
Figure 4 Decursin inactivates NF-κB pathway in BEAS-2B and HPAEC cells exposed to LPS. (A) Protein levels of p-p65, and p-IκBα in BEAS-2B and HPAEC cells of control, LPS, and decursin + LPS groups. (B) The cellular localization of p65 in BEAS-2B and HPAEC cells of control, LPS, and decursin + LPS groups. *P ˂ 0.05, **P ˂ 0.01, and ***P ˂ 0.001.
Discussion
Acute lung injury is a type of acute hypoxic respiratory failure that results from injury to alveolar epithelial cells.15 The treatment of ALI is primarily etiological.16 The main factors leading to ALI include pneumonia, severe sepsis, and gastroesophageal reflux. In order to combat ALI, more effective drugs are required.17 The present study revealed that decursin could alleviate the activity of LPS-induced pneumonia cells, indicating its potential role as a drug of choice for treating ALI.
Cell counting kit-8 (CCK-8) and flow cytometry (FCM) assays revealed that decursin could alleviate the activity of LPS-induced pneumonia cells. Further, ELISA and immunoblot assay found that decursin could decrease oxidative stress and inflammatory cytokine secretion in LPS-induced pneumonia cells. Therefore, these results suggested that decursin could serve as a drug for treating ALI. In fact, the biological activities of decursin have been revealed widely.18 Decursin has antioxidative and anti-inflammation effects in several diseases.19 Decursin inhibits osteoclast formation and bone loss both in vitro and in vivo.20 Decursin could also prevent melanogenesis via inhibiting melanocyte-inducing transcription factor (MITF) expression. It restrained cell growth as well as autophagic flux in gastric cancer by suppression of athepsin C expression. Decursin could alleviate the aggravation of osteoarthritis by suppressing PI3K-Akt and NF-kB pathways.21 Similarly, the present study revealed that decursin suppressed ALI by inhibiting activation in NF-κB pathway.
Notably, pneumonia results in the release of proinflammatory cytokines, which resulted in ALI. Inflammation and injury by production of ROS also contributed to the progression of ALI. The present study also revealed that decursin suppressed ALI by suppressing inflammatory and oxidative stress. In addition, LPS is the main component of endotoxins and considered as the most important pathogen leading to lung inflammation and sepsis. It could activate a large number of inflammatory cells to stimulate pneumonia. Therefore, an LPS-induced BEAS-2B cell model was successfully constructed to clarify the role of decursin in the progression of ALI in vitro.
A previous study indicated that decursin affected MAPK and NF-κB pathways, and therefore attenuated amyloid-β-induced inflammatory response in PC12 cells. The stated study also found that decursin could attenuate ALI by targeting this pathway.10 NF-κB pathway is critical in mediating cell oxidative stress and inflammatory cytokine secretion, both of which are involved in the progression of ALI. Several proteins and drugs affected the progression of ALI via NF-κB pathway. For example, Baicalin could inhibit inflammation of LPS-induced ALI through NF-κB pathway.22,23 In addition, Tanshinone IIA (Tan IIA) could improve sepsis-induced ALI by mediating ROCK2–NF-κB axis.24 These studies, together with our findings, confirmed that NF-κB pathway could serve as a target for treating ALI.
Conclusion
This study revealed that decursin could alleviate the activity of LPS-induced pneumonia cells. Decursin could also reduce LPS-induced oxidative stress and inflammation in pneumonia cells. Mechanically, decursin suppressed NF-κB pathway and, therefore, alleviated ALI. It is therefore thought that decursin could serve as a drug of choice for treating ALI.
Competing Interests
The authors state that there were no conflicts of interest to disclose.
Ethics Approval
Ethical approval was obtained from the Ethics Committee of The First Hospital of Jilin University.
Data Availability
The authors declare that all data supporting the findings of this study are available in the paper; any raw data can be obtained from the corresponding author upon request.
Author Contributions
Jiangfeng Zhu and Xiaoping Dong designed and carried out the study. Both authors supervised data collection, analysis, and interpretation. Both prepared and reviewed the draft of manuscript for publication. Lastly, both authors read and approved the final manuscript.
REFERENCES
1. Song J, Chen D, Pan Y, Shi X, Liu Q, Lu X, et al. Discovery of a novel MyD88 inhibitor M20 and its protection against sepsis-mediated acute lung injury. Front Pharmacol. 2021;12:775117. 10.3389/fphar.2021.775117
2. Jin A, Zhao Y, Yuan Y, Ma S, Chen J, Yang X, et al. Single treatment of vitamin D3 aAmeliorates LPS-induced acute lung injury through changing lung rodentibacter abundance. Mol Nut Food Res. 2021;66(3):e2100952. 10.1002/mnfr.202100952
3. Engeroff P, Belbezier A, Monsel A, Klatzmann D. Anakinra reduces lung inflammation in experimental acute lung injury. Immun Inflamm Dis. 2022;10(2):123–9. 10.1002/iid3.548.
4. Chen X, Sun Z, Zhang H, Wang L. Correlation of impaired NF-kB activation in sepsis-induced acute lung injury (ALI) in diabetic rats. J Healthc Eng. 2021;2021:5657284. 10.1155/2021/5657284
5. Yang L, Zhao L, Zhang H, Chen P. Up-regulation of TUG1 can regulate miR-494/PDK4 axis to inhibit LPS-induced acute lung injury caused by sepsis. Am J Transl Res. 2021;13(11):12375–85. PMid: 34956459;
6. Kim J, Yun M, Kim EO, Jung DB, Won G, Kim B, et al. Decursin enhances TRAIL-induced apoptosis through oxidative stress-mediated endoplasmic reticulum stress signalling in non-small cell lung cancers. Br J Pharmacol. 2016;173(6):1033–44. 10.1111/bph.13408
7. Kim BS, Seo H, Kim HJ, Bae SM, Son HN, Lee YJ, et al. Decursin from Angelica gigas nakai inhibits B16F10 melanoma growth through induction of apoptosis. J Med Food. 2015;18(10):1121–7. 10.1089/jmf.2014.3397
8. Choi HS, Cho SG, Kim MK, Kim MS, Moon SH, Kim IH, et al. Decursin in Angelica gigas Nakai (AGN) enhances doxorubicin chemosensitivity in NCI/ADR-RES ovarian cancer cells via inhibition of P-glycoprotein expression. Phytother Res (PTR). 2016;30(12):2020–6. 10.1002/ptr.5708
9. Lee TK, Kang IJ, Sim H, Lee JC, Ahn JH, Kim DW, et al. Therapeutic effects of decursin and Angelica gigas nakai root extract in gerbil brain after transient ischemia via protecting BBB leakage and astrocyte endfeet damage. Molecules. 2021;26(8):2161. 10.3390/molecules26082161
10. Kim S, Lee SI, Kim N, Joo M, Lee KH, Lee MW, et al. Decursin inhibits cell growth and autophagic flux in gastric cancer via suppression of cathepsin C. Am J Cancer Res. 2021;11(4):1304–20. PMid: 33948359;
11. Rahmati M, Keshvari M, Mirnasouri R, Chehelcheraghi F. Exercise and Urtica dioica extract ameliorate hippocampal insulin signaling, oxidative stress, neuroinflammation, and cognitive function in STZ-induced diabetic rats. Biomed Pharmacother. 2021;139:111577. 10.1016/j.biopha.2021.111577
12. Keshvari M, Rahmati M, Mirnasouri R, Chehelcheraghi F. Effects of endurance exercise and Urtica dioica on the functional, histological and molecular aspects of the hippocampus in STZ-Induced diabetic rats. J Ethnopharmacol. 2020;256:112801. 10.1016/j.jep.2020.112801
13. Song YR, Jang B, Lee SM, Bae SJ, Bak SB, Kim YW. Angelica gigas NAKAI and its active compound, decursin, inhibit cellular injury as an antioxidant by the regulation of AMP-activated protein kinase and YAP signaling. Molecules. 2022;27(6):1858. 10.3390/molecules27061858
14. Son DB, Choi W, Kim M, Go EJ, Jeong D, Park CK, et al. Decursin alleviates mechanical allodynia in a paclitaxel--induced neuropathic pain mouse model. Cells. 2021;10(3):547. 10.3390/cells10030547
15. Wu L, Cheng Y, Peng S, Zhang W, Zhang C. Sphingosine kinase 1 plays an important role in atorvastatin-mediated anti--inflammatory effect against acute lung injury. Mediators Inflamm. 2021;2021:9247285. 10.1155/2021/9247285
16. Techarang T, Jariyapong P, Viriyavejakul P, Punsawad C. High mobility group box-1 (HMGB-1) and its receptors in the pathogenesis of malaria-associated acute lung injury/acute respiratory distress syndrome in a mouse model. Heliyon. 2021; 7(12):e08589. 10.1016/j.heliyon.2021.e08589.
17. Tang L, Song D, Qi R, Zhu B, Wang X. Roles of pulmonary telocytes in airway epithelia to benefit experimental acute lung injury through production of telocyte-driven mediators and exosomes. Cell Biol Toxicol. 2022;1–15. 10.1007/s10565-021-09670-5. Online ahead of print.
18. Park IS, Kim B, Han Y, Yang H, Cho U, Kim SI, et al. Decursin and decursinol angelate suppress adipogenesis through activation of beta-catenin signaling pathway in human visceral adipose-derived stem cells. Nutrients. 2019;12(1):13. 10.3390/nu12010013
19. Oh JM, Lee HS, Baek SC, Lee JP, Jeong GS, Paik MJ, et al. Antidepressant-like activities of hispidol and decursin in mice and analysis of neurotransmitter monoamines. Neurochem Res. 2020;45(8):1930–40. 10.1007/s11064-020-03057-4
20. Lee W, Sim H, Choi YJ, Seo JY, Yun MY, Song GY, et al. The decursin analog, CYJ-27, suppresses inflammation via the downregulation of NF-kappaB and STAT-1. J Med Food. 2021;24(8):852–9. 10.1089/jmf.2021.K.0027
21. Lee TK, Kim B, Kim DW, Ahn JH, Sim H, Lee JC, et al. Effects of decursin and angelica gigas nakai root extract on hair growth in mouse dorsal skin via regulating inflammatory cytokines. Molecules. 2020;25(16):3697. 10.3390/molecules25163697
22. Lee SH, Lee JH, Kim EJ, Kim WJ, Suk K, Kim JH, et al. A novel derivative of decursin, CSL-32, blocks migration and production of inflammatory mediators and modulates PI3K and NF-kappaB activities in HT1080 cells. Cell Biol Int. 2012;36(7):683–8. 10.1042/CBI20110257
23. Yang L, Zhang X, Lv X, Yu H. Intensity modulated radiotherapy and brachytherapy for a cervical cancer after renal transplantation. Eur J Gynaecol Oncol. 2017;38(1):162–5. PMid: 29767890.
24. Jerez-Lillo N, Álvarez BL, Gutiérrez JM, Figueroa-Zúñiga JI, Leiva V. A statistical analysis for the epidemiological surveillance of COVID-19 in Chile. Signa Vitae. 2022;18(2):19–30. 10.22514/sv.2021.130