
Download
REVIEW ARTICLE
HLA-G in asthma and its potential as an effective therapeutic agent
Jinyan Li, Yuqiu Hao, Wei Li, Xuejiao Lv, Peng Gao*
Jilin University Second Hospital, Changchun City, Jilin Province, China
Abstract
Objective: Asthma is a heterogeneous disease. Severity of asthma and sensitivity to medications vary across asthma subtypes. Human leukocyte antigen (HLA)-G has a wide range of functions in normal and pathological physiology. Due to its powerful immune function, HLA-G participates in the pathogenesis of different asthma phenotypes by regulating the activity and function of various immune cells. The mechanism of HLA-G in asthma is not fully clear, and there is no consensus on its mechanism in asthma. Further studies are needed to explore the role of HLA-G in different phenotypes of human asthma.
Methods: Observational study.
Results: HLA-G is an important immunomodulatory factor in asthma. Studies have found different levels of HLA-G in patients with different asthma subtypes and healthy controls, but other studies have come to the opposite conclusion.
Conclusion: We speculate that further study on the mechanism of HLA-G in asthma pheno-types may explain some of the contradictions in current studies. Findings should provide information regarding the potential of HLA-G as a novel target for asthma diagnosis and treatment.
Key words: HLA-G, Asthma, Asthma Subtypes, Inflammation
*Corresponding author: Peng Gao, No.218 Zijiang Street, Nanguan District, Changchun City, 130000, Jilin Province, China. Email address: [email protected]
Received 13 April 2022; Accepted 6 July 2022; Available online 1 January 2023
Copyright: Li J, et al.
License: This open access article is licensed under Creative Commons Attribution 4.0 International (CC BY 4.0). http://creativecommons.org/licenses/by/4.0/
Introduction
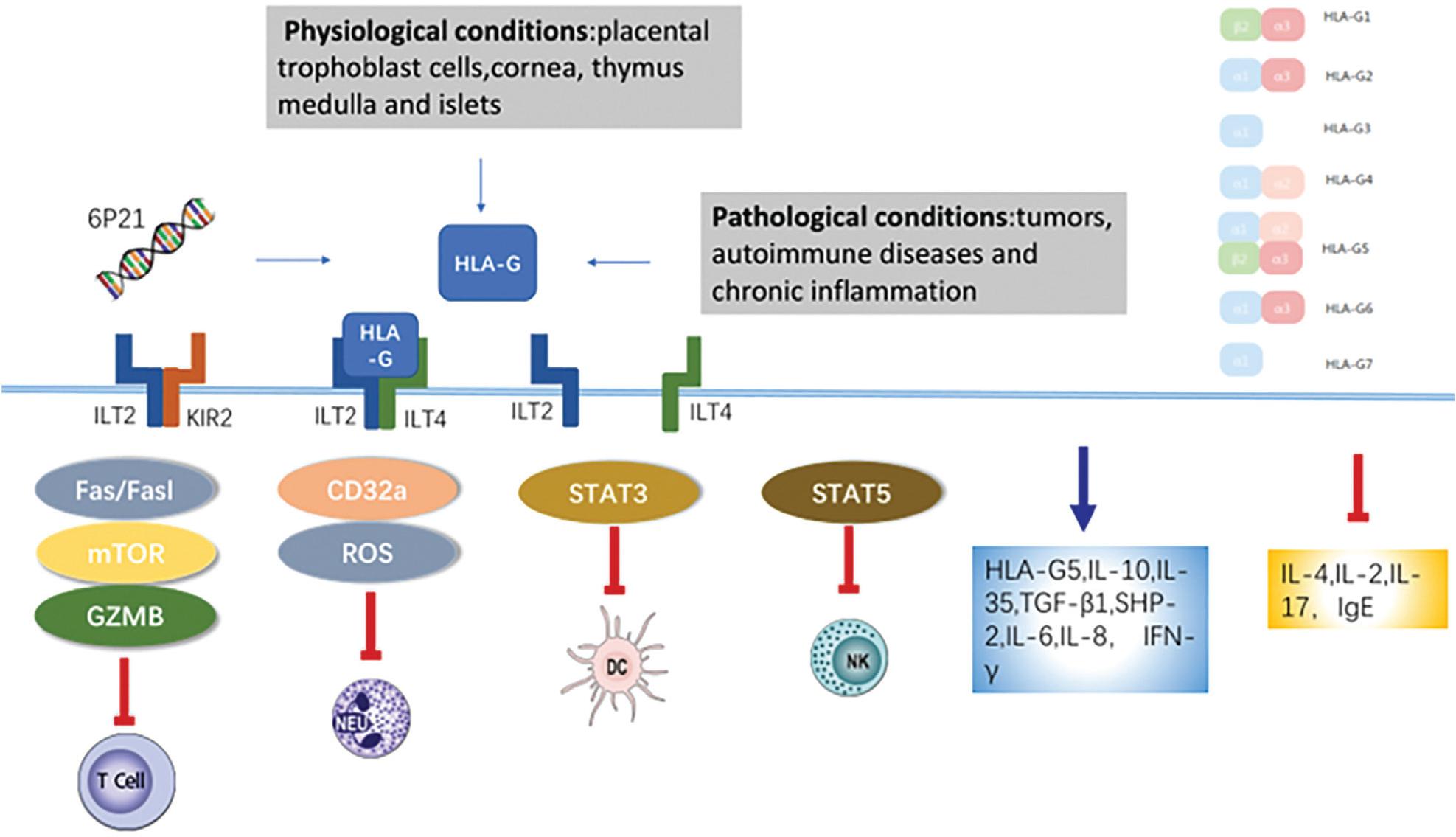
Human leukocyte antigen G (HLA-G) is a non-classical HLA class I molecule that is encoded by a gene on chromosome 6p21.1 Compared to classical HLA-I proteins, HLA-G has limited polymorphism in the coding region and selective tissue distribution, which are driven by an upstream enhancer site.2 HLA-G was originally discovered on fetal extravillous trophoblasts, where it plays a key role in fetus-induced immune tolerance.3 More recently, HLA-G has been implicated in the pathogenesis in pregnancy, organ transplantation, autoimmune diseases, tumors, and inflammatory diseases.4–6 Under physiological conditions, HLA-G mRNA is expressed in a variety of tissues, but HLA-G protein expression is restricted to specific cell types, including placental trophoblast cells, and the cornea, thymic medulla, and pancreatic islets.7 HLA-G expression is strictly controlled at the transcriptional and posttranscriptional level, but may be increased in response to tumors, autoimmune diseases, and chronic inflammation. The HLA-G gene generates different isoforms thorough alternative splicing, including four membrane-bound isoforms (HLA-G1, -G2, -G3, and -G4) and three soluble isoforms (HLA-G5, -G6, and -G7).8 The membrane-bound isoforms correspond to the soluble iso-forms, for example HLA-G5 constitutes the soluble form of HLA-G1. HLA-G1 and HLA-G5 have been extensively studied. They consist of a heavy chain containing three extra globular domains, can form non-covalent associations with β2-microglobulin, and may reduce the cytotoxic activation of natural killer (NK) cells.9 The ratio of HLA-G1/G5 can change under different pathophysiological conditions. Under normal physiological conditions, in human bronchial epithelial cells (HBEC), the transcriptional level of HLAG1 is lower than HLA-G5. However, in lung cancer tissue, the transcriptional level of HLA-G1 may be higher than HLA-G5.10 HLA-G2, -G3, and -G4 are formed by alternative splicing of the full-length transcript HLA-G1. HLA-G2 contains α1 and α3 extracellular domains, HLA-G3 contains an α1 extracellular domain, and HLA-G4 contains α1 and α2 extracellular domains (Figure 1).11
Figure 1 HLA-G binding and signal transduction showing the different isoforms of HLA-G and the effect of HLA-G on different cytokines. The blue arrow indicates positive effects, while the red T-shaped lines indicate negative effects.
Accumulating evidence shows that HLA-G has a wide range of functions in normal and pathological physiology. A membrane-bound form of HLA-G is expressed at the maternal–fetal interface during pregnancy, where it has an important tolerance-inducing function12; however, HLA-G gene polymorphisms are related to habitual abortion and preeclampsia.13,14 HLA-G expression levels are high in tumors, providing a microenvironment for tumor cells to evade immune surveillance.15
Asthma can be defined as a syndrome with multiple phenotypes, which are important for the prediction of treatment response and long-term prognosis, and for the development of individualized treatment regimens.16 Airway inflammation in patients with asthma is classified into four subtypes based on the ratio of eosinophils to neutrophils in induced sputum: neutrophilic asthma (NA), eosinophilic asthma (EA), paucigranulocytic asthma (PA), and mixed granulocytic asthma (MA). NA is mostly seen in symptomatic adult patients and is associated with exposure to stimuli such as ozone, particulate matter, cigarette smoke, occupational irritants, microbial infections and colonization, and aeroallergens.17,18 NA is common in obese women. In males, serum saturated fatty acid levels are strongly associated with serum neutrophil levels.19 Studies have shown that NA is not sensitive to inhaled glucocorticoid therapy;17,20 patients with refractory asthma have increased airway neutrophils, and neutrophilic inflammation is associated with progressive decline in lung function and persistent airflow limitation.21,22 Patients with EA have high fractional exhaled nitric oxide (FENO) levels and atopic symptoms. Most respond well to treatment with inhaled corticosteroids, and the presence or absence of eosinophils in sputum is an important indicator for assessing prognosis.23,24 PA is the most common phenotype in patients with stable asthma and is associated with better and less variable lung function, lower airway reactivity, a lower incidence of severe refractory asthma, a lower need for inhaled high-dose glucocorticoids, and a better response to glucocorticoid therapy than other phenotypes.25,26 Patients with MA have the worst lung function, the most airflow limitation, and more severe symptoms; this type of asthma has a high incidence of severe refractory asthma, clinical disease burden, and use of health-care resources.26,27 Different phenotypes of asthma have different pathophysiological mechanisms and clinical symptoms and respond differently to treatment. Further elucidation of the pathogenesis of asthma based on the classification of inflammatory cells in sputum may identify new therapeutic targets, improve disease control, and facilitate personalized care.
HLA-G is involved in the pathogenesis of many chronic inflammatory and allergic diseases, including, but not limited to, systemic lupus erythematosus (SLE), multiple sclerosis (MS), Crohn’s disease, ulcerative colitis, allergic rhinitis (AR), and asthma.28–34 Current opinion suggests that HLA-G may have an immunomodulatory role in airway allergic disease and suppress allergic inflammation; however, the underlying mechanisms remain to be elucidated.35–37 AR is a chronic inflammation of the mucosa mediated by T-helper 2 (Th2) cells. sHLA-G is significantly increased in patients with AR compared to healthy controls, and significantly correlated with immunoglobulin E (IgE) levels. sHLA-G levels in plasma and bronchoalveolar lavage fluid (BALF) are increased in patients with asthma.38 Tahan et al. found that plasma sHLA-G levels were not significantly different between children with asthma and healthy controls, and there were no significant correlations of plasma sHLA-G levels to eosinophil counts or total IgE levels.39 However, Ciprandi et al. showed that serum sHLA-G levels were significantly higher in patients with AR or allergic asthma compared to nonallergic patients or healthy controls, and serum sHLA-G levels were related to serum allergen–specific IgE levels.38 We speculate that the conflicting results may come from sample differences, not further distinguishing asthma endotypes in the study, and that HLA-G may have different expression levels in different asthma subtypes. Further study of sHLA-G levels in patients with different asthma subtypes will help us to further explore the molecular mechanism of HLA-G on asthma subtypes.
We searched PubMed for papers published between 2000 and 2020 by including the key word “HLA-G.” Relevant papers were included after manual selection based on the abstracts and discussion within the research group. The present review summarizes the published literature on the functions and mechanisms of HLA-G in inflammatory cells. Humans and experimental models explore its role in the pathophysiology of asthma, and discuss the potential of HLA-G-based therapeutic strategies against asthma.
HLA-G and inflammation
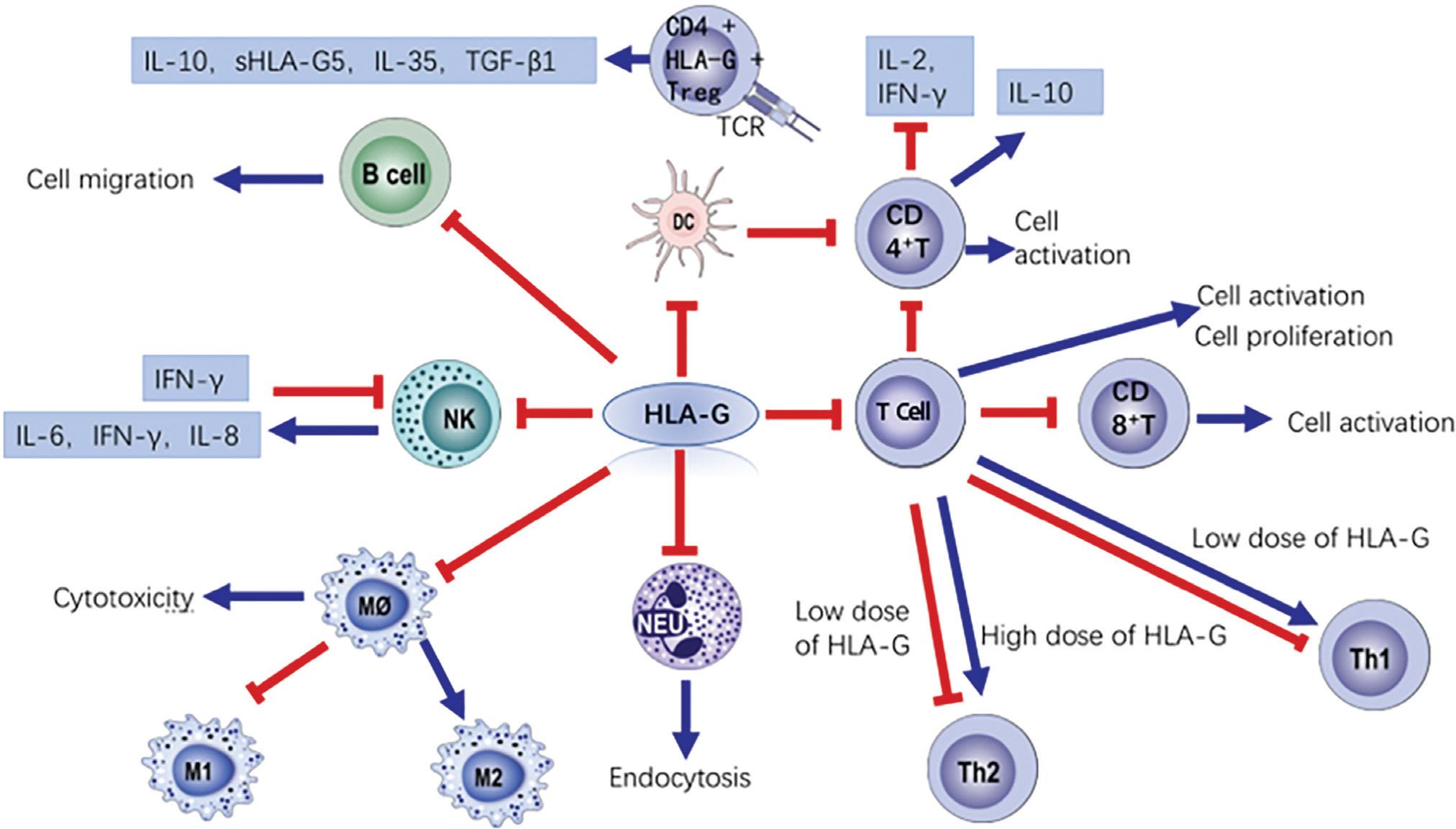
In general, the effects of HLA-G on the immune system include suppression of innate immune responses and activation of adaptive immune responses. HLA-G inhibits the immune response of NK cells, dendritic cells (DC), B cells, T cells, and monocytes or macrophages (Figure 2) by engaging with cell-surface inhibitory receptors,40–42 such as immunoglobulin-like transcript 2(ILT-2 [LILRB1/CD85j]),11,43 immunoglobulin-like transcript 4(ILT-4 [LILRB2/CD85d]) and killer cell immunoglobulin-like receptor, two Ig domains and long cytoplasmic tail 4(KIR2DL4 [CD158d]).44 The HLA-G heavy chain specifically binds to ILT4 and KIR2DL4 receptors, and β2-microglobulin specifically binds to ILT2 receptors. NK cells express ILT2 and KIR2DL4 receptors, DC express ILT4 receptors, B cells express ILT2 receptors, CD4+T cells express ILT2 receptors, and CD8+T cells express ILT2 and KIR2DL4 receptors.45 The ILT2, ILT4/HLA-G pathway has a powerful inhibitory effect on cytotoxicity, which has the potential as a target for antitumor therapy.46
Figure 2 HLA-G effects on inflammation. The blue arrow indicates positive effects, while the red T-shaped lines indicate negative effects.
T cells
HLA-G inhibits T-cell proliferation by activating the phosphatase Src homology-2 domain containing protein tyrosine phosphatase 2 (SHP-2) pathway and inhibiting the mammalian target of rapamycin (mTOR) pathway,47,48 inhibits T cell activity through the FAS/FASL pathway to exert immunosuppressive effects,49 and inhibits CD8+T activation through the GZMB (Granzyme B) pathway.50 Regulatory HLA-G+ CD4+ tTreg (regulatory T cells) are a tTreg subset that exert immunosuppressive effects and secrete high levels of sHLA-G5 and interleukin (IL) 10 and modest levels of IL-35 and transforming growth factor (TGF)-β1.51
A study in patients with tuberculosis showed that HLA-G inhibited the differentiation of Th1 cells, while administration of ILT-4 receptor blockers significantly increased the production of T-helper 1 (Th1) cytokines.52 Some studies have also found that the expression of chemokine cxcr3 decreases after HLA-G stimulation of CD4+T cells, which inhibits the function of Th1.53 Interestingly, HLA-G has a concentration-dependent effect on T-cell polarization. High concentrations of HLA-G induce the polarization of T cells toward the Th2 profile, which promotes the activation of Treg cells and immune tolerance, a mechanism that is involved in Th2-high asthma.54 At low concentrations, HLA- G induces the polarization of T cells toward the Th1 profile.55 A study in women with preeclampsia revealed low levels of HLA-G in peripheral blood mononuclear cells (PBMC), a low frequency of regulatory CD8+CD28- T cells, high levels of IL-17, IL-2, tumor necrosis factor α (TNF-α), IL-4, and IL-10, and an increased proliferative cell activation profile.56 Morandi et al. analyzed the presence or absence of sHLA-G-stimulated CD4+ T cells by flow cytometry and found that sHLA-G impairs the physiological function of Tfh cells by inhibiting the expression level of the chemokine CXCR5.53
Dendritic cells
HLA-G may inhibit the activity of DC through the IL-6-STAT3 pathway,57 while HLA-G modified–DC significantly decrease the activation of CD4+CD25+T cells and the levels of IL-2 and interferon-γ (IFN-γ) produced by CD4+ T cells. An analysis of T-cell subtypes showed that modified DC significantly increased the number of CD4+ T cells capable of producing IL-10. Exposure of LPS-treated DC from ILT-4 transgenic mice to HLA-G regulated CD4+ T-cell activation by decreasing the presentation of peptides by MHC class II molecules.58
B cells
HLA-G inhibits the differentiation and maturation of B cells and inhibits the immune response by promoting the secretion of anti-inflammatory factors.59 The expression of HLA-G in B cells is positively correlated to IFN-γ levels, and the expression of ILT4 is negatively correlated to TNF-α levels.60
Natural killer cells
NK cells produce IFN-γ. HLA-G binds to the KIR2DL4 receptor and inhibits the activity of NK cells; the inhibitory effect of HLA-G5 is greater than that of HLA-G1.6,61,62 HLA-G can also inhibit NK cytotoxicity by interacting with ILT-2 and affecting the NK cell cytoskeleton.63 Specifically, sHLA-G inhibits NK cytotoxicity by downregulating STAT5 phosphorylation.64 The interaction of HLA-G and NK cells can increase the expression of IL-6 and IL-8.65
Neutrophils and macrophages
HLA-G inhibits the phagocytotic function of neutrophils through the ILT4 receptor and inhibits CD32a-mediated reactive oxygen species (ROS) production.63 A study in wild type C57Bl/6 mice that underwent partial hepatectomy showed that knockdown of H2-Bl, a key immunotolerance protein and homolog of human HLA-G, was associated with increased infiltration or activation of macrophages in the liver.66 In human PBMC treated with granulocyte macrophage colony–stimulating factor, the presence of sHLA-G5 promoted the differentiation of macrophages toward an M2 phenotype, inhibited M1 polarization, and promoted the secretion of IL-6 and C-X-C motif ligand 1.67 Porcine endothelial cells transfected with HLA-G1 significantly inhibited the cytotoxicity of macrophages.68
The role of HLA-G in asthma models
Studies using microarray gene probe technology to detect HLA-G expression in human and mouse embryos of different gestational ages demonstrated that HLA-G is involved in the development of normal lung tissue.69 Researches on the role of HLA-G using animal models of asthma are scarce. These studies are required to verify the function of HLA-G in different asthma subtypes, and further understanding of the molecular mechanisms of HLA-G in asthma.
The role of HLA-G in human asthma
Asthma is a heterogeneous disease, characterized by bronchial epithelial hyperplasia, chronic airway inflammation, and airway remodeling.70–72 A variety of inflammatory cells and cytokines are involved in the pathogenesis of asthma, including T lymphocytes, eosinophils, macrophages, neutrophils, IL-10, and IFN-γ.73 According to the type and proportion of cells in an induced sputum smear, asthma may be categorized as EA, NA, MA, or PA.74–76 Severity of asthma and sensitivity to medications vary across asthma sub-types. Eosinophilic asthma is characterized by high levels of systemic eosinophils, a Th2 phenotype, and sensitivity to corticosteroid treatment. Neutrophilic asthma is characterized by activation of the innate immune system (including toll-like receptors [TLRs] and the NOD-like receptor protein 3 [NLRP3] inflammasome) and is steroid-resistant.77
It is essential to clarify the pathophysiological and molecular mechanisms of HLA-G in different asthma sub-types to inform the development of individualized and targeted drug therapies. HLA-G has been identified as an inflammatory mediator in asthma and an asthma susceptibility gene.78 Specifically, maternal asthma status may interact with the fetal HLA-G phenotype to influence the risk for development of asthma in childhood.79 Carlini et al. reported that HLA-G expressed by the pulmonary epithelium may modulate local and distant immune responses.80,81
Tahan et al. found that plasma sHLA-G levels were not significantly different between children with asthma and healthy controls, and there were no significant correlations of plasma sHLA-G levels to eosinophil counts or total IgE levels.39 However, Ciprandi et al showed that serum sHLA-G levels were significantly higher in patients with AR or allergic asthma compared to non-allergic patients or healthy controls, and serum sHLA-G levels were related to serum allergen–specific IgE levels.38 These disparate findings may be explained by the heterogeneity of asthma airway inflammation and asthma subtypes, as well as differences in study design and sample size. Further studies are required to resolve these controversies.
IL-10 is an important cytokine that inhibits inflammation. In children with atopic asthma, plasma sHLA-G levels are significantly higher, and plasma IL-10 levels are significantly lower than healthy controls.82–84 HLA-G suppresses asthma inflammation and upregulates immune tolerance by several mechanisms, including inhibiting antigen-presenting cells (APC) and activating CD4+CD25+FoxP3 T Treg cells.37,85
The localization of HLA-G in airway epithelial cells implies that HLA-G is involved in immune disorders and airway inflammation in chronic asthma. White et al. found that the level of sHLA-G in BALF is related to the asthma subtype. High levels of sHLA-G were found in BALF of patients with low levels of eosinophils in BALF, low FENO, and low serum IgE, but there was no correlation between sHLA-G levels and blood eosinophil count.86 Gene expression data showed that HLA-G may influence or be influenced by airway epithelial cell transcripts and the ERK1/2 signaling pathway.86 HBECs from patients with mild and severe asthma display specific HLA-G expression profiles and less functional HLA-G isoforms than healthy controls.81 Asthma severity varies by subtype; consequently, these data suggest that HLA-G isoforms differ across asthma subtypes. An imbalance between HLA-G isoforms may result in an abnormal interplay of the immune response with the airway epithelium in patients with asthma; therefore, HLA-G represents a potential therapeutic target. Accordingly, treatment of asthma patients with allergen-specific immunotherapy leads to the production of HLA-G+APC or monocytes, which forms a tolerogenic microenvironment with elevated levels of IL-10 and Treg cell activation.37 Sublingual immunotherapy (SLIT) can significantly decrease serum sHLA-G in patients with asthma82 through a response that is mediated by shifting to a Th1 immune response and IFN-γ, suggesting that serum sHLA-G levels may have utility in monitoring the treatment effect of SLIT.37,55 Asthma-related exacerbations were reduced in statin users carrying the G minor allele of rs1063320 compared to noncarriers.87 In vitro, statins upregulated mir-148b and mir-152, HLA-G expression was regulated by mir-148a, mir-148b and mir-152 expression, and rs1063320 appeared to disrupt this interaction.87 PBMC produce significantly more sHLA-G in patients with isocyanate-induced asthma compared to patients with nonisocyanate-induced asymptomatic asthma. Isocyanates can trigger occupational asthma, but asthma symptoms may not improve away from the workplace.84,88 HLA-G may be a biomarker in occupational asthma and a predictor of prognosis after removal from the workplace.
Conclusion
The mechanism of action of HLA-G in asthma is unclear. Although there is increasing evidence that HLA-G is an important immunomodulatory factor in asthma, knowledge of the level and function of HLA-G in human asthma subtypes is limited, and published studies report conflicting results. Disparate research findings may be explained by the heterogeneity of asthma airway inflammation and asthma subtypes. Further studies are required to demonstrate the differences in sputum, BALF, and serum HLA-G expression across asthma subtypes. Experiments in animal models are needed to verify the effect of HLA-G on different asthma subtypes. Findings should provide information regarding the potential of HLA-G as a novel target for asthma diagnosis and treatment.
Funding
This study was funded by the National Natural Science Foundation of China (82070037), Jilin Scientific and Technological Development Program (20200201384JC).
Ethics Approval and Consent to Participate
Not applicable.
Consent for Publication
Not applicable.
Competing Interests
No competing interests.
Contributions
GP’s substantial contributions were to the conception and design of the work. He also contributed to the acquisition, analysis, and interpretation of data for the work. JYL contributed to the drafting of the work or revising it critically for important intellectual content. GP contributed to the final approval of the version to be published. GP Agreement to be accountable for all aspects of the work in ensuring that questions related to the accuracy or integrity of any part of the work are appropriately investigated and resolved. YQH, XJL, and WL contributed substantially to the acquisition and interpretation of data for the work. JYL revised the work critically for important intellectual content, and GP contributed to the drafting of the work or revising it critically for important intellectual content. All authors read and approved the final manuscript.
Data-sharing Statement
The data set (with all identifiers removed) is deposited as a data supplement at the Dryad data repository at http://datadryad.org/ with the doi:10.5061/dryad.q8s5k. The data is free for re-use by all other researchers and there are no additional forms or ethics applications that need to be completed.
REFERENCES
1. Nicolae D, Cox NJ, Lester LA, Schneider D, Tan Z, Billstrand C, et al. Fine mapping and positional candidate studies identify HLA-G as an asthma susceptibility gene on chromo-some 6p21. Am J Hum Genet. 2005;76(2):349–57. 10.1086/427763
2. Ribeyre C, Carlini F, René C, Jordier F, Picard C, Chiaroni J, et al. HLA-G haplotypes are differentially associated with asthmatic features. Front Immunol. 2018;9:278. 10.3389/fimmu.2018.00278
3. Rouas-Freiss N, LeMaoult J, Moreau P, Dausset J, Carosella ED. HLA-G in transplantation: A relevant molecule for inhibition of graft rejection? Am J Transplant. 2003;3(1):11–6. 10.1034/j.1600-6143.2003.30103.x
4. Zhang JB, Wang ZY, Chen J, Wu XD, Zhou B, Yie SM. The expression of human leukocyte antigen G (HLA-G) is associated with sacroiliitis stages of ankylosing spondylitis. Immunol Lett. 2013;152(2):121–5. 10.1016/j.imlet.2013.04.006
5. Nguyen LS, Rouas-Freiss N, Funck-Brentano C, Leban M, Carosella ED, Touraine P, et al. Influence of hormones on the immunotolerogenic molecule HLA-G: A cross-sectional study in patients with congenital adrenal hyperplasia. Eur J Endocrinol. 2019;181(5):481–8. 10.1530/EJE-19-0379
6. Fainardi E, Castellazzi M, Stignani M, Morandi F, Sana G, Gonzalez R, et al. Emerging topics and new perspectives on HLA-G. Cell Mol Life Sci. 2011;68(3):433–51. 10.1007/s00018-010-0584-3
7. Reches A, Berhani O, Mandelboim O. A unique regulation region in the 3’ UTR of HLA-G with a promising potential. Int J Mol Sci. 2020;21(3):900. 10.3390/ijms21030900
8. Riteau B, Rouas-Freiss N, Menier C, Paul P, Dausset J, Carosella ED. HLA-G2,-G3, and-G4 isoforms expressed as nonmature cell surface glycoproteins inhibit NK and antigen-specific CTL cytolysis. J Immunol. 2001;166(8):5018–26. 10.4049/jimmunol.166.8.5018
9. LeMaoult J, Le Discorde M, Rouas-Freiss N, Moreau P, Menier C, McCluskey J, et al. Biology and functions of human leukocyte antigen-G in health and sickness. Tissue Antigens. 2003;62(4):273–84. 10.1034/j.1399-0039.2003.00143.x
10. Urosevic M, Kurrer MO, Kamarashev J, Mueller B, Weder W, Burg G, et al. Human leukocyte antigen G up-regulation in lung cancer associates with high-grade histology, human leukocyte antigen class I loss and interleukin-10 production. Am J Pathol. 2001;159(3):817–24. 10.1016/S0002-9440(10)61756-7
11. Curigliano G, Criscitiello C, Gelao L, Goldhirsch A. Molecular pathways: Human leukocyte antigen G (HLA-G). Clin Cancer Res. 2013;19(20):5564–71. 10.1158/1078-0432.CCR-12-3697
12. Hackmon R, Pinnaduwage L, Zhang J, Lye SJ, Geraghty DE, Dunk CE. Definitive class I human leukocyte antigen expression in gestational placentation: HLA-F, HLA-E, HLA-C, and HLA-G in extravillous trophoblast invasion on placentation, pregnancy, and parturition. Am J Reprod Immunol. 2017;77(6):e12643. 10.1111/aji.12643
13. Pabalan N, Jarjanazi H, Sun C, Iversen AC. Meta-analysis of the human leukocyte antigen-G (HLA-G) 14 bp insertion/ deletion polymorphism as a risk factor for preeclampsia. Tissue Antigens. 2015;86(3):186–94. 10.1111/tan.12627
14. Yie SM, Li LH, Xiao R, Librach CL. A single base-pair mutation in the 3’-untranslated region of HLA-G mRNA is associated with pre-eclampsia. Mol Hum Reprod. 2008;14(11):649–53. 10.1093/molehr/gan059
15. Lin A, Yan WH. Human leukocyte antigen-g (hla-g) expression in cancers: Roles in immune evasion, metastasis and target for therapy. Mol Med. 2015;21(1):782–91. 10.2119/molmed.2015.00083
16. Ozdemir C, Kucuksezer UC, Akdis M, Akdis CA. The concepts of asthma endotypes and phenotypes to guide current and novel treatment strategies. Expert Rev Respir Med. 2018;12(9):733–43. 10.1080/17476348.2018.1505507
17. Chang HS, Lee TH, Jun JA, Baek AR, Park JS, Koo SM, et al. Neutrophilic inflammation in asthma: Mechanisms and therapeutic considerations. Expert Rev Respir Med. 2017;11(1):29–40. 10.1080/17476348.2017.1268919
18. Polosa R, Thomson NC. Smoking and asthma: Dangerous liaisons. Eur Respir J. 2013;41(3):716–26. 10.1183/09031936.00073312
19. Scott HA, Gibson PG, Garg ML, Wood LG. Airway inflammation is augmented by obesity and fatty acids in asthma. Eur Respir J. 2011;38(3):594–602. 10.1183/09031936.00139810
20. Green RH, Brightling CE, Woltmann G, Parker D, Wardlaw AJ, Pavord ID. Analysis of induced sputum in adults with asthma: Identification of subgroup with isolated sputum neutrophilia and poor response to inhaled corticosteroids. Thorax. 2002;57(10):875–9. 10.1136/thorax.57.10.875
21. Choi JS, Jang AS, Park JS, Park SW, Paik SH, Park JS, et al. Role of neutrophils in persistent airway obstruction due to refractory asthma. Respirology. 2012;17(2):322–9. 10.1111/j.1440-1843.2011.02097.x
22. Shaw DE, Berry MA, Hargadon B, McKenna S, Shelley MJ, Green RH, et al. Association between neutrophilic airway inflammation and airflow limitation in adults with asthma. Chest. 2007;132(6):1871–5. 10.1378/chest.07-1047
23. Berry MA, Shaw DE, Green RH, Brightling CE, Wardlaw AJ, Pavord ID. The use of exhaled nitric oxide concentration to identify eosinophilic airway inflammation: An observational study in adults with asthma. Clin Exp Allergy. 2005;35(9):1175–9. 10.1111/j.1365-2222.2005.02314.x
24. Berry M, Morgan A, Shaw DE, Parker D, Green R, Brightling C, et al. Pathological features and inhaled corticosteroid response of eosinophilic and non-eosinophilic asthma. Thorax. 2007; 62(12):1043–9. 10.1136/thx.2006.073429
25. Wang F, He XY, Baines KJ, Gunawardhana LP, Simpson JL, Li F, et al. Different inflammatory phenotypes in adults and children with acute asthma. Eur Respir J. 2011;38(3):567–74. 10.1183/09031936.00170110
26. Ntontsi P, Loukides S, Bakakos P, Kostikas K, Papatheodorou G, Papathanassiou E, et al. Clinical, functional and inflammatory characteristics in patients with paucigranulocytic stable asthma: Comparison with different sputum phenotypes. Allergy. 2017;72(11):1761–7. 10.1111/all.13184
27. Hastie AT, Moore WC, Meyers DA, Vestal PL, Li H, Peters SP, et al. Analyses of asthma severity phenotypes and inflammatory proteins in subjects stratified by sputum granulocytes. J Allergy Clin Immunol. 2010;125(5):1028–36.e13. 10.1016/j.jaci.2010.02.008
28. Negrini S, Contini P, Pupo F, Greco M, Murdaca G, Puppo F. Expression of membrane-bound human leucocyte antigen-G in systemic sclerosis and systemic lupus erythematosus. Hum Immunol. 2020;81(4):162–7. 10.1016/j.humimm.2019.12.004
29. Ghavimi R, Alsahebfosoul F, Salehi R, Kazemi M, Etemadifar M, Zavaran Hosseini A. High-resolution melting curve analysis of polymorphisms within CD58, CD226, HLA-G genes and association with multiple sclerosis susceptibility in a subset of Iranian population: A case-control study. Acta Neurol Belg. 2020;120(3):645–52. 10.1007/s13760-018-0992-y
30. Bae SC, Lee YH. Association of HLA-G polymorphisms with systemic lupus erythematosus and correlation between soluble HLA-G levels and the disease: A meta-analysis. Z Rheumatol. 2021;80(1):96–102. 10.1007/s00393-020-00783-6
31. Zidi I, Ben Yahia H, Bortolotti D, Mouelhi L, Laaribi AB, Ayadi S, et al. Association between sHLA-G and HLA-G 14-bp deletion/ insertion polymorphism in Crohn’s disease. Int Immunol. 2015;27(6):289–96. 10.1093/intimm/dxv002
32. Gomes RG, Brito CAA, Martinelli VF, Santos RND, Gomes F, Peixoto CA, et al. HLA-G is expressed in intestinal samples of ulcerative colitis and Crohn’s disease patients and HLA-G5 expression is differentially correlated with TNF and IL-10 cytokine expression. Hum Immunol. 2018;79(6):477–84. 10.1016/j.humimm.2018.03.006
33. Contini P, Puppo F, Canonica GW, Murdaca G, Ciprandi G. Allergen-driven HLA-G expression and secretion in peripheral blood mononuclear cells from allergic rhinitis patients. Hum Immunol. 2016;77(12):1172–8. 10.1016/j.humimm.2016.08.005
34. Negrini S, Contini P, Murdaca G, Puppo F. HLA-G in allergy: Does it play an immunoregulatory role? Front Immunol. 2021;12:789684. 10.3389/fimmu.2021.789684
35. Avelino MA, Wastowski IJ, Ferri RG, Elias TG, Lima AP, Marinho LC, et al. The human leukocyte antigen G molecule (HLA-G) expression in patients with nasal polyposis. Braz J Otorhinolaryngol. 2014;80(3):208–12. 10.1016/j.bjorl.2014.01.001
36. Ciprandi G, De Amici M, Caimmi S, Marseglia A, Marchi A, Castellazzi AM, et al. Soluble serum HLA-G in children with allergic rhinitis and asthma. J Biol Regul Homeost Agents. 2010;24(2):221–4.
37. Murdaca G, Contini P, Negrini S, Ciprandi G, Puppo F. Immunoregulatory role of hla-g in allergic diseases. J Immunol Res. 2016;2016:6865758. 10.1155/2016/6865758
38. Ciprandi G, Corsico A, Pisati P. Serum-soluble HLA-G is associated with specific IgE in patients with allergic rhinitis and asthma. Inflammation. 2014;37(5):1630–4. 10.1007/s10753-014-9890-5
39. Tahan F, Patiroglu T. Plasma soluble human leukocyte antigen G levels in asthmatic children. Int Arch Allergy Immunol. 2006;141(3):213–6. 10.1159/000095290
40. Dorling A, Monk NJ, Lechler RI. HLA-G inhibits the transendothelial migration of human NK cells. Eur J Immunol. 2000;30(2):586–93. 10.1002/1521-4141(200002)30:2<586::AID-IMMU586>3.0.CO;2-Y
41. Rajagopalan S. Endosomal signaling and a novel pathway defined by the natural killer receptor KIR2DL4 (CD158d). Traffic. 2010;11(11):1381–90. 10.1111/j.1600-0854.2010.01112.x
42. Chen XY, Yan WH, Lin A, Xu HH, Zhang JG, Wang XX. The 14 bp deletion polymorphisms in HLA-G gene play an important role in the expression of soluble HLA-G in plasma. Tissue Antigens. 2008;72(4):335–41. 10.1111/j.1399-0039.2008.01107.x
43. Jacquier A, Dumont C, Carosella ED, Rouas-Freiss N, LeMaoult J. Cytometry-based analysis of HLA-G functions according to ILT2 expression. Hum Immunol. 2020;81(4):168–77. 10.1016/j.humimm.2020.02.001
44. Melo-Lima BL, Poras I, Passos GA, Carosella ED, Donadi EA, Moreau P. The Autoimmune Regulator (Aire) transactivates HLA-G gene expression in thymic epithelial cells. Immunology. 2019;158(2):121–35. 10.1111/imm.13099
45. White SR. Human leucocyte antigen-G: Expression and function in airway allergic disease. Clin Exp Allergy. 2012; 42(2):208–17. 10.1111/j.1365-2222.2011.03881.x
46. Dumont C, Jacquier A, Verine J, Noel F, Goujon A, Wu CL, et al. CD8(+)PD-1(-)ILT2(+) T cells are an intratumoral cytotoxic population selectively inhibited by the immune-checkpoint HLA-G. Cancer Immunol Res. 2019;7(10):1619–32. 10.1158/2326-6066.CIR-18-0764
47. Bainbridge DR, Ellis SA, Sargent IL. HLA-G suppresses proliferation of CD4(+) T-lymphocytes. J Reprod Immunol. 2000;48(1): 17–26. 10.1016/S0165-0378(00)00070-X
48. Ketroussi F, Giuliani M, Bahri R, Azzarone B, Charpentier B, Durrbach A. Lymphocyte cell-cycle inhibition by HLA-G is mediated by phosphatase SHP-2 and acts on the mTOR pathway. PLoS One. 2011;6(8):e22776. 10.1371/journal.pone.0022776
49. Naji A, Durrbach A, Carosella ED, Rouas-Freiss N. Soluble HLA-G and HLA-G1 expressing antigen-presenting cells inhibit T-cell alloproliferation through ILT-2/ILT-4/FasL-mediated pathways. Hum Immunol. 2007;68(4):233–9. 10.1016/j.humimm.2006.10.017
50. Ajith A, Portik-Dobos V, Nguyen-Lefebvre AT, Callaway C, Horuzsko DD, Kapoor R, et al. HLA-G dimer targets Granzyme B pathway to prolong human renal allograft survival. FASEB J. 2019;33(4):5220–36. 10.1096/fj.201802017R
51. Pankratz S, Ruck T, Meuth SG, Wiendl H. CD4(+)HLA-G(+) regulatory T cells: Molecular signature and pathophysiological relevance. Hum Immunol. 2016;77(9):727–33. 10.1016/j.humimm.2016.01.016
52. Saurabh A, Chakraborty S, Kumar P, Mohan A, Bhatnagar AK, Rishi N, et al. Inhibiting HLA-G restores ifn-γ and tnf-α producing t cell in pleural tuberculosis. Tuberculosis. 2018;109:69–79. 10.1016/j.tube.2018.01.008
53. Morandi F, Ferretti E, Bocca P, Prigione I, Raffaghello L, Pistoia V. A novel mechanism of soluble HLA-G mediated immune modulation: Downregulation of T cell chemokine receptor expression and impairment of chemotaxis. PLoS One. 2010 Jul 23;5(7):e11763. 10.1371/journal.pone.0011763
54. Kubo M. Innate and adaptive type 2 immunity in lung allergic inflammation. Immunol Rev. 2017;278(1):162–72. 10.1111/imr.12557
55. Ciprandi G, Continia P, Fenoglio D, Sormani MP, Negrini S, Puppo F, et al. Relationship between soluble HLA-G and HLAA,-B,-C serum levels, and interferon-gamma production after sublingual immunotherapy in patients with allergic rhinitis. Hum Immunol. 2008;69(7):409–13. 10.1016/j.humimm.2008.05.009
56. Vianna P, Mondadori AG, Bauer ME, Dornfeld D, Chies JA. HLA-G and CD8+ regulatory T cells in the inflammatory environment of pre-eclampsia. Reproduction. 2016;152(6):741–51. 10.1530/REP-15-0608
57. Liang S, Ristich V, Arase H, Dausset J, Carosella ED, Horuzsko A. Modulation of dendritic cell differentiation by HLA-G and ILT4 requires the IL-6--STAT3 signaling pathway. Proc Natl Acad Sci U S A. 2008;105(24):8357–62. 10.1073/pnas.0803341105
58. Ristich V, Liang S, Zhang W, Wu J, Horuzsko A. Tolerization of dendritic cells by HLA-G. Eur J Immunol. 2005;35(4):1133–42. 10.1002/eji.200425741
59. Hazenberg MD, Otto SA, van Benthem BH, Roos MT, Coutinho RA, Lange JM, et al. Persistent immune activation in HIV-1 infection is associated with progression to AIDS. AIDS. 2003;17(13):1881–8. 10.1097/00002030-200309050-00006
60. Zhang S, Gan J, Chen BG, Zheng D, Zhang JG, Lin RH, et al. Dynamics of peripheral immune cells and their HLA-G and receptor expressions in a patient suffering from critical COVID-19 pneumonia to convalescence. Clin Transl Immunol. 2020;9(5):e1128. 10.1002/cti2.1128
61. Zhang WQ, Xu DP, Liu D, Li YY, Ruan YY, Lin A, et al. HLA-G1 and HLA-G5 isoforms have an additive effect on NK cytolysis. Hum Immunol. 2014;75(2):182–9. 10.1016/j.humimm.2013.11.001
62. Banerjee PP, Pang L, Soldan SS, Miah SM, Eisenberg A, Maru S, et al. KIR2DL4-HLAG interaction at human NK cell-oligodendrocyte interfaces regulates IFN-γ-mediated effects. Mol Immunol. 2019;115:39–55. 10.1016/j.molimm.2018.09.027
63. Baudhuin J, Migraine J, Faivre V, Loumagne L, Lukaszewicz AC, Payen D, et al. Exocytosis acts as a modulator of the ILT4-mediated inhibition of neutrophil functions. Proc Natl Acad Sci U S A. 2013;110(44):17957–62. 10.1073/pnas.1221535110
64. Morandi F, Ferretti E, Castriconi R, Dondero A, Petretto A, Bottino C, et al. Soluble HLA-G dampens CD94/NKG2A expression and function and differentially modulates chemotaxis and cytokine and chemokine secretion in CD56bright and CD56dim NK cells. Blood. 2011;118(22):5840–50. 10.1182/blood-2011-05-352393
65. Tilburgs T, Evans JH, Crespo  C, Strominger JL. The HLA-G cycle provides for both NK tolerance and immunity at the maternal-fetal interface. Proc Natl Acad Sci U S A. 2015;112(43):13312–7. 10.1073/pnas.1517724112
66. Ishikawa M, Brooks AJ, Fernández-Rojo MA, Medina J, Chhabra Y, Minami S, et al. Growth hormone stops excessive inflammation after partial hepatectomy, allowing liver regeneration and survival through induction of h2-bl/hla-g. Hepatology. 2021;73(2):759–75. 10.1002/hep.31297
67. Lee CL, Guo Y, So KH, Vijayan M, Guo Y, Wong VH, et al. Soluble human leukocyte antigen G5 polarizes differentiation of macrophages toward a decidual macrophage-like phenotype. Hum Reprod. 2015;30(10):2263–74. 10.1093/humrep/dev196
68. Esquivel EL, Maeda A, Eguchi H, Asada M, Sugiyama M, Manabe C, et al. Suppression of human macrophage-mediated cytotoxicity by transgenic swine endothelial cell expression of HLA-G. Transpl Immunol. 2015;32(2):109–15. 10.1016/j.trim.2014.12.004
69. Melén E, Kho AT, Sharma S, Gaedigk R, Leeder JS, Mariani TJ, et al. Expression analysis of asthma candidate genes during human and murine lung development. Respir Res. 2011;12(1): 86. 10.1186/1465-9921-12-86
70. Hough KP, Curtiss ML, Blain TJ, Liu RM, Trevor J, Deshane JS, et al. Airway remodeling in asthma. Front Med. 2020;7:191. 10.3389/fmed.2020.00191
71. Schleich F, Frix AN, Paulus V, Guissard F, Sanchez CE, Henket ME, et al. Asthma: The contribution of biotherapies. Rev Med Liege. 2020;75(5–6):350–5.
72. Gauthier M, Ray A, Wenzel SE. Evolving concepts of asthma. Am J Respir Crit Care Med. 2015;192(6):660–8. 10.1164/rccm.201504-0763PP
73. Asayama K, Kobayashi T, D’Alessandro-Gabazza CN, Toda M, Yasuma T, Fujimoto H, et al. Protein S protects against allergic bronchial asthma by modulating Th1/Th2 balance. Allergy. 2020;75(9):2267–78. 10.1111/all.14261
74. Hynes G, Pavord ID. Targeted biologic therapy for asthma. Br Med Bull. 2020;133(1):16–35. 10.1093/bmb/ldaa004
75. Tanaka A, Sato H, Akimoto K, Matsunaga T, Sagara H. Spontaneous sputum discriminates inflammatory phenotypes in patients with asthma. Ann Allergy Asthma Immunol. 2021;126(1):54–60.e1. 10.1016/j.anai.2020.06.017
76. Gao P, Gibson PG, Baines KJ, Yang IA, Upham JW, Reynolds PN, et al. Anti-inflammatory deficiencies in neutrophilic asthma: Reduced galectin-3 and IL-1RA/IL-1β. Respir Res. 2015;16(1):5. 10.1186/s12931-014-0163-5
77. Zhang L, Zhang J, Gao P. The potential of interleukin-37 as an effective therapeutic agent in asthma. Respir Res. 2017;18(1):192. 10.1186/s12931-017-0675-x
78. Ober C. HLA-G: An asthma gene on chromosome 6p. Immunol Allergy Clin North Am. 2005;25(4):669–79. 10.1016/j.iac.2005.08.001
79. Nicodemus-Johnson J, Laxman B, Stern RK, Sudi J, Tierney CN, Norwick L, et al. Maternal asthma and microRNA regulation of soluble HLA-G in the airway. J Allergy Clin Immunol. 2013;131(6):1496–503. 10.1016/j.jaci.2013.01.037
80. White SR, Loisel DA, Stern R, Laxman B, Floreth T, Marroquin BA. Human leukocyte antigen-G expression in differentiated human airway epithelial cells: Lack of modulation by Th2-associated cytokines. Respir Res. 2013;14(1):4. 10.1186/1465-9921-14-4
81. Carlini F, Picard C, Garulli C, Piquemal D, Roubertoux P, Chiaroni J, et al. Bronchial epithelial cells from asthmatic patients display less functional hla-g isoform expression. Front Immunol. 2017;8:6. 10.3389/fimmu.2017.00006
82. Zheng XQ, Li CC, Xu DP, Lin A, Bao WG, Yang GS, et al. Analysis of the plasma soluble human leukocyte antigen-G and interleukin-10 levels in childhood atopic asthma. Hum Immunol. 2010;71(10):982–7. 10.1016/j.humimm.2010.06.018
83. Gregori S, Tomasoni D, Pacciani V, Scirpoli M, Battaglia M, Magnani CF, et al. Differentiation of type 1 T regulatory cells (Tr1) by tolerogenic DC-10 requires the IL-10-dependent ILT4/HLA-G pathway. Blood. 2010;116(6):935–44. 10.1182/blood-2009-07-234872
84. Mapp CE, Ferrazzoni S, Rizzo R, Miotto D, Stignani M, Boschetto P, et al. Soluble human leucocyte antigen-G and interleukin-10 levels in isocyanate-induced asthma. Clin Exp Allergy. 2009;39(6):812–9. 10.1111/j.1365-2222.2009.03215.x
85. Carosella ED, Gregori S, LeMaoult J. The tolerogenic interplay(s) among HLA-G, myeloid APCs, and regulatory cells. Blood. 2011;118(25):6499–505. 10.1182/blood-2011-07-370742
86. White SR, Nicodemus-Johnson J, Laxman B, Denner DR, Naureckas ET, Hogarth DK, et al. Elevated levels of soluble humanleukocyte antigen-G in the airways are a marker for a low-inflammatory endotype of asthma. J Allergy Clin Immunol. 2017;140(3):857–60. 10.1016/j.jaci.2017.02.031
87. Naidoo D, Wu AC, Brilliant MH, Denny J, Ingram C, Kitchner TE, et al. A polymorphism in HLA-G modifies statin benefit in asthma. Pharmacogenomics J. 2015;15(3):272–7. 10.1038/tpj.2014.55
88. Kenyon NJ, Morrissey BM, Schivo M, Albertson TE. Occupational asthma. Clin Rev Allergy Immunol. 2012;43(1–2): 3–13. 10.1007/s12016-011-8272-0