
Download
ORIGINAL ARTICLE
6'-O-galloylpaeoniflorin alleviates inflammation and oxidative stress in pediatric pneumonia through activating Nrf2 activation
Cheng Xu*, Lei Song, Weiyan Zhang, Rong Zou, Meijun Zhu
Department of Pediatrics, Nantong First People's Hospital (The Second Affiliated Hospital of Nantong University), Nantong, Jiangsu Province, China
Abstract
Objective: To assess the therapeutic effect and mechanism of 6'-o-galloylpaeoniflorin (GPF) in pediatric pneumonia.
Methods: The effects of lipopolysaccharide (LPS) and GPF on cell viability and apoptosis were examined by cell counting kit-8 assay and flow cytometry analysis. The oxidative stress and inflammatory response were assessed by detecting expression levels of superoxide dismutase, glutathione, r-glutamyl cysteingl+glycine, myeloperoxidase, and malondialdehyde as well as tumor necrosis factor-α, Interleukin-18, and Interleukin-10 by using enzyme-linked-immunosorbent serologic assay. Moreover, the activation of nuclear factor erythroid 2-related factor 2 (Nrf2) pathway was detected by immunoblot assay, and the influence of Nrf2-knockdown on cell viability, oxidative stress, and inflammation response was also investigated.
Results: The results established that GPF increased the viability of LPS-induced pneumonia cells. In addition, GPF reduced LPS-induced oxidative stress in pneumonia cells. It was further discovered that GPF reduced LPS-induced inflammation in pneumonic cell. GPF improved the activity of Nrf2 in LPS-treated pneumonic cells, and therefore alleviated inflammation and oxidative stress in pediatric pneumonia.
Conclusion: GPF could serve as a promising drug for treating pediatric pneumonia.
Key words: LPS, Nrf2, oxidative stress, pediatric pneumonia, 6'-o-galloylpaeoniflorin (GPF)
*Corresponding author: Cheng Xu, Department of Pediatrics, Nantong First People’s Hospital (The Second Affiliated Hospital of Nantong University), No. 6 North Road, Child Lane, Chongchuan District, Nantong City, Jiangsu Province 226001, China. Email address: [email protected]
Received 31 March 2022; Accepted 20 April 2022; Available online 1 July 2022
Copyright: Xu C, et al.
License: This open access article is licensed under Creative Commons Attribution 4.0 International (CC BY 4.0). http://creativecommons.org/licenses/by/4.0/
Introduction
The World Health Organization has estimateds that there are 150.7 million cases of lung infections in children aged less than 5 years with lung infections, with of which as many as 20 million patients require hospitalization.1 Pediatric pneumonia is one of the most common pediatric diseases leading to infant death. It is primarily caused by bacterial and viral infections and its primary clinical manifestations include dyspnea, fever, cough, and shortness of breath.2,3 Pneumonia in children can result in lung tissue destruction, neutrophil infiltration, and release of pro-inflammatory cytokines such as tumor necrosis factor (TNF)-α, interleukin (IL)-1β, and IL-6.4 Activated neutrophils aggravate inflammation and injury by producing reactive oxygen species (ROS) and proteolytic enzymes. Lipopolysaccharide (LPS) is the prime component of endotoxin, and is considered as the most important pathogen leading to lung inflammation and sepsis.5 LPS activates a large number of inflammatory cells and induces pneumonia through a variety of signaling pathways. Therefore, developing new drugs to combat pneumonia in children is urgently required.
Herbs such as Urtica dioica (stinging nettle) have exhibited beneficial anti-inflammatory functions. For example, comparison of the substrate specificities of endo--beta-N- acetylglucosaminidases from Streptomyces griseus and Diplococcus pneumoniae has been revised. The beneficial effects of Urtica dioica on the function, histology, and molecular changes in the hippocampus of streptozotocin (STZ)-induced diabetic mice have been reported.6,7
6'-O-galloylpaeoniflorin (GPF) is an acylated monoterpenoid glucoside compound extracted from the roots of peony plant (genus Paeonia). It is composed of d-glucose, benzoyl, and gallic parts, and has antitumor, antiplatelet aggregation, anticoagulant, and antiallergic potential, with ability to promote angiogenesis.8 In addition, it can alleviate inflammatory responses, cell damage, and apoptosis in vitro, and has free radical scavenging and antioxidant properties.9 GPF attenuates osteoclast generation and alleviates osteoporosis by inhibiting production of reactive oxygen species (ROS) and MAPKs/C-FOS/NFATc1 signaling pathway, and alleviates neuroinflammation and oxidative stress induced by cerebral ischemia-reperfusion through activating phosphoinositide 3-kinase (PI3K)/Ak strain transforming (Akt)/nuclear factor erythroid 2-related factor 2 (Nrf2) pathway.8 GPF attenuates osteoclastogenesis and relieves osteoporosis via suppressing ROS generation and MAPKs/c-Fos/NFATc1 pathway.8 However, GPF has not been reported to relieve pneumonia in children.
In this study, pneumonia cell model was constructed by LPS stimulation. Our data demonstrated that GPF increased the activity of LPS-treated pneumonia cells. GPF reduced LPS-induced oxidative stress in pneumonia cells. In addition, GPF reduced LPS-induced inflammation in pneumonic cells. Mechanically, we found that GPF improved the activity of Nrf2 in LPS-treated pneumonic cells, and therefore alleviated inflammation and oxidative stress in pediatric pneumonia. We anticipated that GPF could serve as a promising drug for treating pediatric pneumonia.
Materials and Methods
Cell culture
Human bronchial epithelial (Beas-2B) cells (the Cell Resource Center of Peking Union Medical College, China) were incubated with RPMI-1640 medium (Gibco; Thermo Fisher Scientific Inc., Waltham, MA. USA) and 10% fetal bovine serum (FBS; Thermo Fisher Scientific Inc.) supplemented with 1% penicillin–streptomycin in a humidified culture hood with 5% CO2 at 37°C. The lung injury was achieved by LPS stimulation at a concentration of 5 µg/mL for 12 h at 37°C. Cells were stimulated with increasing dose of GPF. For Nrf2 knockdown, small interfering RNAs (siRNAs) targeting Nrf2 (si-Nrf2) (5'-CCGGCAUUUCACUAAACACAATT-3’) were generated by GenePharma (Shanghai, China) and transfected into Beas-2B cells with lipofectamine 3000 (Invitrogen, Carlsbad, CA) for 48 h.
Cell Counting Kit-8 (CCK-8) assay
For assessing cell viability, CCK-8 assay was performed. After indicated treatment, Beas-2B cells were added to 10-μL CCK-8 solution in each well. After incubation for 1 h at 37°C, absorbance in each well was determined at 450 nm using a microplate reader (R&D Systems, Minneapolis, MN) at 450 nm.
Flow cytometry (FCM)
Cells were placed into 96-well plate and treated as designed. Then the cells were fixed and labeled with 50-μg/mL propidium iodide (PI) at room temperature for 30 min in dark. Then stained cells were washed by centrifugation and resuspended, followed by analysis on a BD FACSCanto™ II system (BD Biosciences, San Diego, California).
Western blotting
Cells were lysed with radioimmunoprecipitation assay (RIPA) buffer. After collection of supernatant, protein concentration was determined by bicinchoninic (BCA) protein assay kit (Beyotime Biotechnology, Shanghai, China). Then, proteins were separated by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE), and transferred onto polyvinylidene difluoride (PVDF) membranes. After that, membranes were incubated with 5% bovine serum albumin (BSA), followed by primary antibodies targeting Bcl-2 (1:1000, Abcam, Cambridge, UK), Bax (1:1000, Abcam), Nrf2 (1:1000, Abcam), NQO-1 (1:1000, Abcam), HO-1 (1:1000, Abcam), and β-actin (1:10000; Abcam). Membranes were maintained in horseradish peroxidase (HRP)-conjugated secondary antibodies at a dilution of 1:1000 for 2 h. The membranes were rinsed in tris buffered saline with tween (TBST) for 15 min before the signals were detected with Luminata Creseendo Western HRP substrate via Molecular Imager ChemiDoc XRS+ System (Bio-Rad, Philadelphia, PA).
Detection of antioxidant activity
Levels of superoxide dismutase (SOD), glutathione, r---glutamyl cysteingl + glycine (GSH), myeloperoxidase (MPO), and malondialdehyde (MDA) were measured using detection kits obtained from Nanjing Jiancheng Bio-Engineering Institute (Jiangsu, China) and following the manufacturer’s instructions.
Enzyme-linked-immunosorbent serologic assay (ELISA)
Concentrations of TNF-α, IL-18, and IL-10 in cell lysates were measured by ELISA kit following manufacturer’s protocols. Briefly, samples were added into wells, and biotin-conjugated primary antibodies were placed prior to addition of avidin-conjugated HRP. Then, enzyme substrate was added for chromogenic reaction. Intensity of each well was measured using a microplate reader (R&D Systems). All experiments were conducted according to the stated guidelines.
Statistical analysis
Statistical analysis was conducted by unpaired two-tailed Student’s t-test using the SPSS software. Data were displayed as mean ± standard error of mean (SEM), and P ˂ 0.05 was considered as significant difference.
Results
GPF could improve LPS-treated Beas-2B cell viability
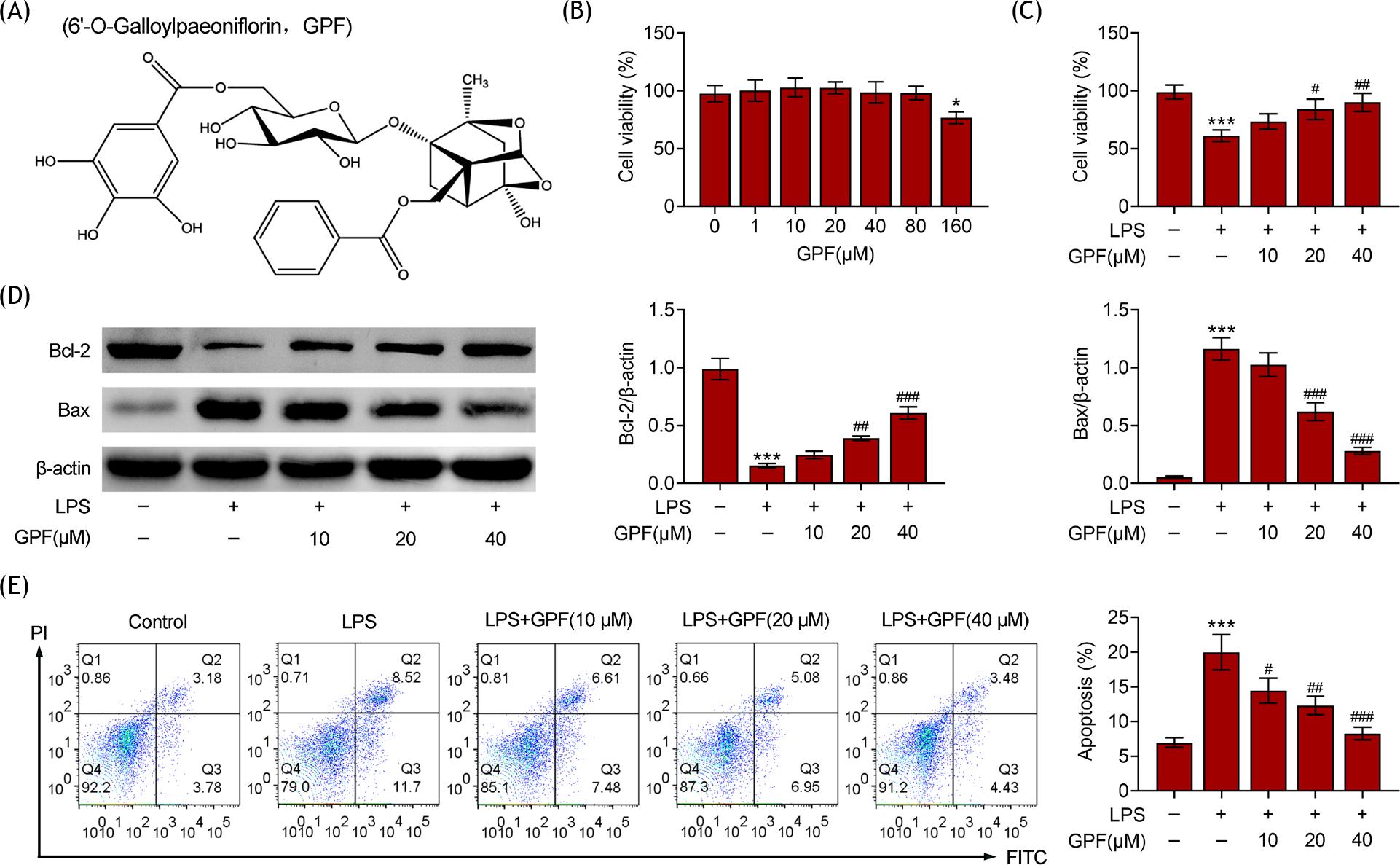
The cell viability of Beas-2B cells in response to LPS and increasing concentration of GPF was examined. It was noted that cells stimulated by varying doses of GPF demonstrated no significant difference in cell viability, indicating that GPF had no toxicity in cell viability (Figures 1A and 1B). The cell viability was decreased in cells treated with GPF at a concentration of 160 µM (Figure 1B). The cell viability was also decreased post-stimulation with LPS. However, GPF treatment relieved cell viability in LPS-stimulated cells (Figure 1C). Expressions of Bcl-2 and Bax were suppressed and enhanced, respectively, in LPS-treated cells, and were significantly reversed by GPF treatment in a dose-dependent manner (Figure 1D). In addition, the apoptosis was examined by flow cytometry. LPS stimulation significantly increased the percentage of apoptotic cells, while GPF significantly reduced apoptotic cell numbers (Figure 1E). Thus, GPF was proved to enhance cell viability and relieve cell apoptosis in Beas-2B cells.
Figure 1 GPF improves LPS-induced decrease in cell viability. (A) The chemical structure of GPF. (B) Cell viability of cells treated with increasing levels of GPF in control, LPS, LPS+GPF (10 µM), LPS+GPF (20 µM), LPS+GPF (40 µM), LPS+GPF (80 µM), and LPS+GPF (160 µM). (C) Cell viability in control, LPS, LPS+GPF (10 µM), LPS+GPF (20 µM), and LPS+GPF (40 µM). (D) Expression levels of Bcl-2 and Bax in control, LPS, LPS+GPF (10 µM), LPS+GPF (20 µM), and LPS+GPF (40 µM) group. (E) Cell apoptosis as detected by flow cytometry in control, LPS, LPS+GPF (10 µM), LPS+GPF (20 µM), and LPS+GPF (40 µM) groups. ***P ˂ 0.001 vs. control; #P ˂ 0.05, ##P ˂ 0.01, and ###P ˂ 0.001 vs. LPS.
GPF ameliorated oxidative stress induced by LPS
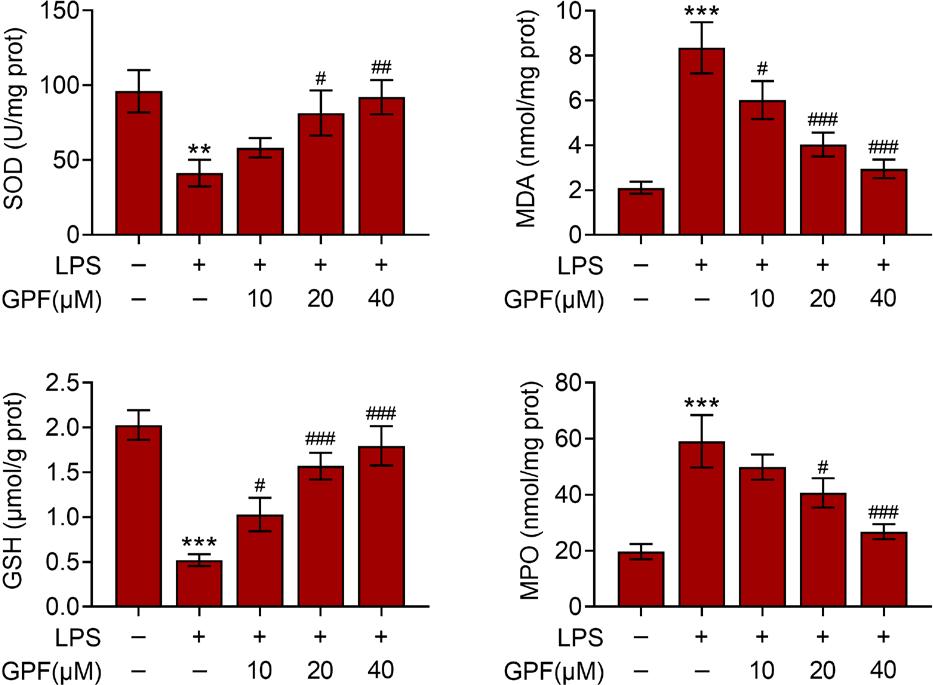
Lipopolysaccharide stimulation is believed to induce oxidative stress. Therefore, oxidative response in Beas-2B cells stimulated by LPS and GPF was evaluated. The expression of MDA and MPO was enhanced but that of SOD and GSH was reduced in LPS-stimulated cells (Figure 2). GPF treatment significantly reversed the effect of LPS on the expression of of SOD, MDA, MPO, and GSH in a dose-dependent manner, indicating its antioxidant effects.
Figure 2 GPF ameliorates LPS-induced oxidative stress. Levels of SOD, MDA, GSH, and MPO in control, LPS, LPS+GPF (10 µM), LPS+GPF (20 µM), and LPS+GPF (40 µM) groups. ***P ˂ 0.001 vs. control; #P ˂ 0.05, ##P ˂ 0.01, and ###P ˂ 0.001 vs. LPS.
GPF reduced LPS-induced inflammation in -pneumonic cells
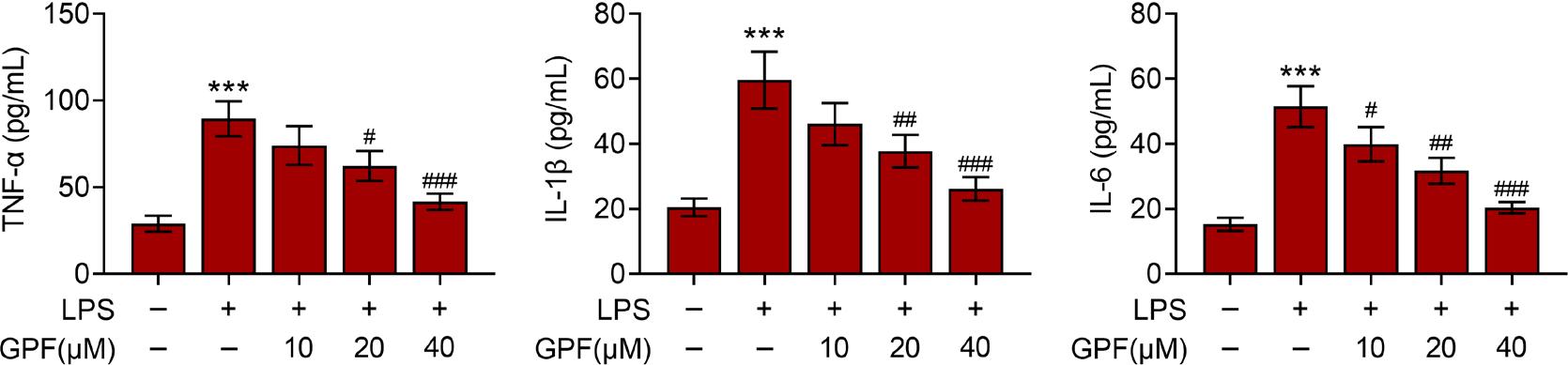
In order to explore inflammatory response in LPS-treated cells, levels of IL-6, IL-1β, and TNF-α were monitored. LPS significantly increased the levels of IL-6, IL-1β, and TNF-α (Figure 3). It was observed that administration of GPF alleviated cellular inflammation as identified by reduction of these cytokines (Figure 3).
Figure 3 GPF alleviates LPS-induced inflammation in pneumonic cells. (A–D) Levels of TNF-α IL-1β, and IL-6 in control, LPS, LPS+GPF (10 µM), LPS+GPF (20 µM), and LPS+GPF (40 µM) groups. ***P ˂ 0.001 vs. control; #P ˂ 0.05, ##P ˂ 0.01, and ###P ˂ 0.001 vs. LPS.
GPF improved the activity of Nrf2 in LPS-treated pneumonic cells
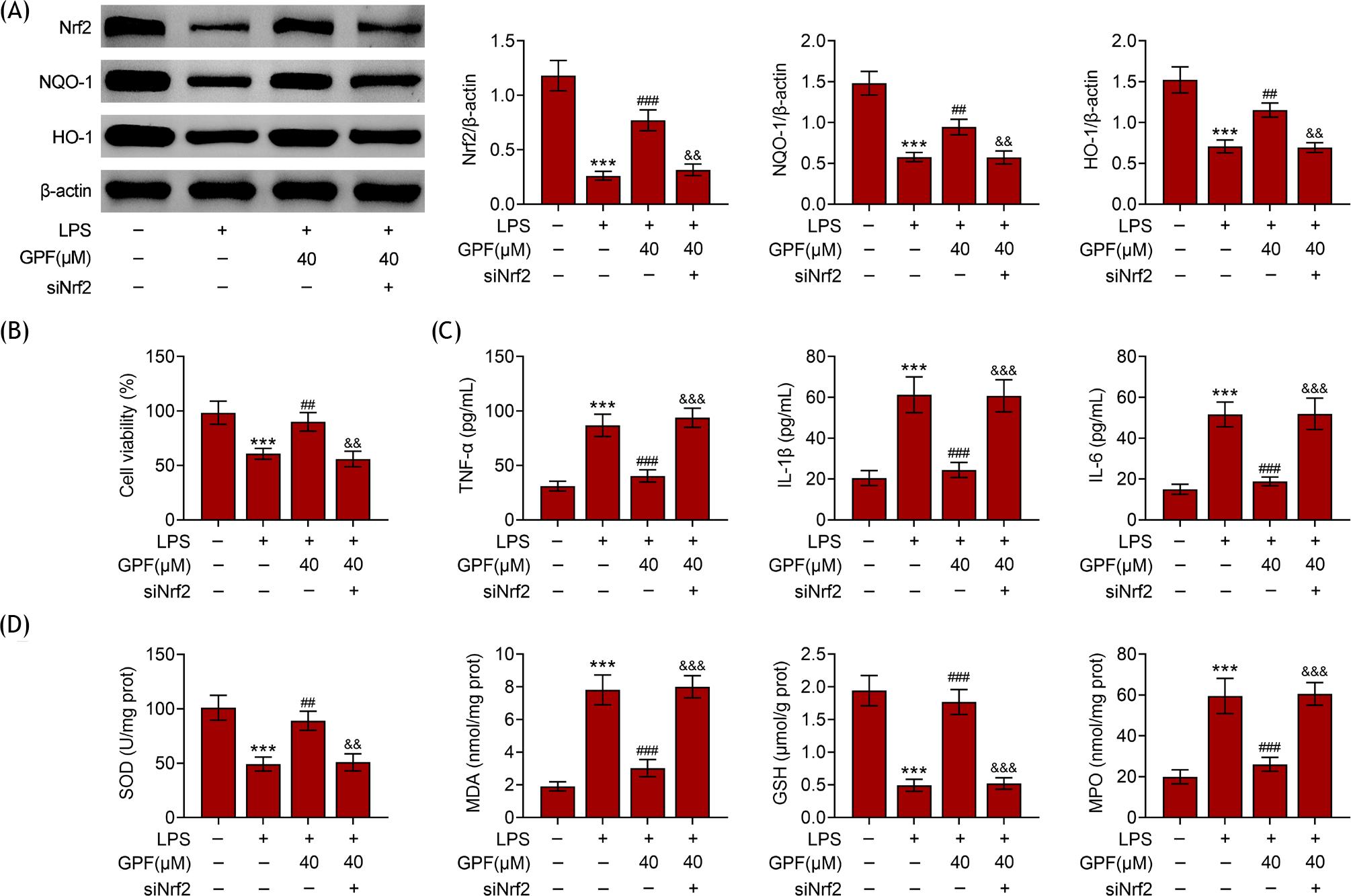
In order to delineate the potential mechanism of GPF-mediated cell proliferation and apoptosis recovery, the expression level of Nrf2 was detected in each group. The expression levels of Nrf2, NQO-1, and HO-1 were increased in LPS-treated cells (Figure 4A). GPF treatment decreased the expression levels of Nrf2, NQO-1, and HO-1 (Figure 4A). However, the effect of GPF was abolished by Nrf2 knockdown. Moreover, the perfection effects of GPF on cell viability and inflammation were destroyed by Nrf2 ablation (Figures 4B and 4C). Similarly, the antioxidant effects of GPF were weakened by ablation of Nrf2, suggesting the involvement of Nrf2 in GPF-mediated anti-inflammatory and antioxidant properties in pediatric pneumonia (Figure 4D).
Figure 4 GPF improved the activity of Nrf2 in LPS-induced pneumonic cells. (A) Levels of Nrf2, NQO-1, and HO-1 in control, LPS, LPS+GPF (40 µM), and LPS+GPF+siNrf2 (40 µM) groups. (B) Cell viability in Control, LPS, LPS+GPF (40 µM), and LPS+GPF+siNrf2 (40 µM) groups. (C) Oxidative stress in control, LPS, LPS+GPF (40 µM), and LPS+GPF+siNrf2 (40 µM) groups. (D) Inflammation response in control, LPS, LPS+GPF (40 µM), and LPS+GPF+siNrf2 (40 µM) groups. ***P ˂ 0.001 vs. control; #P ˂ 0.05, ##P ˂ 0.01, and ###P ˂ 0.001 vs. LPS.
Discussion
Pediatric pneumonia is one of the important causes of neonatal death with high morbidity and mortality.10 Bacterial infection is the fundamental cause of pneumonia in children. Therefore, it is important to thoroughly clarify the pathogenesis of pneumonia in children, carry out effective anti-inflammatory treatment, and develop effective clinical drugs to improve the cure rate of pneumonia in children.11 Up to now, although a large number of antibacterial/antiviral drugs have been developed for treating pediatric pneumonia, more of effective drugs are required to improve the therapeutic effect to combat pediatric pneumonia. Traditional Chinese medicine (TCM) has several advantages for treating pediatric pneumonia, as many TCM elements have been found to be suitable for the same.12 In this study, we discovered that GPF alleviated inflammation and oxidative stress in pediatric pneumonia. We, therefore, believed that GPF could serve as a promising drug for treating pediatric pneumonia.
Lipopolysaccharide is the prime component of endotoxin and is considered as the most important pathogen resulting in lung inflammation and sepsis.13-15 LPS activates a large number of inflammatory cells and induces pneumonia through a variety of signaling pathways.16-18 In this study, we successfully constructed a pneumonia model using LPS, and confirmed the effects of GPF on this model.
Results of CCK-8 and FCM assays demonstrated that GPF increased the viability of LPS-induced pneumonia cells. ELISA demonstrated that GPF reduced LPS-induced oxidative stress and inflammation in pneumonia cells. The results confirmed that GPF could serve as a promising drug for treating pediatric pneumonia. Multiple biological activities of GPF have been unraveled in several types of diseases.8,19 GPF attenuated osteoclastogenesis as well as relieved osteoporosis by suppressing MAPKs/c-Fos/NFATc1 pathway.20 GPF regulated growth and metastasis of non-small cell lung cancer (NSCLS) by regulating the AMPK/miR-299-5p/ATF2 pathway.19 In addition, GPF could protect keratinocytes against oxidative stress-induced cell damage.21 Similarly, we also discovered that GPF suppressed oxidative stress in pediatric pneumonia. It could also reduce neuroinflammation and oxidative stress induced by cerebral ischemia-reperfusion through activating PI3K/Akt/Nrf2 pathway.9 Here, GPF also improved the activity of Nrf2 in LPS-induced pneumonic cells, and hence alleviated inflammation and oxidative stress in pediatric pneumonia. However, this precise mechanism requires further study.
Pneumonia results in lung tissue destruction, neutrophil infiltration, and release of pro-inflammatory cytokines, including TNF-α, IL-1β, and IL-6 in children.22-24 Activated neutrophils spread out inflammation and injury by producing ROS and proteolytic enzymes.25,26.
The present research also revealed that GPF improved the activity of Nrf2 in LPS-induced pneumonic cells. As observed, this pathway plays a critical role in mediating inflammation and oxidative stress in different types of diseases. Several drugs or proteins could affect and control the progression of pediatric pneumonia through this pathway. We assume that the Nrf2 pathway could act as a target for pediatric pneumonia.
To summarize, we established that GPF reduced LPS-induced oxidative stress and inflammation in pneumonia cells. It also improved the activity of Nrf2 in LPS-treated pneumonic cells, and therefore alleviated pediatric pneumonia.
Competing Interests
The authors stated that there were no conflicts of interest to disclose.
Ethics Approval
This study did not contain any research involving human participants or animals performed by any of the authors.
Data Availability
The authors declare that all data supporting the findings of this study are available within the paper and any raw data can be obtained from the corresponding author upon request.
Author Contribution
Cheng Xu and Lei Song designed the experiments, and Weiyan Zhang conducted the same. Rong Zou analyzed and interpreted the data, and Meijun Zhu prepared the manuscript with contributions from all coauthors.
REFERENCES
1. Ciuca IM, Dediu M, Pop LL. Pediatric pneumonia (Ped Pne) lung ultrasound score and inflammatory markers: A pilot study. Pediatr Pulmonol. 2022;57:576–82. 10.1002/ppul.25760
2. Ayan E, Karabulut B, Unver HM. Diagnosis of pediatric pneumonia with ensemble of deep convolutional neural networks in chest x-ray images. Arab J Sci Eng. 2022;47:2123–139. 10.1007/s13369-021-06127-z
3. Camilloni A, Nati G, Maggiolini P, Romanelli A, Latina R. Chronic non-cancer pain in primary care: An Italian cross-sectional study. Signa Vitae. 2021;7:54–62.
4. Wang S, Lu B, Liu J, Gu Y. TRIM27 suppresses inflammation injuries in pediatric pneumonia by targeting TLR4/NF-kappa B signaling pathway. Allergol Immunopathol. 2022;50:33–9. 10.15586/aei.v50i2.558
5. Gravel CA, Neuman MI, Monuteaux MC, Neal JT, Miller AF, Bachur RG. Significance of sonographic subcentimeter, subpleural consolidations in pediatric patients evaluated for pneumonia. J Pediatr. 2021; 10.1016/j.jpeds.2021.12.052
6. Tarentino AL, Maley F. A comparison of the substrate specificities of endo-beta-N-acetylglucosaminidases from streptomyces griseus and diplococcus pneumoniae. Biochem Biophys Res Commun. 1975;67:455–62. 10.1016/0006-291X(75)90337-X
7. Keshvari M, Rahmati M, Mirnasouri R, Chehelcheraghi F. Effects of endurance exercise and Urtica dioica on the functional, histological and molecular aspects of the hippocampus in STZ-Induced diabetic rats. J Ethnopharmacol. 2020;256:112801. 10.1016/j.jep.2020.112801
8. Liu W, Xie G, Yuan G, Xie D, Lian Z, Lin Z, et al. 6'-O-galloylpaeoniflorin attenuates osteoclasto-genesis and relieves ovariectomy-induced osteoporosis by inhibiting reactive oxygen species and MAPKs/c-Fos/NFATc1 signaling pathway. Front Pharmacol. 2021;12:641277. 10.3389/fphar.2021.641277
9. Gao J, Song L, Xia H, Peng L, Wen Z. 6'-O-galloylpaeoniflorin regulates proliferation and metastasis of non-small cell lung cancer through AMPK/miR-299-5p/ATF2 axis. Respir Res. 2020;21:39. 10.1186/s12931-020-1277-6
10. Geanacopoulos AT, Lipsett SC, Hirsch AW, Monuteaux MC, Neuman MI. Impact of viral radiographic features on antibiotic treatment for pediatric pneumonia. J Pediat Inf Dis Soc. 2022. 10.1093/jpids/piab132
11. Esposito S, Argentiero A, Rebecchi F, Fainardi V, Pisi G, Principi N. The remaining unsolved problems for rational antibiotic therapy use in pediatric community-acquired pneumonia. Expert Opin Pharmacother. 2022;23:497–505. 10.1080/14656566.2022.2028773
12. El-Mekkawy MS, Soliman SE, Saleh NY. Soluble urokinase plasminogen activator receptor: A novel biomarker of pediatric community-acquired and hospitalacquired pneumonia. Turkish J Pediatr. 2022;64:98–109. 10.24953/turkjped.2021.4321
13. Pu Z, Shen C, Zhang W, Xie H, Wang W. Avenanthramide C from oats protects pyroptosis through dependent ROS-induced mitochondrial damage by PI3K ubiquitination and phosphorylation in pediatric pneumonia. J Agricul Food Chem. 2022;70:2339–53. 10.1021/acs.jafc.1c06223
14. Thummuri D, Guntuku L, Challa VS, Ramavat RN, Naidu VGM. Abietic acid attenuates RANKL-induced osteoclastogenesis and inflammation associated osteolysis by inhibiting the NF-KB and MAPK signaling. J Cell Physiol. 2018;234:443–53. 10.1002/jcp.26575
15. Peng Q, Zhao B, Lin J, Liu H, Zhou R, Lan D, et al. SPRC-suppresses experimental periodontitis by modulating Th17/-Treg imbalance. Front Bioengin Biotechnol. 2021;9: 737334.
16. Yin S, Ding M, Fan L, Yu X, Liang Z, Wu L, et al. Inhibition of inflammation and regulation of AQPs/ENaCs/Na(+)-K(+)-ATPase mediated alveolar fluid transport by total flavonoids extracted from nervilia fordii in lipopolysaccharide-induced acute lung injury. Front Pharmacol. 2021;12:603863. 10.3389/fphar.2021.603863
17. Xuan TQ, Gong G, Du H, Liu C, Wu Y, Bao G, et al. Protective effect of pteryxin on LPS-induced acute lung injury via modulating MAPK/NF-kappaB pathway and NLRP3 inflammasome activation. J Ethnopharmacol. 2022;286:114924.
18. Nelin LD, Jin Y, Chen B, Liu Y, Rogers LK, Reese J. Cyclooxygenase-2 deficiency attenuates lipopolysaccharide-induced inflammation, apoptosis and acute lung injury in adult mice. Am J Physiol Regul integr Comp Physiol. 2022. 10.1152/ajpregu.00140.2021
19. Yao CW, Piao MJ, Kim KC, Zheng J, Cha JW, Hyun CL, et al. Cytoprotective effects of 6'-O-galloylpaeoniflorin against ultraviolet B radiation-induced cell damage in human keratinocytes. In vitro Cell Dev Biol Anim. 2014;50:664–74. 10.1007/s11626-014-9747-0
20. Yao CW, Piao MJ, Kim KC, Zheng J, Cha JW, Hyun JW. 6'-O-galloylpaeoniflorin protects human keratinocytes against oxidative stress-induced cell damage. Biomol Ther. 2013;21,349–57. 10.4062/biomolther.2013.064
21. Wen Z, Hou W, Wu W, Zhao Y, Dong X, Bai X, et al. 6'-O-galloylpaeoniflorin attenuates cerebral ischemia reperfusion--induced neuroinflammation and oxidative stress via PI3K/Akt/Nrf2 activation. Oxid Med Cell Longev. 2018;2018:8678267. 10.1155/2018/8678267
22. Jin X, Gao X, Lan M, Li CN, Sun JM, Zhang H. Study the mechanism of peimisine derivatives on NF-kappaB inflammation pathway on mice with acute lung injury induced by lipopolysaccharide. Chem Biol Drug Design. 2021. 10.1111/cbdd.14013
23. Zhang L, Zhu XZ, Badamjav R, Zhang JZ, Kou JP, Yu BY, et al. Isoorientin protects lipopolysaccharide-induced acute lung injury in mice via modulating Keap1/Nrf2-HO-1 and NLRP3 inflammasome pathways. Eur J Pharmacol. 2022;917:174748. 10.1016/j.ejphar.2022.174748
24. Yu YY, Li XQ, Hu WP, Cu SC, Dai JJ, Gao YN, et al. Self-developed NF-kappaB inhibitor 270 protects against LPS-induced acute kidney injury and lung injury through improving inflammation. Biomed Pharmacother Biomed Pharmacother. 2022;147:112615.
25. Yoon HS, Park CM. Chrysoeriol ameliorates COX-2 expression through NF-kappaB, AP-1 and MAPK regulation via the TLR4/MyD88 signaling pathway in LPS-stimulated murine macrophages. Exp Ther Med. 2021;22:718. 10.3892/etm.2021.10150
26. Choi DY, Lee JY, Kim MR, Woo ER, Kim YG, Kang KW. Chrysoeriol potently inhibits the induction of nitric oxide synthase by blocking AP-1 activation. J Biomed Sci. 2005;12:949–59. 10.1007/s11373-005-9028-8