
Download
ORIGINAL ARTICLE
CTRP6 suppresses neutrophil extracellular traps formation to ameliorate sepsis-induced lung injury through inactivation of ERK pathway
Jing Lia, Ruijing Xuanb, Weidong Wua*, Yang Hana, Jiani Guoa, Meixia Yangc
aDepartment of Critical Care Medicine, Shanxi Bethune Hospital, Shanxi Academy of Medical Sciences, Tongji Shanxi Hospital, Third Hospital of Shanxi Medical University, Taiyuan, Shanxi Province, China
bDepartment of Experimental Zoology, Laboratory Animal Center, Shanxi Medical University, Taiyuan, Shanxi Province, China
cDepartment of Emergency, Shanxi Bethune Hospital, Shanxi Academy of Medical Sciences, Tongji Shanxi Hospital, Third Hospital of Shanxi Medical University, Taiyuan, Shanxi Province, China
Abstract
Background: Septic lung injury is associated with excessive neutrophil activation, while neutrophil extracellular traps formation contributes to inflammatory lung injury in sepsis. C1q/tumor necrosis factor–related protein-6 (CTRP6) is a paralog of adiponectin and exerts anti- inflammatory and antioxidant properties. The role of CTRP6 in sepsis-associated inflammatory lung injury was investigated in this study.
Methods: Mice were injected with lipopolysaccharides (LPS) intraperitoneally to establish the mouse sepsis model. They were first tail-vein injected with adenovirus-mediated overexpression CTRP6 (Ad-CTRP6) and then subjected to the LPS injection. Pathological changes in lungs were detected by hematoxylin and eosin staining. Inflammation cytokine levels in bronchoalveolar lavage fluid were assessed by qRT-PCR and ELISA. Flow cytometry was used to detect the number of neutrophils in bronchoalveolar lavage fluid, and immunofluorescence was performed to assess neutrophil extracellular traps.
Results: Lipopolysaccharides induced pulmonary congestion, interstitial edema, and alveolar wall thickening in the lungs, as well as upregulated lung histology score and wet/dry weight ratio. CTRP6 was reduced in lung tissues of septic mice. Injection with Ad-CTRP6 ameliorated extensive histopathological changes in LPS-induced mice and decreased lung histology score and wet/dry weight ratio. Overexpression of CTRP6 reduced the levels of TNF-α, IL-6, and IL-1β in septic mice. Injection with Ad-CTRP6 also decreased the number of neutrophils and downregulated Cit-H3 and myeloperoxidase polymers in septic mice. Protein expression of p-ERK in septic mice was reduced by overexpression of CTRP6.
Conclusion: CTRP6 attenuated septic lung injury, exerted anti-inflammatory effect, and suppressed neutrophil extracellular traps formation against sepsis through inactivation of extracellular signal-regulated kinase signaling.
Key words: CTRP6, Extracellular Signal-regulated Kinase, Inflammation, Lipopolysaccharide, Lung Injury, Neutrophil Extracellular Traps, Sepsis
*Corresponding author: Weidong Wu, Department of Critical Care Medicine, Shanxi Bethune Hospital, Shanxi Academy of Medical Sciences,Tongji Shanxi Hospital,Third Hospital of Shanxi Medical University, No. 99 Longcheng Dajie, Xiaodian District, Taiyuan City, Shanxi Province, China. Email address: [email protected]
Received 20 May 2022; Accepted 9 June 2022; Available online 1 November 2022
Copyright: Li J, et al.
License: This open access article is licensed under Creative Commons Attribution 4.0 International (CC BY 4.0). http://creativecommons.org/licenses/by/4.0/
Introduction
Sepsis is a life-threatening multi-organ dysfunction disease caused by the host’s disorder of immune response to infection.1 Lung injury and acute respiratory distress syndrome (ARDS) refer to common sepsis-associated multi-organ dysfunction and contribute to increased mortality.2 Each year, about 20 million people suffer from sepsis-associated organ dysfunction worldwide, leading to 5 million deaths.3 (The Global Epidemiology of Sepsis. Does It Matter That We Know So Little?). In China, at least 5.68 million people suffer from sepsis-associated organ dysfunction each year with a mortality of 0.24–0.83 per 100,000 patients.4 (Current epidemiology of sepsis in mainland China).
Neutrophils are the most abundant circulating granulocytes, and represent the first barrier against pathogenic infection.5 Neutrophils eliminate the pathogens through phagocytosis, degranulation, and neutrophil extracellular trap formation.5 Therefore, neutrophils play a critical role in both adaptive and innate immune systems.5 Previous studies have shown that a large number of neutrophils infiltrate into lungs, andwhich promote secretion of pro-inflammatory cytokines, thus damaging the epithelium and endothelium of lungs, resulting in pulmonary edema during the development of sepsis-associated lung injury.6 Suppression of neutrophilic inflammation ameliorated acute lung injury.7 Hub immune genes involved in sepsis-associated organ dysfunction were identified through bioinformatics analysis (Iidentification of key immune genes for sepsis-induced ARDS based on bioinformatics analysis). Therefore, genes involved in neutrophilic immunoreaction might be potential targets for sepsis.
Neutrophil extracellular traps are net-like extracellular structures, composed of DNA, histones, myeloperoxidase (MPO), and neutrophil elastic cell enzymes.5 Neutrophil extracellular traps could trap, neutralize, and kill pathogens to prevent dissemination, while excessive neutrophil extracellular traps could also induce epithelial and endothelial cells during acute lung injury.5,8 Neutrophil extracellular traps stimulated infiltration of neutrophils into lungs and induced tissue injury through secretion of pro- inflammatory cytokines in sepsis-associated lung injury.9,10 Inhibition of neutrophil extracellular traps formation alleviated sepsis-associated lung injury.11
C1q/tumor necrosis factor-related protein-6 (CTRP6) is a paralog of adiponectin that regulates energy metabolism.12 For example, CTRP6 is involved in glucose and fat metabolism, and mediate insulin resistance.13 This protein function as an endogenous complement regulator to inhibit rheumatoid arthritis.14 It reduces myofibroblast differentiation to alleviate post-infarct cardiac fibrosis.15 Moreover, it is also associated with inflammatory responses in adipose tissues.13 CTRP6 reduces oxygen–glucose deprivation and reperfusion-induced oxidative stress and inflammation to attenuate cerebral ischemia/reperfusion injury.16 However, the role of CTRP6 in inflammation of sepsis-associated lung injury remains unknown.
In this study, the effects of CTRP6 on inflammation and neutrophil extracellular traps formation in septic mice were investigated. The results might provide potential target for sepsis.
Materials and Methods
Animal model
Female C57BL/6J mice (n = 24; 8–10 weeks old), weighing 20–25 g, were acquired from Shanghai Slack Laboratory Animal Co., Ltd (Shanghai, China). Mice acquired standard chow and water in a pathogen-free room. This study was approved by the Laboratory Animals Welfare Ethics Committee of Shanxi Bethune Hospital (Shanxi Academy of Medical Sciences), and is in accordance with the National Institute of Health, Laboratory Animal Care and Use Guidelines. Mice were divided into four groups: Sham (n = 6), lipopolysaccharide (LPS; n = 6), LPS + Ad-NC (n = 6), and LPS + Ad-CTRP6 (adenovirus-mediated overexpression CTRP6; n = 6). Mice in the Sham group were intraperitoneally injected with 100 μL sterile saline, and those in the LPS group were injected with 30 mg/kg LPS (Sigma-Aldrich, St. Louis, MO, USA). Ad-NC and Ad-CTRP6 were purchased from Genepharm (Shang, China). Mice were injected with 100 μL adenovirus (107 particles/L) via tail vein 7 days before LPS injection. Mice in each group were anesthetized for 6 h after LPS administration. Lung tissues were collected for histologic examination, and saline was injected into tracheal cannula to collect bronchoalveolar lavage fluid for the analysis of neutrophils.
Histologic examination
Lung tissues were fixed in 10% formalin and embedded in paraffin. The tissues were sliced into 5 μm sections, and these sections were then deparaffinized in toluene and hydrated in gradient concentrations of alcohol. The sections were stained with hematoxylin and eosin (Sigma-Aldrich) and observed under microscope (Olympus, Tokyo, Japan). Lung histology score was evaluated by pathologists that were blind to image information. Five random areas in lung parenchyma were scored based on the following criteria: 0 for absence, 1 for light, 2 for moderate, 3 for strong, and 4 for intense for the four parameters (neutrophil infiltration, alveolar hemorrhage, thickness of alveolar septa, and size of alveolar spaces).
Lung edema
Left lung from each group was gently dried and weighed. The tissues were dried at 60°C for 3 days and weighed. The ratio of wet weight to dry weight was calculated as an indicator of lung edema.
qRT-PCR
Lung tissues were lyzed in Trizol (Invitrogen, Carlsbad, CA, USA) to isolate total RNAs. RNAs were reverse- transcribed into cDNAs, and cDNAs were subjected to qRT-PCR analysis with SYBR Green Master (Roche, Mannheim, Germany). GAPDH (Forward: 5’-GGATTTGGTCGTATTGGG-3’ and Reverse: 5’-GGAAGATGGTGATGGGATT-3’) was used as endogenous control. Expressions of CTRP6 (Forward: 5’-C-C-A -TCCTGAAAGGTGACAAAGG-3’ and Reverse: 5’-AGTAATGCGT CTGGCACGAG-3’), TNF-α (Forward: 5’-ACTGAACTTCGGGGT GATTG-3’ and Reverse: 5’-GCTTGGTGGTTTGCTACGAC-3’), IL-6 (Forward: 5’-TGATGGATGCTTCCAAACTG-3’ and Reverse: 5’-GAGCATTGGAAGTTGGGGTA-3’), and IL-1β (Forward: 5’-CA CCTTCTTTTCCTTCATCTTTG-3’ and Reverse: 5’-GTCGTTGC TTGTCTCTCCTTGTA-3’) were determined by 2-∆∆Ct method.
ELISA
Lung tissues were lyzed in RIPA buffer (Beyotime, Beijing, China), and then centrifuged at 12,000 g for 60 min at 4°C to collect tissue lysates. Levels of TNF-α, IL-6, and IL-1β were determined using commercial ELISA kits (MultiSciences Biotech Co., Ltd., Wuhan, China). The level of MPO in bronchoalveolar lavage fluid was also evaluated using ELISA kit.
Flow cytometry
The isolated bronchoalveolar lavage fluids were filtered with 40 μm filter, and cells were harvested by centrifugation. The cells were incubated with fluorescence-activated cell- sorting buffer containing PE-labeled Ly6G and analyzed under BD flow cytometer (BD Pharmingen, San Diego, CA, USA).
Immunofluorescent staining
Lung sections were incubated in citrate buffer and permeabilized with 0.1% Triton X-100. The sections were blocked in 5% bovine serum albumin and probed with rabbit against mice anti-Cit-H3 (1:100; Abcam, Cambridge, MA, USA) and rabbit against mice anti-MPO (1:100; Abcam) antibodies. The sections were treated with goat anti-rabbit fluorescein isothiocyanate–conjugated IgG and observed under Zeiss LSM 510 Confocal Imaging System (Zeiss, Jena, Germany) with DAPI counterstaining.
Western blot
Supernatant of lung tissues was segregated using SDS-PAGE and transferred onto nitrocellulose membranes. Membranes were blocked in 5% bovine serum albumin and incubated with primary antibodies: anti-CTRP6 and anti- β-actin (1:2000), anti-p-ERK and anti-ERK (1:3000). The membranes were then incubated with secondary antibodies (1:4000), and subjected to chemiluminescence reagent kit (Beyotime). All the proteins were purchased from Abcam.
Statistical Analysis
All data were expressed as mean ± SEM and analyzed by Student’s t test or one-way analysis of variance. P < 0.05 was considered statistically significant.
Results
CTRP6 ameliorated septic lung injury
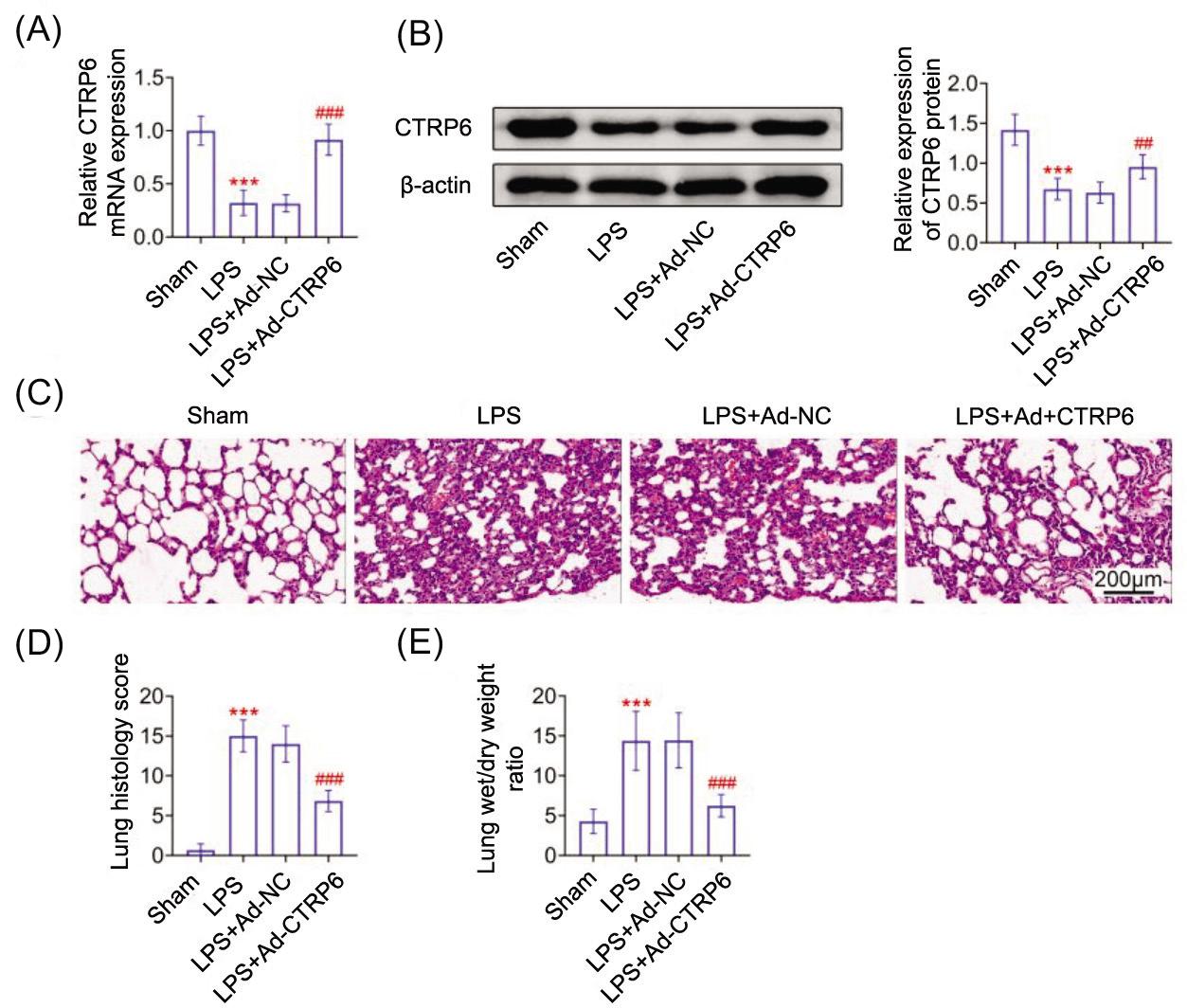
To induce sepsis, mice were injected with 30 mg/kg LPS. Results showed that CTRP6 was downregulated in lungs of septic mice (Figures 1A and 1B). Lipopolysaccharides induced pulmonary congestion, interstitial edema, and alveolar wall thickening in the lungs of septic mice (Figure 1C), as well as upregulated lung histology score (Figure 1D) and wet/dry weight ratio (Figure 1E). Mice were administered Ad-CTRP6 and were subjected to LPS treatment. Injection with Ad-CTRP6 enhanced expression of CTRP6 (Figures 1A and 1B). The overexpression of CTRP6 attenuated extensive histopathological changes in lungs of septic mice (Figure 1C) and reduced lung histology score (Figure 1D) and wet/dry weight ratio (Figure 1E) to protect against sepsis-associated lung injury.
Figure 1 CTRP6 ameliorated septic lung injury. (A) CTRP6 mRNA was downregulated in lungs of septic mice, while injection with Ad-CTRP6 enhanced expression of CTRP6. (B) CTRP6 protein was downregulated in lungs of septic mice, while injection with Ad-CTRP6 enhanced expression of CTRP6. (C) Lipopolysaccharide induced pulmonary congestion, interstitial edema, and alveolar wall thickening in the lungs of septic mice, while overexpression of CTRP6 attenuated extensive histopathological changes in lungs of septic mice. (D) Lipopolysaccharide induced upregulation of lung histology score, while overexpression of CTRP6 reduced lung histology score in septic mice. (E) Overexpression of CTRP6 attenuated lipopolysaccharide-induced increase of wet/dry weight ratio in mice. *** vs Sham, P < 0.001. ### vs Ad-NC, P < 0.001.
CTRP6 ameliorated septic inflammation
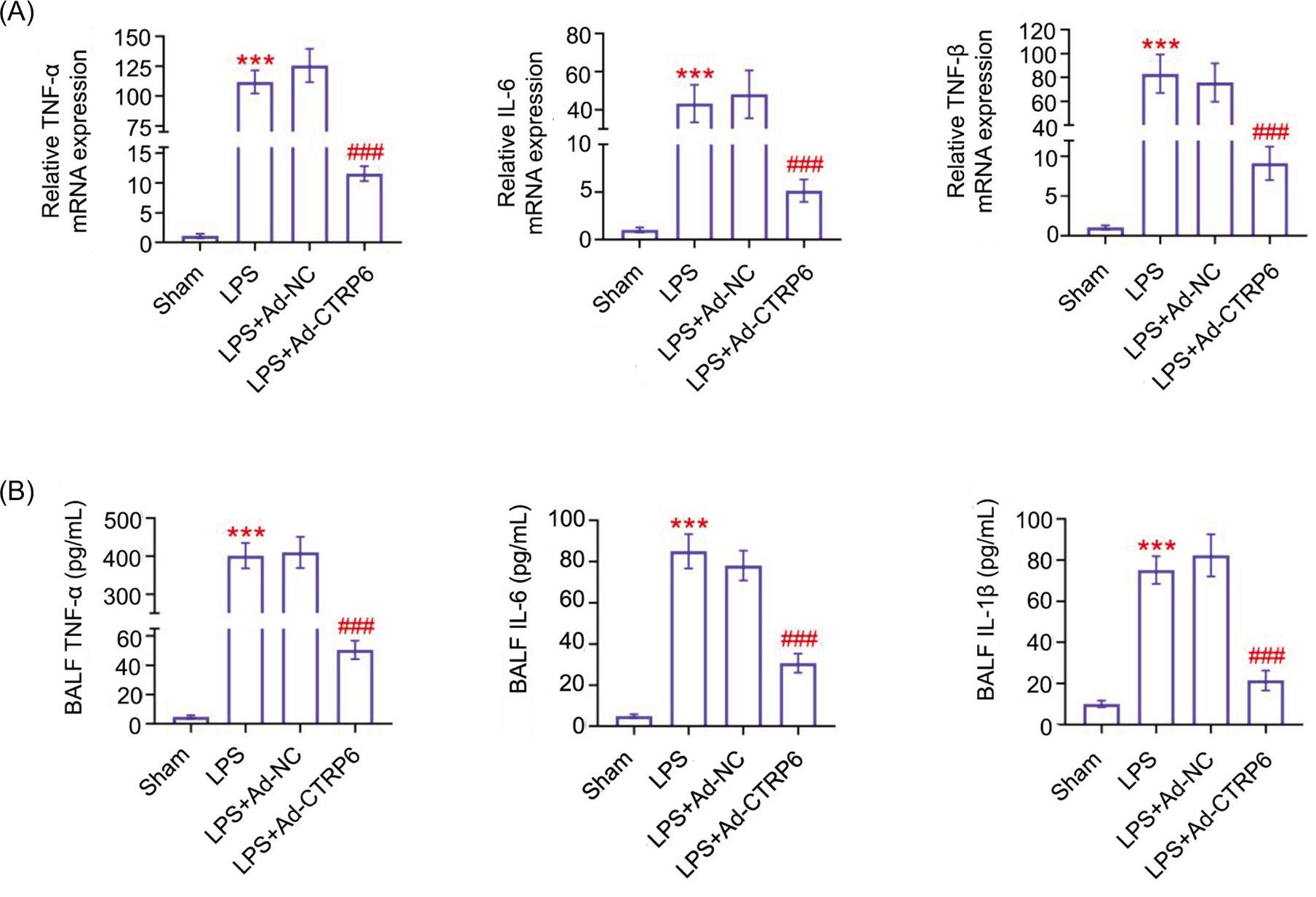
Expression levels of TNF-α, IL-6, and IL-1β were upregulated in lungs of septic mice (Figures 2A and 2B). However, injection with Ad-CTRP6 reduced TNF-α, IL-6, and IL-1β (Figures 2A and 2B) thus inhibiting inflammation in septic mice.
Figure 2 CTRP6 ameliorated septic inflammation. (A) Overexpression of CTRP6 attenuated lipopolysaccharide-induced increase of TNF-α, IL-6, and IL-1β mRNAs in mice. (B) Overexpression of CTRP6 attenuated lipopolysaccharide-induced increase of TNF-α, IL-6, and IL-1β proteins in mice. *** vs Sham, P < 0.001. ### vs Ad-NC, P < 0.001.
CTRP6 suppressed neutrophil extracellular traps formation
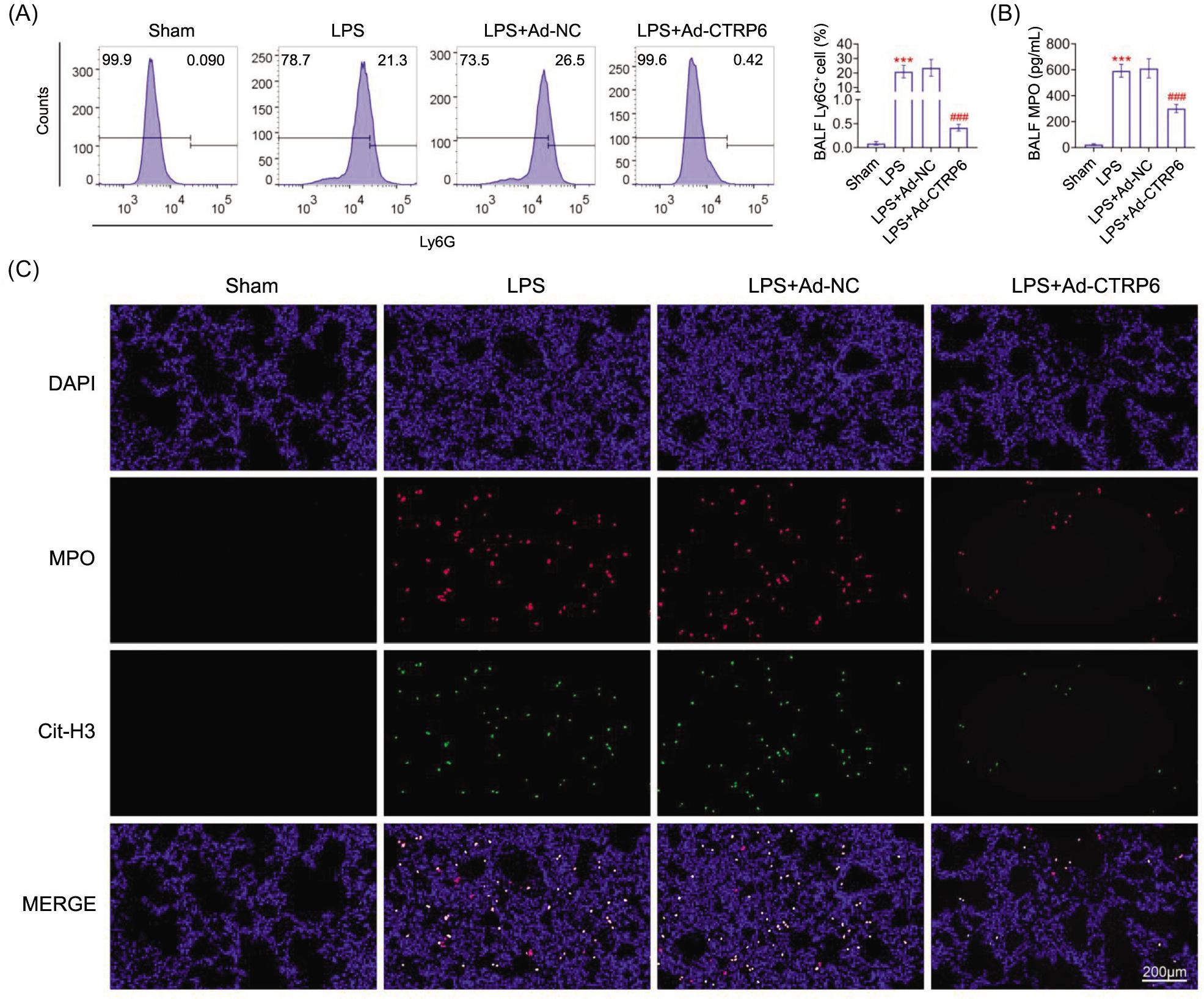
The number of Ly6G+ neutrophils was increased (Figure 3A) and MPO was also upregulated (Figure 3B) in bronchoalveolar lavage fluids of septic mice. Overexpression of CTRP6 reduced the number of Ly6G+ neutrophils (Figure 3A) and decreased the level of MPO (Figure 3B) in septic mice. Lipopolysaccharide induced co-localization of MPO with Cit-H3 in lungs of septic mice (Figure 3C), whereas overexpression of CTRP6 reduced co-staining of MPO and Cit-H3 (Figure 3C) to inhibit neutrophil extracellular traps formation.
Figure 3 CTRP6 suppressed neutrophil extracellular traps formation. (A) Overexpression of CTRP6 attenuated lipo polysaccharide- induced increase of number of Ly6G+ neutrophils in bronchoalveolar lavage fluids of mice. (B) Overexpression of CTRP6 attenuated lipopolysaccharide-induced increase of MPO in bronchoalveolar lavage fluids of mice. (C) Overexpression of CTRP6 attenuated lipo polysaccharide- induced increase of double staining for MPO and Cit-H3 in lungs of mice. ***vs Sham, P < 0.001. ###vs Ad-NC, P < 0.001.
CTRP6 inhibited activation of extracellular signal-regulated kinase (ERK) signaling in septic mice
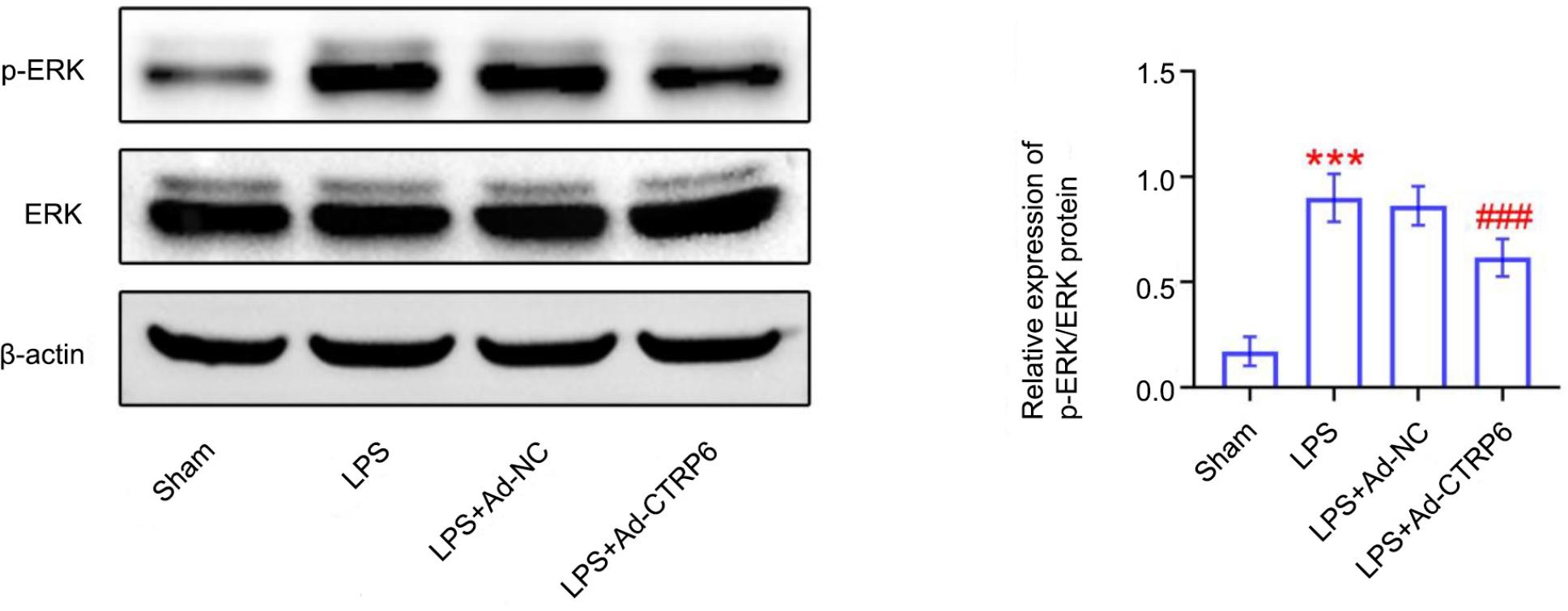
Protein expression of p-ERK was elevated in lungs of septic mice (Figure 4). However, overexpression of CTRP6 reduced p-ERK expression in septic mice (Figure 4) to inhibit activation of ERK signaling.
Figure 4 CTRP6 inhibited activation of ERK signaling in septic mice. Overexpression of CTRP6 attenuated lipopolysaccharide-induced increase of p-ERK expression in lungs of mice. *** vs Sham, P < 0.001. ### vs Ad-NC, P < 0.001.
Discussion
CTRPs were implicated in inflammatory responses.17,18 CTRP3 exerted anti-inflammatory effect to protect against diabetic peripheral neuropathy,19 while CTRP6 deficiency enhanced complement activation and contributed to development of inflammatory diseases.14 This study found that CTRP6 exhibited anti-inflammatory effect to protect against sepsis- associated lung injury and inhibited neutrophil extracellular traps formation.
Previous studies have shown that LPS induced excessive vascular permeability and inflammation of lungs, and LPS-induced mice was widely used as septic models.20 This study also established the septic model through administration with LPS. Lipopolysaccharide stimulated histopathological changes in lungs of mice and enhanced lung edema. CTRP6 was found to be downregulated in lungs of septic mice. Injection with Ad-CTRP6 ameliorated histopathological changes in lungs of septic mice and reduced the lung edema. Therefore, CTRP6 showed protective effect against sepsis-associated lung injury.
Pathogenic infection has been shown to evoke pulmonary inflammation through excessive production of pro- inflammatory cytokines (IL-1β, IL-6, TNF-α) and facilitate for impairment of capillary endothelial barrier during the development of septic lung injury.20 Suppression of alveolar inflammation ameliorated lung injury.21 Results showed that LPS induced upregulation of IL-1β, IL-6, and TNF-α in lungs. However, overexpression of CTRP6 reduced mRNA and protein expression of IL-1β, IL-6, and TNF-α in septic mice, suggesting that CTRP6 exerted anti-inflammatory effect to attenuate sepsis-associated lung injury. Moreover, oxidative stress was involved in pathogenesis of sepsis, and antioxidant therapies showed promising effect in clinical trials of septic patients.22 CTRP6 was reported to reduce oxidative stress to alleviate cerebral ischemia or reperfusion injury.16 Therefore, CTRP6 might exert antioxidant effect to attenuate sepsis-associated lung injury.
Neutrophil extracellular traps exhibited pro-inflammatory effect to aggravate lung injury and pulmonary infiltration of neutrophils during development of sepsis.9 Inhibition of LPS-stimulated neutrophil extracellular traps formation alleviated sepsis-associated lung injury.23 Flow cytometry demonstrated that CTRP6 reduced the number of Ly6G+ neutrophils and decreased the level of MPO in septic mice. Moreover, co-staining of cit-H3 and MPO in lungs of septic mice was also repressed by overexpression of CTRP6. The above results indicated that CTRP6 reduced excessive neutrophil extracellular traps formation to protect against sepsis-associated lung injury.
ERK signaling is implicated in various cellular processes, including cell survival, apoptosis, stress response, motility, differentiation, and proliferation.24 Activation of ERK contributed to septic lung injury by promoting inflammatory response.25 Moreover, ERK signaling was also involved in MPO- mediated activation of neutrophil elastase, promoted chromatin decondensation, and was associated with neutrophil extracellular traps formation.5 Inhibition of ERK phosphorylation ameliorated septic lung injury through suppression of neutrophil extracellular traps formation.26 CTRP6 regulated lipogenesis in myoblasts through ERK signaling27 and reduced phosphorylation of ERK in TGF-β-induced fibroblasts.28 Results of this study demonstrated that overexpression of CTRP6 reduced protein expression of p-ERK in septic lungs, indicating that CTRP6 might attenuate septic lung injury and inhibit neutrophil extracellular traps formation through inactivation of ERK signaling.
Collectively, our results showed that CTRP6 exerted anti-inflammatory effect and reduced neutrophil extracellular traps formation via downregulation of ERK signaling, thus ameliorating septic lung injury. Therefore, CTRP6 might be a potential target for sepsis-associated lung injury.
Acknowledgments
Not applicable.
Funding
This work was supported by the Shanxi Provincial Depart-ment of Science and Technology, Shanxi Provincial Basic Research Program, Free Exploration Youth Scientific Research Project (Grant No. 202103021223407).
Competing Interests
The authors state that there are no conflicts of interest to disclose.
Ethics Approval
Ethical approval was obtained from the Laboratory Animals Welfare Ethics Committee of Shanxi Bethune Hospital (Shanxi Academy of Medical Sciences).
Data Availability
The authors declare that all data supporting the findings of this study are available within the paper, and any raw data can be obtained from the corresponding author upon request.
Authors’ Contributions
Jing Li and Ruijing Xuan designed the experiments, and Weidong Wu and Yang Han carried them out. Jiani Guo analyzed and interpreted the data, and Meixia Yang prepared the manuscript with contributions from all coauthors.
REFERENCES
1. Chuanlong Song AA, Kari A, Abuduhaer A. FSTL1 aggravates sepsis-induced acute kidney injury through regulating TLR4/MyD88/NF-κB pathway in newborn rats. Signa Vitae. 2021;17(3):167–73.
2. Liang W, Guo L, Liu T, Qin S. MEF2C alleviates acute lung injury in cecal ligation and puncture (CLP)-induced sepsis rats by up-regulating AQP1. Allergol Immunopathol. 2021;49(5): 117–24. 10.15586/aei.v49i5.477
3. Finfer S, Machado FR. The global epidemiology of sepsis. Does it matter that we know so little? Am J Respir Crit Care Med. 2016;193(3):228–30. 10.1164/rccm.201510--1976ED
4. Liao X, Du B, Lu M, Wu M, Kang Y. Current epidemiology of sepsis in mainland China. Ann Transl Med. 2016;4(17):324. 10.21037/atm.2016.08.51
5. Papayannopoulos V. Neutrophil extracellular traps in immunity and disease. Nat Rev Immunol. 2018;18(2):134–47. 10.1038/nri.2017.105
6. Grommes J, Soehnlein O. Contribution of neutrophils to acute lung injury. Mol Med. 2011;17(3–4):293–307. 10.2119/molmed.2010.00138
7. Kao T-I, Chen P-J, Wang YH, Tseng HH, Chang SH, Wu TS, et al. Bletinib ameliorates neutrophilic inflammation and lung injury by inhibiting Src family kinase phosphorylation and activity. Br J Pharmacol. 2021;178(20):4069–84. 10.1111/bph.15597
8. Mikacenic C, Moore R, Dmyterko V, West TE, Altemeier WA, Liles WC, et al. Neutrophil extracellular traps (NETs) are increased in the alveolar spaces of patients with ventilator-associated pneumonia. Crit Care. 2018;22(1):358. 10.1186/s13054-018-2290-8
9. Luo L, Zhang S, Wang Y, Rahman M, Syk I, Zhang E, et al. Proinflammatory role of neutrophil extracellular traps in abdominal sepsis. Am J Physiol Lung Cell Mol Physiol. 2014; 307(7):L586–96. 10.1152/ajplung.00365.2013
10. Zhang H, Zhou Y, Qu M, Yu Y, Chen Z, Zhu S, et al. Tissue factor-enriched neutrophil extracellular traps promote immunothrombosis and disease progression in sepsis-induced lung injury. Front Cell Infect Microbiol. 2021;11:677902. 10.3389/fcimb.2021.677902
11. Ding Z, Du F, Averitt V RG, Jakobsson G, Rönnow C-F, Rahman M, et al. Targeting S100A9 reduces neutrophil recruitment, inflammation and lung damage in abdominal sepsis. Int J Mol Sci. 2021;22(23):12923. 10.3390/ijms222312923
12. Dong X, Hu H, Fang Z, Cui J, Liu F. CTRP6 inhibits PDGF-BB-induced vascular smooth muscle cell proliferation and migration. Biomed Pharmacother. 2018;103:844–50. 10.1016/j.biopha.2018.04.112
13. Lei X, Seldin MM, Little HC, Choy N, Klonisch T, Wong GW. C1q/TNF-related protein 6 (CTRP6) links obesity to adipose tissue inflammation and insulin resistance. J Biol Chem. 2017; 292(36):14836–50. 10.1074/jbc.M116.766808
14. Murayama MA, Kakuta S, Inoue A, Umeda N, Yonezawa T, Maruhashi T, et al. CTRP6 is an endogenous complement regulator that can effectively treat induced arthritis. Nat Commun. 2015;6(1):8483. 10.1038/ncomms9483
15. Lei H, Wu D, Wang J-Y, Li L, Zhang C-L, Feng H, et al. C1q/tumor necrosis factor-related protein-6 attenuates post-infarct cardiac fibrosis by targeting RhoA/MRTF-A pathway and inhibiting myofibroblast differentiation. Basic Res Cardiol. 2015;110(4):35. 10.1007/s00395-015-0492-7
16. Mitsis A, Gragnano FJCCR. Myocardial infarction with and without ST-segment elevation: A contemporary reappraisal of similarities and differences. Curr Cardiol Rev. 2021;17(4):e230421189013. 10.2174/1573403X16999201210195702
17. Wang H, Liu Q, Zhang X. C1q/tumor necrosis factor-related protein-1 attenuates microglia autophagy and inflammatory response by regulating the Akt/mTOR pathway. Life Sci. 2020;256:117992. 10.1016/j.lfs.2020.117992
18. Schmid A, Vlacil A-K, Schuett J, Karrasch T, Schieffer B, Schäffler A, et al. Anti-inflammatory effects of C1q/tumor necrosis factor-related protein 3 (CTRP3) in endothelial cells. Cells. 2021;10(8):2146. 10.3390/cells10082146
19. Lin K, Yang L, Xiong Y, Feng K, Zeng W, Deng B. Plasma C1q/tumor necrosis factor-related protein-3 concentrations are associated with diabetic peripheral neuropathy. BMJ Open Diabetes Res Care. 2022;10(2):e002746. 10.1136/bmjdrc-2021-002746
20. Ganapathy-Kanniappan S, Geschwind JFH. Tumor glycolysis as a target for cancer therapy: Progress and prospects. Mol Cancer. 2013;12(1):152. 10.1186/1476-4598-12-152
21. Rentsendorj O, Damarla M, Aggarwal NR, Choi J-Y, Johnston L, D’Alessio FR, et al. Knockdown of lung phosphodiesterase 2A attenuates alveolar inflammation and protein leak in a two-hit mouse model of acute lung injury. Am J Physiol Lung Cell Mol Physiol. 2011;301(2):L161–70. 10.1152/ajplung.00073.2011
22. Prauchner CA. Oxidative stress in sepsis: Pathophysiological implications justifying antioxidant co-therapy. Burns. 2017; 43(3):471–85. 10.1016/j.burns.2016.09.023
23. Wang Y-P, Guo Y, Wen P-S, Zhao Z-Z, Xie J, Yang K, et al. Three ingredients of safflower alleviate acute lung injury and inhibit NET release induced by lipopolysaccharide. Mediators Inflamm. 2020;2020:2720369. 10.1155/2020/2720369
24. Chen Z, Ding X, Jin S, Pitt B, Zhang L, Billiar T, et al. WISP1-αvβ3 integrin signaling positively regulates TLR-triggered inflammation response in sepsis induced lung injury. Sci Rep. 2016;6(1):28841. 10.1038/srep28841
25. Cai X, Chen Y, Xie X, Yao D, Ding C, Chen M. Astaxanthin prevents against lipopolysaccharide-induced acute lung injury and sepsis via inhibiting activation of MAPK/NF-κB. Am J Transl Res. 2019;11(3):1884–94.
26. Yang C, Song C, Liu Y, Qu J, Li H, Xiao W, et al. Re-Du-Ning injection ameliorates LPS-induced lung injury through inhibiting neutrophil extracellular traps formation. Phytomedicine. 2021;90:153635. 10.1016/j.phymed.2021.153635
27. Wu W, Sun Y, Zhao C, Zhao C, Chen X, Wang G, et al. Lipogenesis in myoblasts and its regulation of CTRP6 by AdipoR1/Erk/PPARγ signaling pathway. Acta Biochim Biophys Sin (Shanghai). 2016;48(6):509–19. 10.1093/abbs/gmw032
28. Wang S, Sun Z, Yang S, Chen B, Shi J. CTRP6 inhibits cell proliferation and ECM expression in rat mesangial cells cultured under TGF-β1. Biomed Pharmacother. 2018;97:280–5. 10.1016/j.biopha.2017.10.091