
Download
ORIGINAL ARTICLE
Protopine alleviates lipopolysaccharide-triggered intestinal epithelial cell injury through retarding the NLRP3 and NF-κB signaling pathways to reduce inflammation and oxidative stress
Junyu Lia, Zhongjun Xub*, Canhui OuYanga, Xiongjian Wua, Yun Xiea, Jun Xiea
aDepartment of Gastroenterology, First Affiliated Hospital of Gannan Medical University, Ganzhou, Jiangxi, China
bDepartment of Medical Imaging, First Affiliated Hospital of Gannan Medical University, Ganzhou, Jiangxi, China
Abstract
Background: Inflammatory bowel disease (IBD) is a common chronic intestinal disease. Protopine isolated from different plants has been investigated to understand its special functions on varied diseases. However, the regulatory effects of protopine on the progression of IBD remain unclear. Our study is aimed to explore the effects of protopine on the progression of IBD and its underlying regulatory mechanism of action.
Methods: The cell viability was assessed through MTT colorimetric assay. The protein expressions of genes were examined by Western blot analysis. The cell apoptosis and reactive oxygen species level were measured using flow cytometry. The levels of inflammation and oxidative stress-related proteins were tested through enzyme-linked-immunosorbent serologic assay. The intracellular Ca2+ concentration and mitochondrial membrane potential were measured through immunofluorescence assay.
Results: First, different concentrations of lipopolysaccharide (LPS) were treated with NCM460 cells to establish IBD cell model, and 5-μg/mL LPS was chosen for followed experiments. In this study, we discovered that protopine relieved the LPS-induced inhibited intestinal epithelial cell viability and enhanced cell apoptosis. Moreover, protopine attenuated LPS-stimulated inflammation activation and oxidative stress. Further experiments illustrated that the increased intracellular Ca2+ concentration and decreased mitochondrial membrane potential stimulated by LPS were reversed by protopine treatment. Finally, through Western blot analysis, it was demonstrated that protopine retarded the activated NLR family pyrin domain containing 3 (NLRP3) and nuclear factor kappa B (NF-κB) signaling pathways mediated by LPS.
Conclusion: Protopine alleviated LPS-triggered intestinal epithelial cell injury by inhibiting NLRP3 and NF-κB signaling pathways to reduce inflammation and oxidative stress. This discovery may provide a useful drug for treating IBD.
Key words: inflammation, intestinal epithelial cell injury, LPS, oxidative stress, protopine
*Corresponding author: Zhongjun Xu, Department of Medical Imaging, First Affiliated Hospital of Gannan Medical University, No. 128 Jinling Road, Zhanggong District, Ganzhou, Jiangxi, 341000, China. Email address: [email protected]
Received 12 May 2022; Accepted 30 May 2022; Available online 1 November 2022
Copyright: Li J, et al.
License: This open access article is licensed under Creative Commons Attribution 4.0 International (CC BY 4.0). http://creativecommons.org/licenses/by/4.0/
Introduction
Inflammatory bowel disease (IBD) is a common sickness associated with inflammation in the gastrointestinal tract (GIT).1 IBD is a noninfectious, chronic, and relapsed inflammatory disease of GIT, featured by repeated occurrence and persistent diarrhea and abdominal pain, mainly manifested as ulcerative colitis and Crohn’s disease.2–4 The course of these two diseases consists of numerous responses involving environmental, intestinal, and immune factors, and the confusion of immune system homeostasis is a hallmark of IBD.5,6 At present, the treatment of IBD principally consists of anti-inflammatory drugs, immunosuppressants, and antibiotics, but drug resistance remains the main difficulty that limits therapeutic efficacy.7–9 Therefore, there is an urgent requirement to develop novel and useful therapies for IBD.
The study of protopine, which is derived from a variety of plants belonging to poppy (e.g., corydalis), berberis, walnut, and buttercup families,10 has become a hot topic of investigation. Previous reports have demonstrated that protopine has a variety of biological activities and exerts key regulatory functions in varied diseases.11,12 For example, protopine and tetrahydropalmatine modulate the dopamine receptor D2 expression to relieve migraine.13 Protopine suppresses cell apoptosis and inflammation through the toll-like receptor 4 (TLR4) pathway, and improves lipopolysaccharide (LPS)-stimulated acute renal injury.14 In addition, protopine modulates the reactive oxygen species (ROS)/phosphoinositide-3-kinase (PI3K)/protein kinase (Akt) pathway to stimulate apoptosis in liver carcinoma.15 Protopine stabilizes p53 to trigger colon cancer cell apoptosis and autophagy.16 Moreover, protopine inhibits Histone Deacetylase 6 (HDAC6) gene expression in Alzheimer’s disease to accelerate the proteasomal degradation of pathological tau.17 Protopine affects the mitogen-activated protein kinase–nuclear factor kappa B (MAPK/ NF-κB) pathway to reduce inflammation triggered by LPS and carrageenan.18 Interestingly, the total alkaloid fraction from Fumaria capreolata (AFC) (with 1.3% stylopine and 0.9% protopine) demonstrates intestinal anti-inflammatory effects in colitis.19 However, special functions of protopine in the progression of IBD remains unclear.
In this study, we aimed to explore the regulatory effects of protopine on the progression of IBD. Our results demonstrated that protopine modulated LPS-stimulated cell viability, apoptosis, inflammation, and oxidative stress, and inhibited NLR family pyrin domain containing 3 (NLRP3) and NF-κB signaling pathway activity to relieve progression of IBD.
Materials and Methods
Cell lines and culture
NCM460 (a normal human colon mucosal epithelial cell line) cells were acquired from American Tissue Culture Collection (ATCC, Manassas, VA, USA). The Dulbecco’s Modified Eagle Medium (DMEM; Thermo Fisher, Shanghai, China) containing 10% fetal bovine serum (FBS; Gibco, MA, USA) and 1% penicillin/streptomycin was applied to cultivate NCM460 cells at 37°C in a moist incubator with 5% CO2. LPS (from Escherichia coli 0111:B4; Sigma-Aldrich St. Louis, MO, USA; Merck KGaA, Darmstadt, Germany; 0, 2, 5, and 10 μg/mL) or protopine (Sigma-Aldrich; 0, 5, 10, and 20 μM) was used to treat NCM460 cells.
MTT assay
MTT colorimetric assay was carried out following previous methods.20,21 Cells (1000 cells/well) were plated in a 96-well plate, and each well was mixed with 20-μL MTT solution (5 mg/L; Sigma-Aldrich). After 4 h, the generated formazan crystals were dissolved in 150-μL dimethyl sulfoxide (DMSO; Sigma-Aldrich). The cell viability was evaluated at 490 nm under a microplate reader (Multiskan EX; Lab Systems, Helsinki, Finland).
Western blot analysis
Proteins from NCM460 cells were separated with radio-immunoprecipitation assay (RIPA) lysis buffer (Beyotime Institute of Biotechnology, Jiangsu, China), and the concentration was evaluated through bicinchoninic acid assay (BCA) kit (Thermo Fisher Scientific Inc., Waltham, MA, USA).22,23 Protein electrophoresis was done through 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE). The separated proteins were transferred to polyvinylidene fluoride (PVDF) membranes (Beyotime, Shanghai, China). After blocking, the membranes were incubated with the following primary antibodies at 4°C for 12 h: Bax (1:1000; ab32503; Abcam, Shanghai, China), Bcl-2 (1:2000; ab182858), NLRP3 (1:1000; ab263899), p-IκBα (1:10,000; ab133462), IκBα (1:1000; ab32518), p-P65 (1:2000; ab86299), P65 (0.5 µg/mL; ab16502), and β-actin (1:1000; ab8227). Next, the secondary antibodies (1:2000; ab7090) were supplemented into membranes. β-actin acted as an internal reference. Eventually, the blots were assessed using chemiluminescence detection kit (Thermo Fisher Scientific).
Flow cytometry
The cell apoptosis was determined through an Annexin-V-PI apoptosis detection kit (BD Biosciences, Franklin Lakes, NJ, USA).24,25 After rinsing with phosphate-buffered saline (PBS), NCM460 cells were resuspended. Next, NCM460 cells were subjected to Annexin V-fluorescein isothiocyanate (FITC, 5 μL, 50 μg/mL) and propidium iodide (PI, 5 μL, 50 μg/mL) for counterstaining in darkness. Cell apoptosis was examined under flow cytometry (FCM; BD Biosciences).
Enzyme-linked immunosorbent serological assay (ELISA)
The collected cell supernatant was applied to measure the levels of tumor necrosis factor-α (TNF-α; Cat. No. ab181421), Interleukin (IL)-1β (Cat. No. ab214025), and IL-6 (Cat. No. ab178013) in line with the corresponding instructions provided with ELISA kits (Abcam).26
The total contents of superoxide dismutase (SOD), malondialdehyde (MDA), total antioxidant capacity (T-AOC), and myeloperoxidase (MPO) were evaluated with the corresponding commercial ELISA kits (Thermo Fisher Scientific).27
Detection of ROS
The ROS level was assessed by staining NCM460 cells with 2'7'-dichlorodihydrofluorescein diacetate (DCFH-DA; Sigma-Aldrich).28,29 NCM460 cells and DCFH-DA (10 mM) were mixed in darkness for 0.5 h for incubation. Finally, the ROS index was assessed through flow cytometry (BD Bioscience).
Immunofluorescence assay
NCM460 cells were cultured in 5-μM Fluo-3 AM (Biyuntian, China) at a loading temperature (TL) of 37°C. After washing in pure Hanks’ balanced salt solution (HBSS), cells were allowed to de-esterify at a de-esterification temperature (TD) of 37°C. Finally, in order to assess intracellular calcium concentration, images of NCM460 cells with Fluo-3 AM staining were captured under a fluorescence microscope (Olympus, Tokyo, Japan).30
The NCM460 cells were incubated with 4-μg/mL rhodamine 123 (Rh123; KeyGEN Biotech. Co. Ltd, China). The fluorescence intensity of Rh123 in cells was observed to assess the level of mitochondrial membrane potential (MMP).31,32
Statistical analysis
The data were represented as the mean ± standard deviation (SD). The statistical analysis was conducted using the GraphPad Prism software, version 8.0 (GraphPad Software, La Jolla, CA, USA). All experiments were repeated for three times. The Student’s t-test or one-way analysis of variance (ANOVA) was employed for comparison between two or more groups; P < 0.05 was deemed as statistically significant.
Results
Protopine relieved LPS-triggered intestinal epithelial cell injury
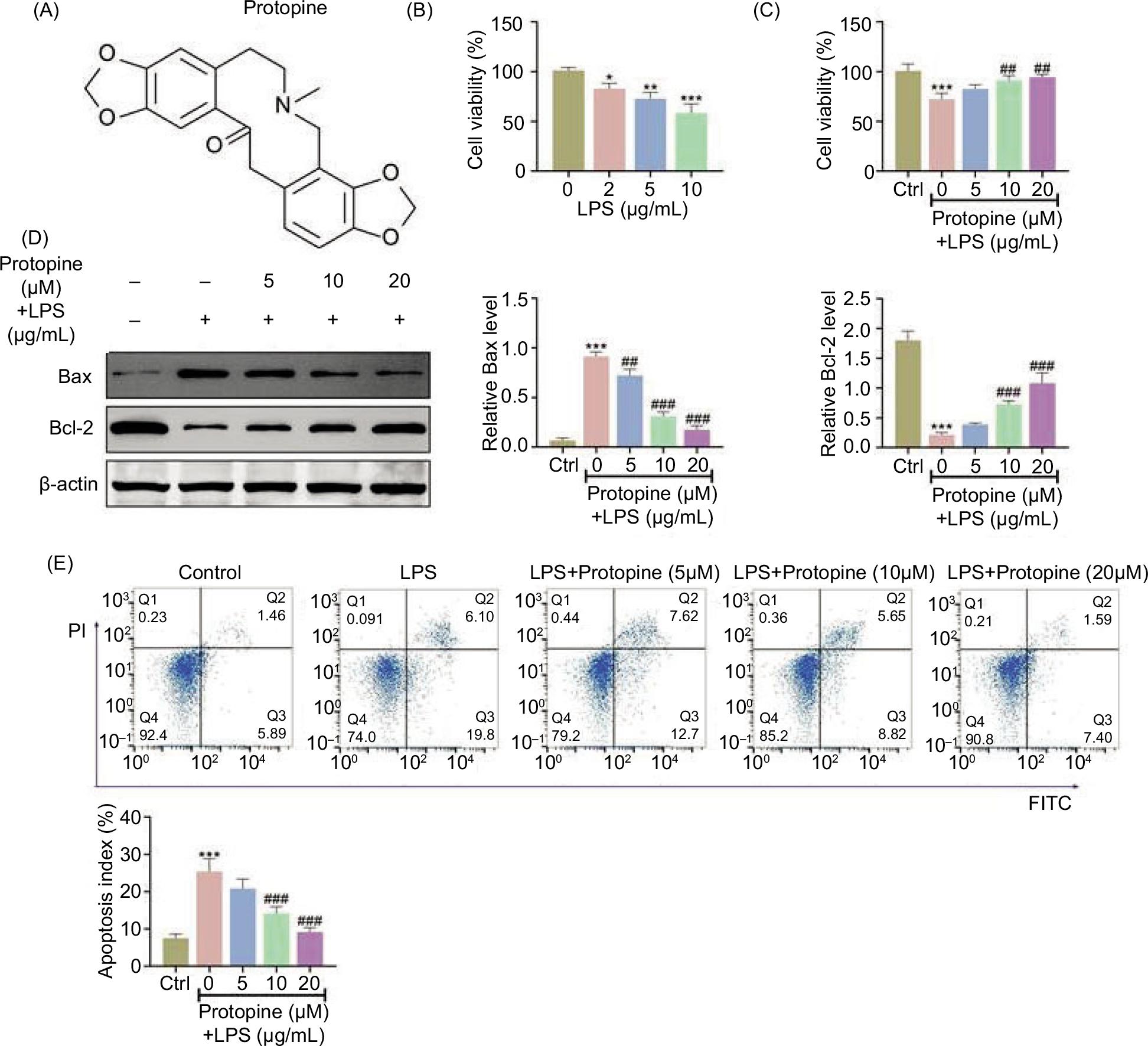
The chemical structure of protopine is displayed in Figure 1A. The cell viability of NCM460 cells was weakened with the increased concentration of LPS (0, 2, 5, and 10 μg/mL; Figure 1B). The cell viability was weakened to about 70% in NCM460 cells treated with 5-μg/mL LPS. Thus, the concentration of 5-μg/mL LPS was chosen for further experiments. Next, it was demonstrated that the weakened cell viability stimulated by LPS was reversed by protopine treatment in a dose-dependent manner (Figure 1C). Specially, 20-μM protopine could restore cell viability to 94%. In addition, cell apoptosis proteins (Bax and Bcl-2) were determined by Western blot analysis. The up-regulated Bax expression and down-regulated Bcl-2 expression triggered by LPS were offset by protopine treatment (Figure 1D). Moreover, the enhanced cell apoptosis index mediated by LPS (7%–25%: Ctrl to 5-μg/mL LPS) and relieved by protopine treatment (25%–8%: 0–20-μM protopine) was detected by flow cytometry (Figure 1E). These data revealed that protopine relieved LPS-triggered intestinal epithelial cell injury.
Figure 1 Protopine relieved LPS-triggered intestinal epithelial cell injury. (A) The chemical structure diagram of protopine. (B) The cell viability was assessed with the increased concentration of LPS (0, 2, 5, and 10 μg/mL) through MTT assay. (C) Groups were divided as follows: control, LPS, LPS+protopine (5 μM), LPS+protopine (10 μM), and LPS+protopine (20 μM) groups. The cell viability was evaluated through MTT assay. (D) Groups were divided as follows: control, LPS, LPS+protopine (5 μM), LPS+protopine (10 μM), and LPS+protopine (20 μM) groups. The protein expressions of Bax and Bcl-2 were examined through Western blot analysis. (E) Groups were divided as follows: control, LPS, LPS+protopine (5 μM), LPS+protopine (10 μM), and LPS+protopine (20 μM) groups. The cell apoptosis was measured through flow cytometry. *P < 0.05, **P < 0.01, ***P < 0.001 vs. the control group; ##P < 0.01, ###P < 0.001 vs, the LPS group.
Protopine reduced LPS-mediated inflammation
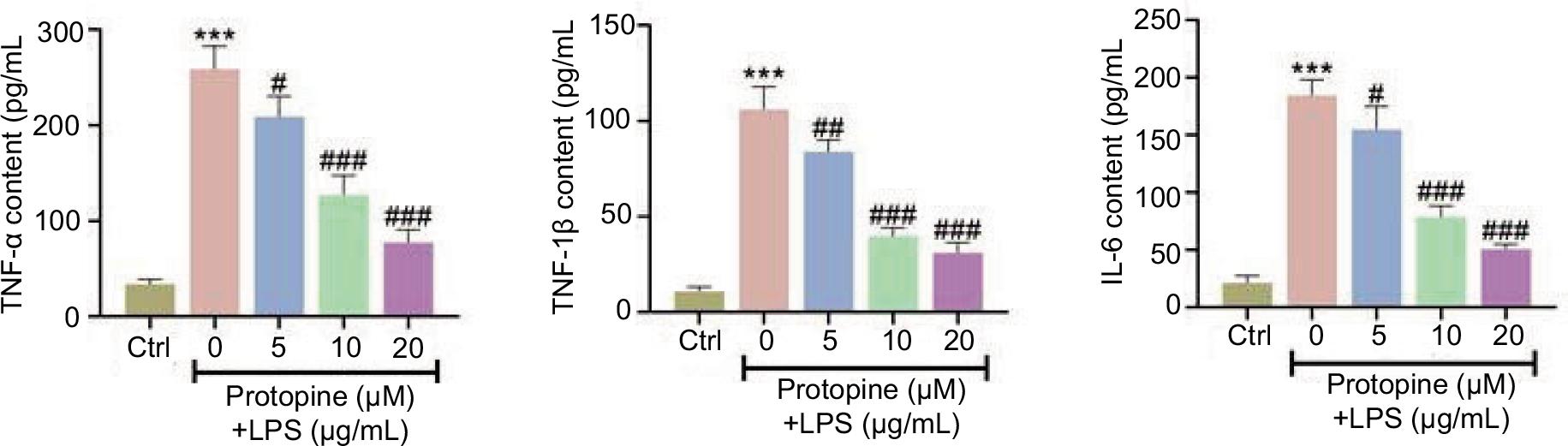
Results from ELISA established that the increased levels of TNF-α, IL-1β, and IL-6 triggered by LPS were attenuated by protopine treatment (TNF-α, 259–77 pg/mL; IL-1β, 106–31 pg/mL; IL-6, 185–51 pg/mL: 0–20-μM protopine) (Figure 2). In general, protopine reduced LPS-mediated inflammation.
Figure 2 Protopine reduced LPS-mediated inflammation. Groups were divided as follows: control, LPS, LPS+protopine (5 μM), LPS+protopine (10 μM), and LPS+protopine (20 μM) groups. The levels of TNF-α, IL-1β, and IL-6 were tested through ELISA. ***P < 0.001 vs. the control group; ##P < 0.01, ###P < 0.001 vs. the LPS group.
Protopine attenuated LPS-induced oxidative stress
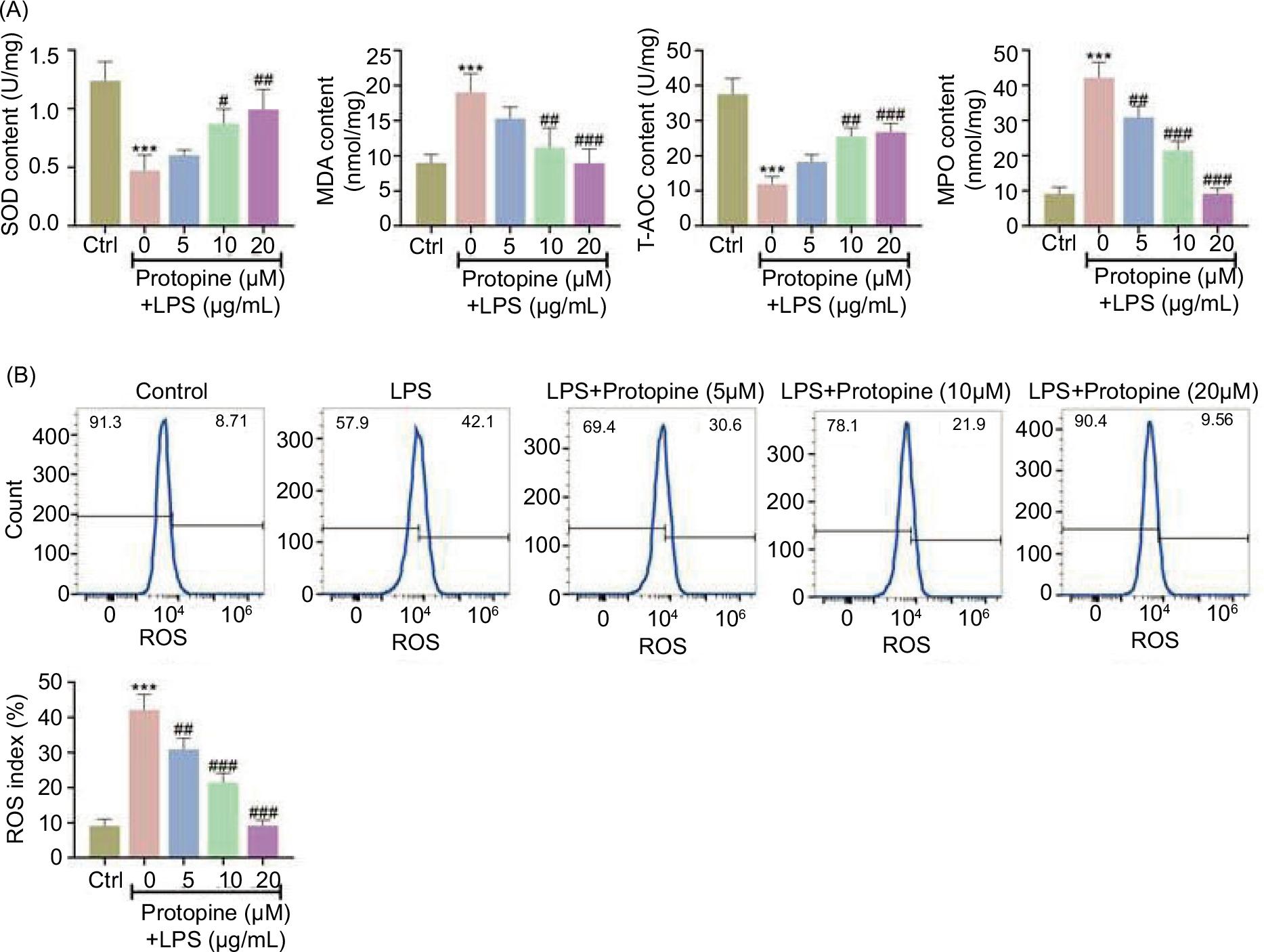
The decreased SOD and T-AOC levels and increased MDA and MPO levels stimulated by LPS were rescued after cells were treated with protopine (Figure 3A). Moreover, the ROS level was strengthened by LPS (9%–42%: Ctrl to 5-μg/mL LPS), but this effect was weakened after treatment with protopine (42%–10%: 0–20-μM protopine; Figure 3B). Taken together, protopine attenuated LPS-induced oxidative stress.
Figure 3 Protopine attenuated oxidative stress. Groups were divided as follows: control, LPS, LPS+protopine (5 μM), LPS+protopine (10 μM), and LPS+protopine (20 μM) groups. (A) The levels of MDA, SOD, T-AOC, and MPO were evaluated through ELISA. (B) The ROS level was assessed through flow cytometry. ***P < 0.001 vs. the control group; ##P < 0.01, ###P < 0.001 vs. the LPS group.
Protopine decreased intracellular Ca2+ concentration and increased mitochondrial membrane potential
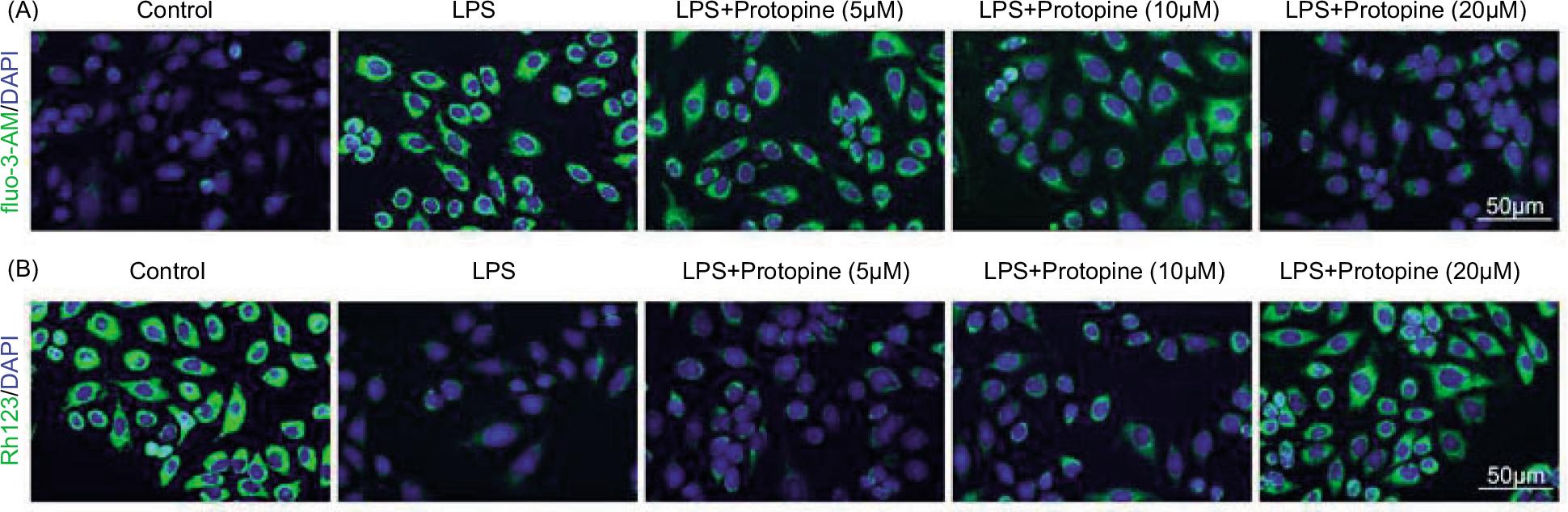
The increased fluo-3-AM fluorescence intensity (intracellular Ca2+ concentration) stimulated by LPS was offset by protopine treatment (Figure 4A). Additionally, the decreased Rh123 fluorescence intensity (mitochondrial membrane potential) mediated by LPS was reversed by protopine treatment (Figure 4B).
Figure 4 Protopine decreased intracellular Ca2+ concentration and increased mitochondrial membrane potential. Groups were divided as follows: control, LPS, LPS+protopine (5 μM), LPS+protopine (10 μM), and LPS+protopine (20 μM) groups. (A and B) The intracellular Ca2+ concentration and mitochondrial membrane potential were measured through immunofluorescence assay.
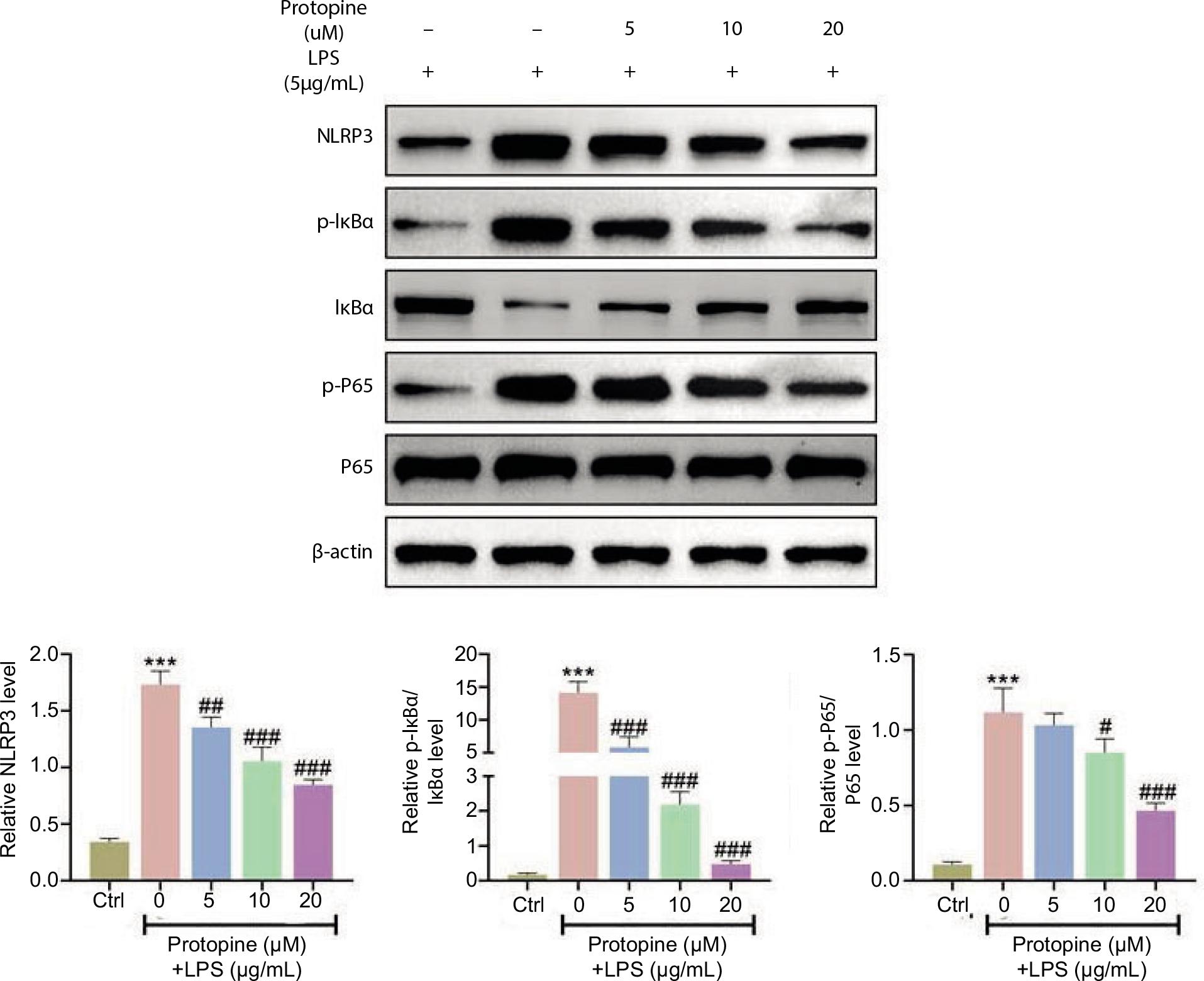
Protopine inhibited NLRP3 and NF-κB signaling pathways
Finally, the effects of protopine on the NLRP3 and NF-κB signaling pathways were investigated. The up-regulated protein expressions of NLRP3, p-IκBα/IκBα, and p-P65/P65 triggered by LPS were inhibited by protopine treatment (NLRP3, 1.7–0.8; p-IκBα/IκBα, 14–0.5; p-P65/P65, 1.1–0.5: 0–20-μM protopine; Figure 5). These findings indicated that protopine inhibited the NLRP3 and NF-κB signaling pathways.
Figure 5 Protopine inhibited the NLRP3 and NF-κB signaling pathways. Groups were divided as follows: control, LPS, LPS+protopine (5 μM), LPS+protopine (10 μM), and LPS+protopine (20 μM) groups. The protein expressions of NLRP3, p-IκBα/IκBα, and p-P65/P65 were determined through Western blot analysis. ***P < 0.001 vs. the control group; ##P < 0.01, ###P < 0.001 vs. the LPS group.
Discussion
Increasing varieties of plant extracts have been demonstrated as involved in the regulation of IBD progression. For example, Ginsenoside Rg1 modulates the balanced differentiation of Tfh/Treg cells to alleviate experimental colitis.33 Additionally, oxymatrine represses the RhoA/ROCK signaling pathway to relieve dextran sulfate sodium (DSS)-triggered acute intestinal inflammation.34 Moreover, pogostone suppresses T helper cells to attenuate 2,4,6-trinitrobenzenesulfonic acid-stimulated experimental colitis.35 In DSS-stimulated acute colitis, alpinetin inhibits the TLR4 and NLRP3 pathways to improve inflammatory responses.36
Protopine, an isoquinoline alkaloid, has asthma relieving, cough relieving, sedation, analgesic, antiplatelet aggregation, and antibacterial functions. It can be used for treating malaria, fundus congestion, and thrombotic disorders. Protopine is often used as anti-tumor agent, smooth muscle relaxant, antibacterial agent, and sedative drug in clinical applications.10,37,38 Protopine has been investigated for its special functions in varied diseases.11,14–18 However, regulatory effects of protopine in the progression of IBD remains unclear. Our work explored the effects of protopine on the progression of IBD and its related regulatory pathways. In previous studies, protopine was found to modulate cell viability and apoptosis to affect the progression of diseases.14–16 Similarly, in this study, we discovered that protopine rescued the inhibited intestinal epithelial cell viability and the enhanced cell apoptosis triggered by LPS. Moreover, protopine also controlled inflammation, oxidative stress, and other cellular progressions.11,18,39 In the present study, the results demonstrated that protopine attenuated LPS-induced inflammation and oxidative stress. Further experiments illustrated that the increased intra-cellular Ca2+ concentration and decreased mitochondrial membrane potential stimulated by LPS were reversed by protopine treatment.
The production of pro-inflammatory cytokines has been under the jurisdiction of NLRP3.40 NLRP3 is an apoptosis-relevant dot-like protein, containing activated procaspase-1 and caspase-1.41,42 Activation of caspase-1 is essential for the conversion of pro-IL-1β into its mature active form IL-1β, which is activated in macrophages by LPS-activated NLRP3 inflammasome.43,44 Upon activation, NLRP3 binds to ASC connector, and triggers the translocation and activation of pro-caspase-1.45 In addition, the NF-κB pathway modulates immune response and cell survival by regulating the expressions of pro-inflammatory and pro-survival genes.46 The NLRP3 and NF-κB signaling pathways have been revealed to participate in progression of IBD. For instance, NEK7 and NLRP3 affect NF-κB signaling to regulate pyroptosis in progression of IBD.47 Monotropein modulates the NLRP3 inflammasome and the TLR4/NF-κB pathway in chronic colitis to relieve secondary liver injury.48 In addition, the fermentation broth of Platycodon grandiflorum root regulates the adenosine monophosphate- activated protein kinase (AMPK)/NF-κB/NLRP3 pathway to attenuate inflammation in IBD.49 Moreover, carboxyamidotriazole affects NLRP3 inflammasome and NF-κB activation to ameliorate 2,4,6-trinitrobenzene sulfonic acid-triggered colitis.50 In the present work, we proved that protopine repressed the activation of the NLRP3 and NF-κB signaling pathways mediated by LPS.
Conclusion
Our study for the first time explored the effects of protopine on progression of IBD and its associated regulatory pathways. Our results revealed that protopine alleviated LPS-triggered intestinal epithelial cell injury by inhibition of the NLRP3 and NF-κB signaling pathways to reduce inflammation and oxidative stress. This findings of the present study established that protopine could be a validated drug for IBD therapeutic strategy. However, this study also has some limitations regarding the impact of protopine on progression of IBD. For instance, the effects of protopine on IBD in animal models, clinical samples as well as other cellular progressions must be investigated in the future studies. In addition, more experiments must be conducted to explore other regulatory functions of protopine in IBD and different diseases.
Availability of Data and Materials
All data generated or analyzed in this study are included in this published article.
Competing Interests
The authors stated that there were no conflicts of interest to disclose.
Author Contributions
Junyu Li and Zhongjun Xu designed the study, completed the experiments, and supervised data collection. Canhui OuYang analyzed and interpreted the data. Xiongjian Wu, Yun Xie, and Jun Xie prepared the manuscript for publication and reviewed final draft of the manuscript. All authors read and approved the final manuscript.
REFERENCES
1. Sairenji T, Collins KL, Evans DV. An update on inflammatory bowel disease. Prim Care. 2017;44(4):673–92. 10.1016/j.pop.2017.07.010
2. Seyedian SS, Nokhostin F, Malamir MD. A review of the diagnosis, prevention, and treatment methods of inflammatory bowel disease. J Med Life. 2019;12(2):113–22. 10.25122/jml-2018-0075
3. Zhang YZ, Li YY. Inflammatory bowel disease: Pathogenesis. World J Gastroenterol. 2014;20(1):91–9. 10.3748/wjg.v20.i1.91
4. Kennedy B, Sexton K, Gleeson N. Outcomes of palliative surgery for bowel obstruction in end of life ovarian cancer care in a tertiary cancer centre. Eur J Gynaecol Oncol. 2021;42(6):1112–5. 10.31083/j.ejgo4206162
5. Flynn S, Eisenstein S. Inflammatory bowel disease presentation and diagnosis. Surg Clin N Am. 2019;99(6):1051–62. 10.1016/j.suc.2019.08.001
6. Davis BK, Philipson C, Hontecillas R, Eden K, BassaganyaRiera J, Allen IC. Emerging significance of NLRs in inflammatory bowel disease. Inflamm Bowel Dis. 2014;20(12):2412–32. 10.1097/MIB.0000000000000151
7. Nitzan O, Elias M, Peretz A, Saliba W. Role of antibiotics for treatment of inflammatory bowel disease. World J Gastroenterol. 2016;22(3):1078–87. 10.3748/wjg.v22.i3.1078
8. Wehkamp J, Götz M, Herrlinger K, Steurer W, Stange EF. Inflammatory bowel disease. Dtsch Arzteblatt Int. 2016;113(5):72–82. 10.3238/arztebl.2016.0072
9. Wallam S, Dugan K, Levinson K. Salvage chemotherapy after treatment with immune checkpoint inhibitors in women with recurrent cervical cancer. Eur J Gynaecol Oncol. 2021;42(2):394.
10. Huang W, Kong L, Cao Y, Yan L. Identification and quantification, metabolism and pharmacokinetics, pharmacological activities, and botanical preparations of protopine: A review. Molecules (Basel, Switzerland). 2021;27(1):215. 10.3390/molecules27010215
11. Bae DS, Kim YH, Pan CH, Nho CW, Samdan J, Yansan J, et al. Protopine reduces the inflammatory activity of lipopolysaccharide-stimulated murine macrophages. BMB Rep. 2012;45(2):108–13. 10.5483/BMBRep.2012.45.2.108
12. Kosina P, Gregorova J, Gruz J, Vacek J, Kolar M, Vogel M, et al. Phytochemical and antimicrobial characterization of Macleaya cordata herb. Fitoterapia. 2010;81(8):1006–12. 10.1016/j.fitote.2010.06.020
13. Zhang X, Wang Y, Zhang K, Sheng H, Wu Y, Wu H, et al. Discovery of tetrahydropalmatine and protopine regulate the expression of dopamine receptor D2 to alleviate migraine from Yuanhu Zhitong formula. Phytomed Int J Phytother Phytopharmacol. 2021;91:153702. 10.1016/j.phymed.2021.153702
14. Zhang B, Zeng M, Li M, Kan Y, Li B, Xu R, et al. Protopine protects mice against LPS-induced acute kidney injury by inhibiting apoptosis and inflammation via the TLR4 signaling pathway. Molecules (Basel, Switzerland). 2019;25(1):15. 10.3390/molecules25010015
15. Nie C, Wang B, Wang B, Lv N, Yu R, Zhang E. Protopine triggers apoptosis via the intrinsic pathway and regulation of ROS/PI3K/Akt signalling pathway in liver carcinoma. Cancer Cell Int. 2021;21(1):396. 10.1186/s12935-021-02105-5%
16. Son Y, An Y, Jung J, Shin S, Park I, Gwak J, et al. Protopine isolated from Nandina domestica induces apoptosis and autophagy in colon cancer cells by stabilizing p53. Phytother Res (PTR). 2019;33(6):1689–96. 10.1002/ptr.6357
17. Sreenivasmurthy SG, Iyaswamy A, Krishnamoorthi S, Senapati S, Malampati S, Zhu Z, et al. Protopine promotes the proteasomal degradation of pathological tau in Alzheimer’s disease models via HDAC6 inhibition. Phytomed Int J Phytother Phytopharmacol. 2022;96:153887. 10.1016/j.phymed.2021.153887
18. Alam MB, Ju MK, Kwon YG, Lee SH. Protopine attenuates inflammation stimulated by carrageenan and LPS via the MAPK/NF-κB pathway. Food Chem Toxicol. 2019;131:110583. 10.1016/j.fct.2019.110583
19. Bribi N, Algieri F, Rodriguez-Nogales A, Vezza T, GarridoMesa J, Utrilla MP, et al. Intestinal anti-inflammatory effects of total alkaloid extract from Fumaria capreolata in the DNBS model of mice colitis and intestinal epithelial CMT93 cells. Phytomed Int J Phytother Phytopharmacol. 2016;23(9):901–13. 10.1016/j.phymed.2016.05.003
20. Li YQ, Wang LC, Li AX, Huang W, Song Y, Wang W. LINC00958/ miR-627 signal axis regulates the proliferation, migration, and invasion of thyroid papillary carcinoma cells by TRIM44. Kaohsiung J Med Sci. 2022;38(5):415–24. 10.1002/kjm2.12502
21. Liang M, Zhu B, Wang M, Jin J. Knockdown of long non-coding RNA DDX11-AS1 inhibits the proliferation, migration and paclitaxel resistance of breast cancer cells by upregulating microRNA-497 expression. Mol Med Rep. 2022;25(4):123. 10.3892/mmr.2022.12639
22. Xu X, Xie Q, Xie M, Zeng Y, Liu Q. LncRNA SNHG8 serves as an oncogene in breast cancer through miR-634/ZBTB20 Axis. Cancer Manag Res. 2021;13:3017–28. 10.2147/CMAR.S270128
23. Xu J, Chen Z, Fang Z, Chen S, Guo Y, Liu X, et al. Long non-coding RNA OIP5-AS1 promotes the progression of esophageal cancer by regulating miR-30a/VOPP1 expression. Oncol Lett. 2021;22(3):651. 10.3892/ol.2021.12912
24. Liu XL, Meng YH, Wang JL, Yang BB, Zhang F, Tang SJ. FOXL2 suppresses proliferation, invasion and promotes apoptosis of cervical cancer cells. Int J Clin Exp Pathol. 2014;7(4):1534–43.
25. Zuo X, Li W, Yan X, Ma T, Ren Y, Hua M, et al. Long non-coding RNA LINC01224 promotes cell proliferation and inhibits apoptosis by regulating AKT3 expression via targeting miR-485-5p in endometrial carcinoma. Oncol Rep. 2021;46(3):186. 10.3892/or.2021.8137
26. Ye Z, Wei L, Yin X, Li H, Qin G, Li S, et al. Long non-coding RNA cancer susceptibility candidate 2 regulates the function of human fibroblast-like synoviocytes via the microRNA-18a-5p/B-cell translocation gene 3 signaling axis in rheumatoid arthritis. Bioengineered. 2022;13(2):3240–50. 10.1080/21655979.2021.2022075
27. Zhang J, Jiang H, Liu DH, Wang GN. Effects of dexmedetomidine on myocardial ischemia-reperfusion injury through PI3K-Akt-mTOR signaling pathway. Eur Rev Med Pharmacol Sci. 2019;23(15):6736–43.
28. Wu ZC, Hui XG, Huo L, Sun DX, Peng W, Zhang Y, et al. Antiproliferative effects of isoalantolactone in human liver cancer cells are mediated through caspase-dependent apoptosis, ROS generation, suppression of cell migration and invasion and targeting Ras/Raf/MEK signalling pathway. Acta Biochim Pol. 2022;69(2):299–304. 10.18388/abp.2020_5704
29. Skonieczna M, Hudy D, Poterala-Hejmo A, Hejmo T, Buldak RJ, Dziedzic A. Effects of resveratrol, berberine and their combinations on reactive oxygen species, survival and apoptosis in human squamous carcinoma (SCC-25) cells. Anti-Cancer Agents Med Chem. 2019;19(9):1161–71. 10.2174/1871520619666190405111151
30. Wang L, Li M, Sha B, Hu X, Sun Y, Zhu M, et al. Inhibition of deubiquitination by PR-619 induces apoptosis and autophagy via ubi-protein aggregation-activated ER stress in oesophageal squamous cell carcinoma. Cell Prolif. 2021;54(1):e12919. 10.1111/cpr.12919
31. Jouan E, Le Vée M, Mayati A, Denizot C, Parmentier Y, Fardel O. Evaluation of P-glycoprotein inhibitory potential using a rhodamine 123 accumulation assay. Pharmaceutics. 2016;8(2):12. 10.3390/pharmaceutics8020012.
32. Guo CC, Jiao CH, Gao ZM. Silencing of LncRNA BDNF-AS attenuates Aβ(25-35)-induced neurotoxicity in PC12 cells by suppressing cell apoptosis and oxidative stress. Neurol Res. 2018;40(9):795–804. 10.1080/01616412.2018.1480921
33. Jin J, Zhong Y, Long J, Wu T, Jiang Q, Wang H, et al. Ginsenoside Rg1 relieves experimental colitis by regulating balanced differentiation of Tfh/Treg cells. Int Immunopharmacol. 2021;100:108133. 10.1016/j.intimp.2021.108133
34. Wang Y, Shou Z, Fan H, Xu M, Chen Q, Tang Q, et al. Protective effects of oxymatrine against DSS-induced acute intestinal inflammation in mice via blocking the RhoA/ROCK signaling pathway. Biosci Rep. 2019;39(7):BSR20182297. 10.1042/BSR20182297
35. Su J, Li C, Yu X, Yang G, Deng J, Su Z, et al. Protective effect of pogostone on 2,4,6-trinitrobenzenesulfonic acid-induced experimental colitis via inhibition of T helper cell. Front Pharmacol. 2017;8:829. 10.3389/fphar.2017.00829
36. He X, Wei Z, Wang J, Kou J, Liu W, Fu Y, et al. Alpinetin attenuates inflammatory responses by suppressing TLR4 and NLRP3 signaling pathways in DSS-induced acute colitis. Sci Rep. 2016;6:28370. 10.1038/srep28370
37. Chen CH, Liao CH, Chang YL, Guh JH, Pan SL, Teng CM. Protopine, a novel microtubule-stabilizing agent, causes mitotic arrest and apoptotic cell death in human hormone-refractory prostate cancer cell lines. Cancer Lett. 2012; 315(1):1–11. 10.1016/j.canlet.2015.06.019
38. Saeed SA, Gilani AH, Majoo RU, Shah BH. Anti-thrombotic and anti-inflammatory activities of protopine. Pharmacol Res. 1997;36(1):1–7. 10.1006/phrs.1997.0195
39. Xiao X, Liu J, Hu J, Zhu X, Yang H, Wang C, et al. Protective effects of protopine on hydrogen peroxide-induced oxidative injury of PC12 cells via Ca(2+) antagonism and antioxidant mechanisms. Eur J Pharmacol. 2008;591(1–3):21–7. 10.1016/j.ejphar.2008.06.045
40. Jang SE, Jeong JJ, Hyam SR, Han MJ, Kim DH. Ursolic acid isolated from the seed of Cornus officinalis ameliorates colitis in mice by inhibiting the binding of lipopolysaccharide to toll-like receptor 4 on macrophages. J Agric Food Chem. 2014;62(40):9711–21. 10.1021/jf501487v
41. Faustin B, Lartigue L, Bruey JM, Luciano F, Sergienko E, Bailly-Maitre B, et al. Reconstituted NALP1 inflammasome reveals two-step mechanism of Caspase-1 activation. Mol Cell. 2007;25(5):713–24. 10.1016/j.molcel.2007.01.032
42. Zhang S, Shao Q, Jia L, Zhou F. ANXA3 regulates HIF1 alpha-Induced NLRP3 inflammasome activity and promotes LPS-induced inflammatory response in bronchial epithelial cells. Signa Vitae. 2021;17(3):206–13.
43. Lamkanfi M, Dixit VM. Mechanisms and functions of inflammasomes. Cell. 2014;157(5):1013–22. 10.1016/j.cell.2014.04.007
44. Cho Y-D, Choi S-H, Park S-J, Yu W-S, Cho H-J, Kim K-H, et al. The effects of oxygen and medicines on T cells in hypoxic co-culture. Signa Vitae. 2021;17(6):43–51.
45. Chao LK, Lin CH, Chiu HW, Wong WT, Chiu HW, Tasi YL, et al. Peroxyauraptenol inhibits inflammation and NLRP3 inflammasome activation by inhibiting reactive oxygen species generation and preserving mitochondrial integrity. J Agric Food Chem. 2015;63(4):1210–9. 10.1021/jf5054436
46. Zhang Q, Lenardo MJ, Baltimore D. 30 Years of NF-κB: A blossoming of relevance to human pathobiology. Cell. 2017;168(1–2):37–57. 10.1016/j.cell.2016.12.012
47. Chen X, Liu G, Yuan Y, Wu G, Wang S, Yuan L. NEK7 interacts with NLRP3 to modulate the pyroptosis in inflammatory bowel disease via NF-κB signaling. Cell Death Dis. 2019;10(12):906. 10.1038/s41419-019-2157-1
48. Chen Y, Lu Y, Pei C, Liang J, Ding P, Chen S, et al. Monotropein alleviates secondary liver injury in chronic colitis by regulating TLR4/NF-κB signaling and NLRP3 inflammasome. Eur J Pharmacol. 2020;883:173358. 10.1016/j.ejphar.2020.173358
49. Wang Z, Li C, He X, Xu K, Xue Z, Wang T, et al. Platycodon grandiflorum root fermentation broth reduces inflammation in a mouse IBD model through the AMPK/NF-κB/NLRP3 pathway. Food Funct. 2022;13(7):3946–56. 10.1039/D1FO03969E
50. Du X, Chen W, Wang Y, Chen C, Guo L, Ju R, et al. Therapeutic efficacy of carboxyamidotriazole on 2,4,6-trinitrobenzene sulfonic acid-induced colitis model is associated with the inhibition of NLRP3 inflammasome and NF-κB activation. Int Immunopharmacol. 2017;45:16–25. 10.1016/j.intimp.2017.01.015