
Download
ORIGINAL RESEARCH
Polyphyllin I alleviates lipopolysaccharide-induced inflammation reduces pyroptosis in BEAS-2B and HPAEC cells by inhibiting NF-κB signaling
Fangli Maoa, Aiping Wub*
aDepartment of General Practice, Fenghua District People’s Hospital, Ningbo, Zhejiang, 315500, China
bDepartment of Paediatrics, Xiangyang Central Hospital, Affiliated Hospital of Hubei University of Arts and Science, Xiangyang, Hubei, 441000, China
Abstract
Polyphyllin I is an active steroidal saponin isolated from Paris polyphylla with anti-cancer and anti-inflammatory properties. The present study investigates the role of polyphyllin I in acute lung injury. Firstly, the human bronchial epithelial cells (BEAS-2B) and human pulmonary artery endothelial cells (HPAEC) were stimulated with increasing concentrations of lipopolysaccharide at 2, 5, and 10 μg/mL. The treatment with lipopolysaccharide reduced the cell viabilities of BEAS-2B and HPAEC, downregulated superoxide dismutase (SOD) and glutathione (GSH), and up-regulated myeloperoxidase (MPO) and malondialdehyde (MDA). Moreover, the levels of TNF-α, IL-1β, and IL-6 were also up-regulated in lipopolysaccharide-treated BEAS-2B/HPAEC cells. Secondly, the lipopolysaccharide-treated cells were then incubated with different concentrations of polyphyllin I. Incubation with polyphyllin I enhanced the cell viabilities of lipopolysaccharide--treated BEAS-2B/HPAEC, up-regulated levels of SOD and GSH, and reduced MPO and MDA. Moreover, polyphyllin I reduced TNF-α, IL-1β, and IL-6 in lipopolysaccharide-treated BEAS-2B/HPAEC cells. Thirdly, the up-regulation of GSDMD-N, pro-caspase-1, and cleaved caspase-1 proteins in lipopolysaccharide-treated BEAS-2B/HPAEC cells were decreased by polyphyllin I. Polyphyllin I increased the protein expression of GSDMD-D in the lipopolysaccharide-treated BEAS-2B/HPAEC cells, and inhibited the translocation of GSDMD from cytoplasm to plasma membrane. Lastly, polyphyllin I reduced the expression of p-p65 in lipopolysaccharide-treated BEAS-2B/HPAEC cells. The over-expression of p65 counteracted with the inhibitory effects of polyphyllin I on oxidative stress and inflammation in lipopolysaccharide-treated BEAS-2B. In conclusion, polyphyllin I repressed the lipopolysaccharide-induced oxidative stress and inflammation in BEAS-2B and HPAEC, and reduced pyroptosis through inhibition of NF-κB signaling.
Key words: Polyphyllin I, lipopolysaccharide, oxidative stress, inflammation, pyroptosis, pneumonia, NF-κB
*Corresponding author: Aiping Wu, Department of Paediatrics, Xiangyang Central Hospital, Affiliated Hospital of Hubei University of Arts and Science, No. 136 Jingzhou Street, Xiangcheng District, Xiangyang, Hubei, China. Email address: [email protected]
Received 1 January 2022; Accepted 16 March 2022; Available online 1 July 2022
Copyright: Mao F and Wu A
License: This open access article is licensed under Creative Commons Attribution 4.0 International (CC BY 4.0). http://creativecommons.org/licenses/by/4.0/
Introduction
Acute lung injury and the related complication - acute respiratory distress syndrome - is associated with the development of multiple organ failure, and contributes to the death of patients with sepsis, shock, trauma, and pneumonia.1 Although multiple approaches, such as immune checkpoint inhibitors,2,3 have been clinically applied for septic patients, acute lung injury and the related complications still have a high mortality rate. Therefore, innovative strategies are urgently needed for the treatment of this disease.
Distinct risk factors such as virulence pathogens and bacterial infection induce the development of acute lung injury.4 Acute pulmonary inflammation is the main characteristic of acute lung injury. Neutrophil infiltration and the release of pro-inflammatory cytokines result in lung tissue destruction, leading to deterioration of gas exchange and impairment of alveolar-capillary barrier.5 Anti-inflammatory strategies have been considered to be beneficial for the clinical therapy of acute lung injury,6,7 and Lipopolysaccharide - main component of endotoxin - is regarded as the most important pathogen that leads to lung inflammation and sepsis.8
Lipopolysaccharide activates a large number of inflammatory cells and induces the release of pro--inflammatory cytokines, thus contributing to acute lung injury through a variety of signaling pathways.9 Toll-like receptor 4 is the main receptor of lipopolysaccharide, and lipopolysaccharide/toll-like receptor 4 complex triggers the activation of an intracellular signaling pathway, such as NF-κB, to participate in the pathogenesis of acute lung injury.10 Therefore, the alleviation of lipopolysaccharide-triggered lung inflammation reduces the acute lung injury.11
Traditional herbal medicines exert antioxidant and anti-inflammatory properties in diverse metabolic disorders.12 Moreover, polyphenols also function as plant bioactive compounds to ameliorate metabolic disorders such as non-alcoholic fatty liver disease.13 Polyphyllin I is a steroid saponin isolated from the rhizome of Paris polyphylla, and is widely used as a traditional Chinese medicine to treat fevers and headaches.14 Polyphyllin I has been reported to show anti-cancer effect in distinct tumors through promoting autophagy, cell apoptosis, and cell cycle, as well as modulation of the inflammatory response.15 For example, polyphyllin I inhibited tumor development in hepatocellular carcinoma and reduced the expression of phosphorylation of NF-κB and the downstream targets of NF-κB (16). Polyphyllin I reduced the secretion of IL-8 to inhibit keratinocyte cell proliferation and migration,17 and also provided protection against the left anterior descending coronary artery ligation-induced myocardial cell apoptosis, and inflammation through the inhibition of NF-κB signaling.18 However, the role of polyphyllin I in acute lung injury-associated inflammation has not been reported.
In this study, the effects of polyphyllin I on inflammation, oxidative stress, and pyroptosis of lipopolysaccharide--treated bronchial epithelial and pulmonary artery endothelial cells were investigated.
Materials and Methods
Cell culture and treatment
BEAS-2B and HPAEC cells were purchased from ATCC (Manassas, VA, USA). The cells were grown in DMEM (Sigma-Aldrich, St. Louis, MO, USA) with 10% fetal bovine serum (Gibco, Carlsbad, CA, USA) at a 37°C incubator. To induce the injury, cells were incubated with increasing concentrations of lipopolysaccharide (Sigma-Aldrich) at 2, 5, and 10 μg/mL for 12 hours. To investigate the protective role of polyphyllin I, the cells were treated with 1, 2, 4, 6, 8, or 10 μM polyphyllin I (ChemFaces Biochemical Co., Ltd, Chengdu, China) for 24 hours. The cells were also treated with 5 μg/mL lipopolysaccharide for 12 hours, and then incubated with 0.5, 1, or 2 μM polyphyllin I for another 24 hours.
Cell transfection and viability assays
The BEAS-2B cells were transfected with pcDNA-p65 or the negative control (Invitrogen, Carlsbad, CA, USA) using Lipofectamine 2000 (Invitrogen) for 24 hours, and then incubated with 5 μg/mL lipopolysaccharide and 1 μM polyphyllin I. The BEAS-2B and HPAEC post indicated treatment were incubated with MTT solution (Beyotime, Beijing, China) for 4 hours. Dimethyl sulfoxide was added, and the absorbance at 450 nm was measured by microplate reader (Bio-Rad, Hercules, CA, USA).
ELISA
BEAS-2B and HPAEC cells were lysed in RIPA buffer (Beyotime), and the supernatants were collected and detected by BCA kit (Applygen, Beijing, China) to determine the protein concentration. The levels of SOD, MDA, GSH, and MPO were evaluated using ELISA kits (ExCell Biology, Shanghai, China). The cultured medium of BEAS-2B and HPAEC were also harvested, and the levels of TNF-α, IL-6, and IL-1β were also determined by the ELISA kits (ExCell Biology).
qRT-PCR
BEAS-2B was lysed using TRIzol kit (Invitrogen) to isolate the RNAs. The RNAs were then synthesized into cDNAs using MultiscribeTM Reverse transcription Kit (Applied Biosystems, CA, USA). The mRNA expression of caspase-1 was detected by PreTaq II kit (Takara, Dalian, Liaoning, China) with following primers: caspase-1 (Forward: 5’-GCCTGTTCCTGTGATGTGGAG-3’ and Reverse: 5’-TGCCCACAGACATTCATACAGTTTC-3’) and GAPDH (Forward: 5’-GGCATGGACTGTGGTCATGAG-3’ and Reverse: 5’-TGCACCCCAACTGCTTAGC-3’). The mRNA expression was normalized to GAPDH through 2-∆∆Ct method.
Immunofluorescence
BEAS-2B cells were fixed in 4% paraformaldehyde, and permeabilized with 0.2% Triton X-100. The cells were then treated with 5% bovine serum albumin, and incubated with rabbit anti-human GSDMD antibody (1:100; Abcam, Cambridge, MA, USA). The cells were then incubated with horseradish peroxidase-conjugated goat anti--rabbit secondary antibody, and then stained with diaminobenzidine. The cells were also counterstained with DAPI, and observed under inverted fluorescence microscope (Olympus Corporation, Tokyo, Japan).
Western blot
The proteins isolated from BEAS-2B and HPAEC cells were separated by 10% SDS-PAGE, and transferred onto nitrocellulose membranes. The membranes were blocked in 5% bovine serum albumin, and then probed with specific antibodies: anti-GSDMD and anti-GSDMD-N (1:1500), anti-pro caspase-1 and anti-cleaved caspase-1 (1:2000), anti-β-actin (1:2500), anti-IκBα and anti-p-IκBα (1:3000), anti-p-p65, and anti-p65 (1:3500). The membranes were probed with horseradish peroxidase-conjugated secondary antibody (1:4000), and the immunoreactivities were determined using enhanced chemiluminescence (Sigma-Aldrich), and the protein signals were detected with β-actin as a reference using Image J software. All the antibodies were acquired from Abcam.
Statistical analysis
All the data with at least triple replicates were expressed as mean ± SD, and analyzed by one-way analysis of variance under SPSS software, followed by post hoc analysis. The normality and homogeneity of the data were analyzed by Shapiro-Wilk and Levene test respectively. A p value of ˂ 0.05 was considered as statistically significant.
Results
Polyphyllin I enhanced the cell viability of -lipopolysaccharide-treated BEAS-2B/HPAEC
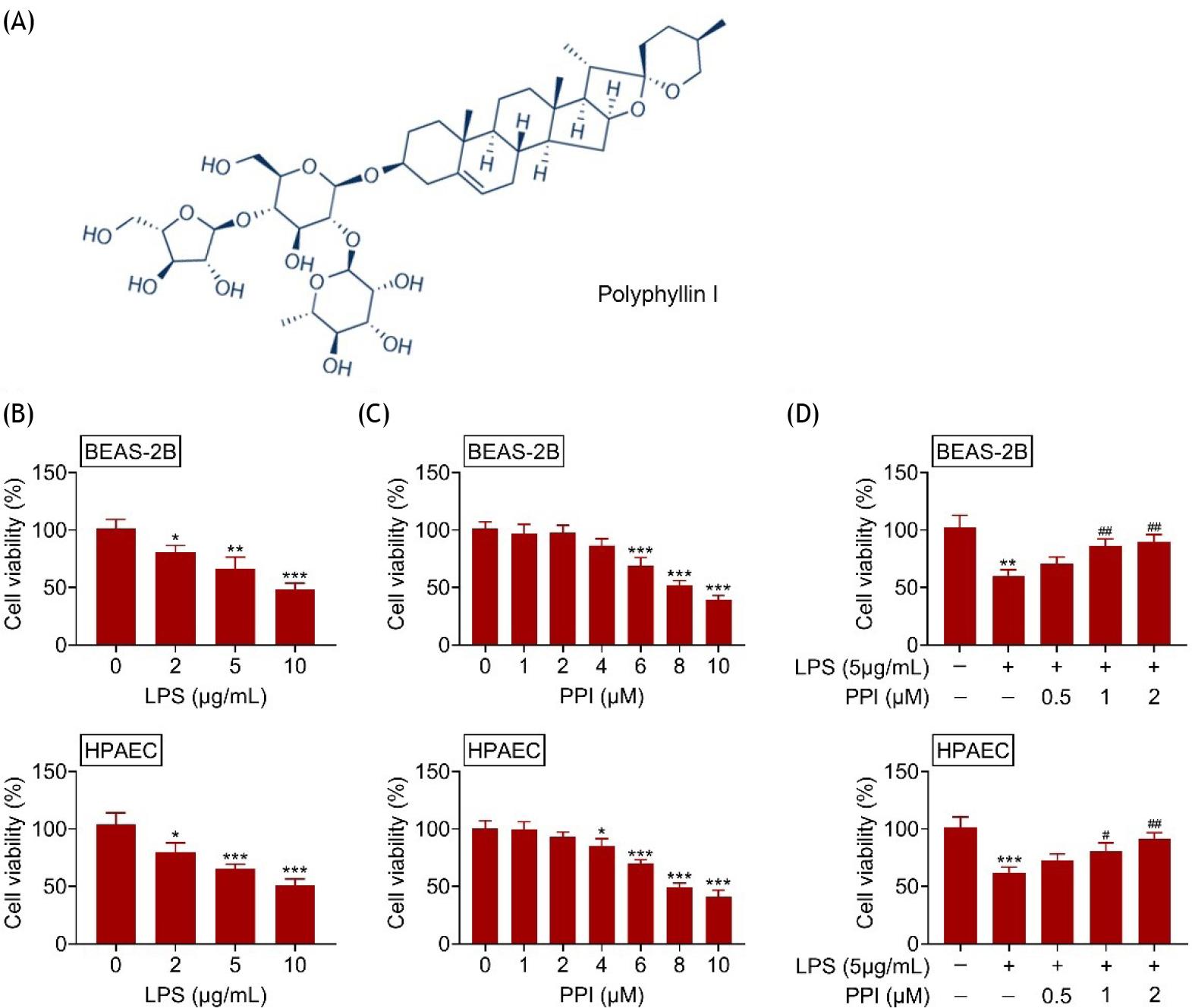
To investigate the role of polyphyllin I (Figure 1A) in acute lung injury, BEAS-2B and HPAEC cells were first stimulated with lipopolysaccharide. Incubation with lipopolysaccharides reduced the cell viabilities of BEAS-2B and HPAEC in a dosage dependent way (Figure 1B). BEAS-2B and HPAEC cells were also treated with polyphyllin I to detect the cytotoxicity. The results showed that polyphyllin I below 2 μM did not affect the cell viabilities of BEAS-2B and HPAEC (Figure 1C). However, polyphyllin I more than 4 μM significantly reduced the cell viabilities of BEAS-2B and HPAEC (Figure 1C). BEAS-2B and HPAEC cells were treated with 5 μg/mL lipopolysaccharide, and then incubated with 0.5, 1, or 2 μM polyphyllin I. Polyphyllin I increased cell viabilities of lipopolysaccharide-treated BEAS-2B/HPAEC (Figure 1D), thus providing protection against lipopolysaccharide--induced cytotoxicity in acute lung injury.
Figure 1 Polyphyllin I enhanced cell viability of lipopolysaccharide-treated BEAS-2B/HPAEC. (A) Chemical structure of polyphyllin I; (B) Incubation with lipopolysaccharide reduced cell viabilities of BEAS-2B and HPAEC in a dosage dependent way; (C) Polyphyllin I below 2 μM did not affect the cell viabilities of BEAS-2B and HPAEC, polyphyllin I more than 4 μM significantly reduced the cell viabilities of BEAS-2B and HPAEC.; (D) Polyphyllin I increased cell viabilities of lipopolysaccharide-treated BEAS-2B/HPAEC. *, **, *** vs. BEAS-2B/HPAEC without lipopolysaccharide and polyphyllin I treatment, p ˂ 0.05, p ˂ 0.01, p ˂ 0.001. #, ## vs. BEAS-2B/HPAEC with lipopolysaccharide treatment, p ˂ 0.05, p ˂ 0.01.
Polyphyllin I reduced the oxidative stress of lipopolysaccharide-treated BEAS-2B/HPAEC cells
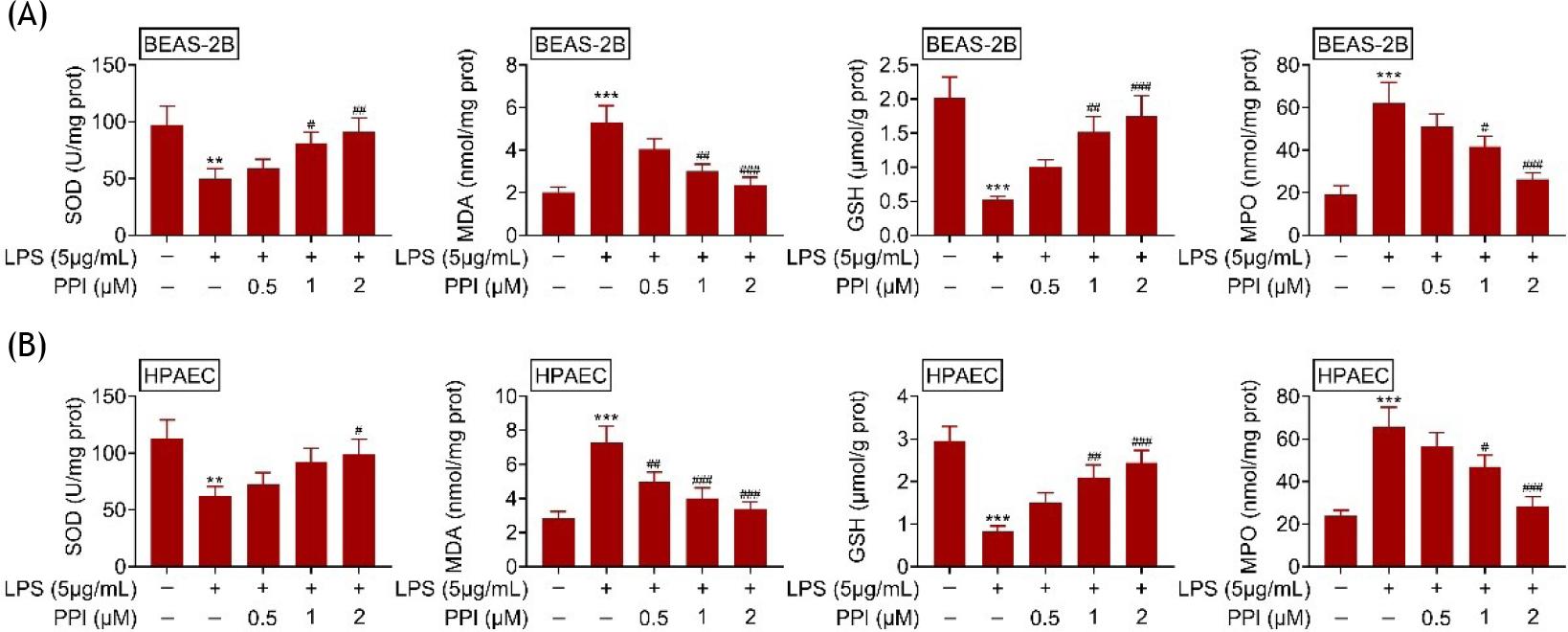
The levels of SOD and GSH in BEAS-2B (Figure 2A) and HPAEC (Figure 2B) were down-regulated by lipopolysaccharides. However, MDA and MPO were up-regulated in lipopolysaccharide-treated BEAS-2B/HPAEC cells (Figure 2A and B). Polyphyllin I increased the SOD and GSH levels, and decreased MDA and MPO in lipopolysaccharide-treated BEAS-2B/HPAEC cells (Figure 2A and B), demonstrating anti-oxidant property against acute lung injury.
Figure 2 Polyphyllin I reduced oxidative stress of lipopolysaccharide-treated BEAS-2B/HPAEC; (A) Polyphyllin I increased SOD and GSH, while decreased MDA and MPO in lipopolysaccharide-treated BEAS-2B; (B) Polyphyllin I increased SOD and GSH, while decreased MDA and MPO in lipopolysaccharide-treated HPAEC. **, *** vs. BEAS-2B/HPAEC without lipopolysaccharide and polyphyllin I treatment, p ˂ 0.01, p ˂ 0.001. #, ##, ### vs. BEAS-2B/HPAEC with lipopolysaccharide treatment, p ˂ 0.05, p ˂ 0.01, p ˂ 0.001
Polyphyllin I reduced the inflammation of -lipopolysaccharide-treated BEAS-2B/HPAEC cells
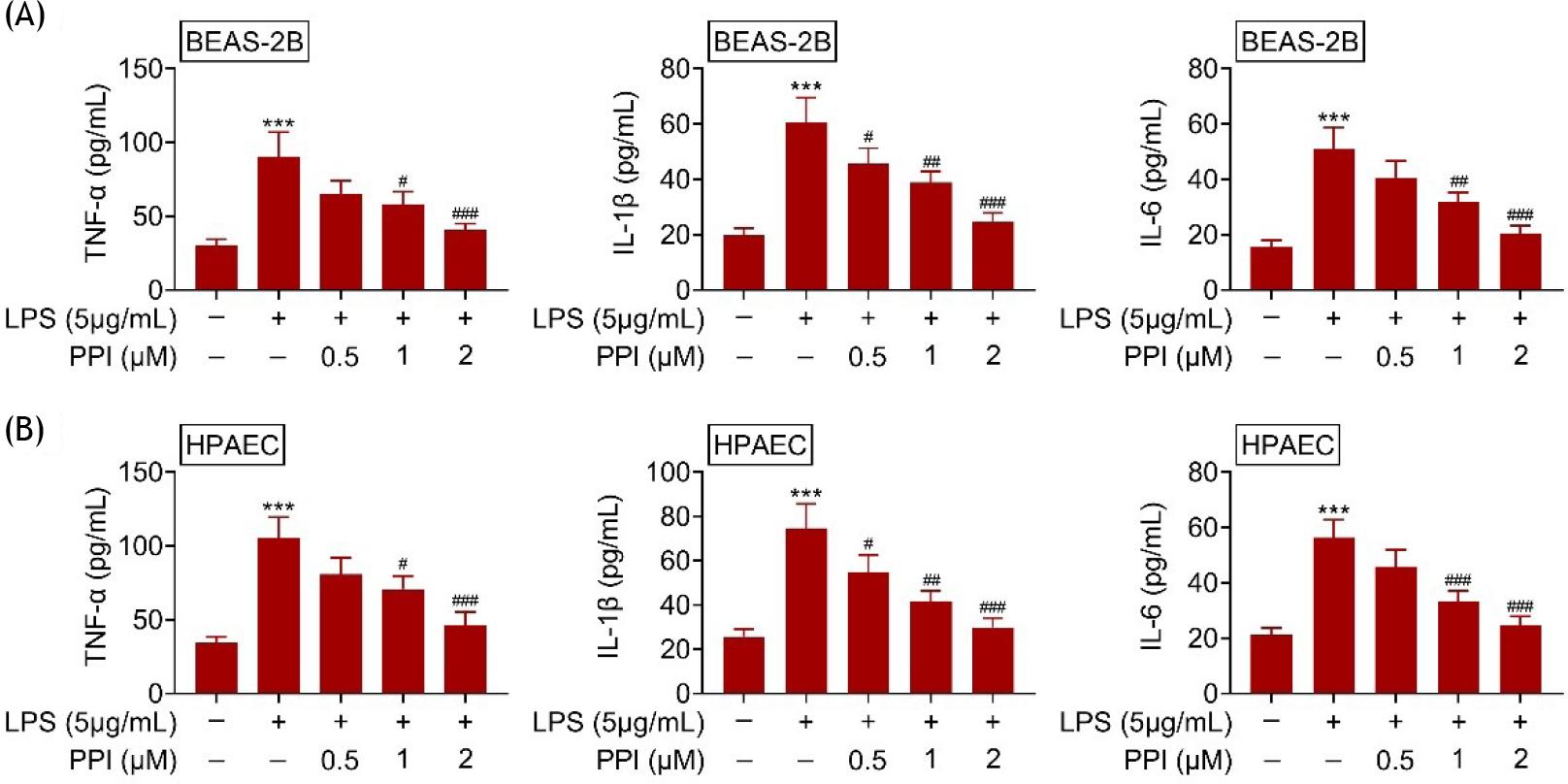
The levels of TNF-α, IL-1β, and IL-6 in BEAS-2B (Figure 3A) and HPAEC (Figure 3B) were up-regulated by lipopolysaccharides. However, polyphyllin I decreased the TNF-α, IL-1β, and IL-6 in lipopolysaccharide-treated BEAS-2B/HPAEC cells in a dose-dependent manner (Figure 3A and B), demonstrating the anti-inflammatory effects against acute lung injury.
Figure 3 Polyphyllin I reduced the inflammation of lipopolysaccharide-treated BEAS-2B/HPAEC; (A) Polyphyllin I decreased TNF-α, IL-1β, and IL-6 in lipopolysaccharide-treated BEAS-2B; (B) Polyphyllin I decreased TNF-α, IL-1β, and IL-6 in lipopolysaccharide-treated HPAEC. *** vs. BEAS-2B/HPAEC without lipopolysaccharide and polyphyllin I treatment, p ˂ 0.001. #, ##, ### vs. BEAS-2B/HPAEC with lipopolysaccharide treatment, p ˂ 0.05, p ˂ 0.01, p ˂ 0.001
Polyphyllin I reduced pyroptosis of lipopolysaccharide-treated BEAS-2B
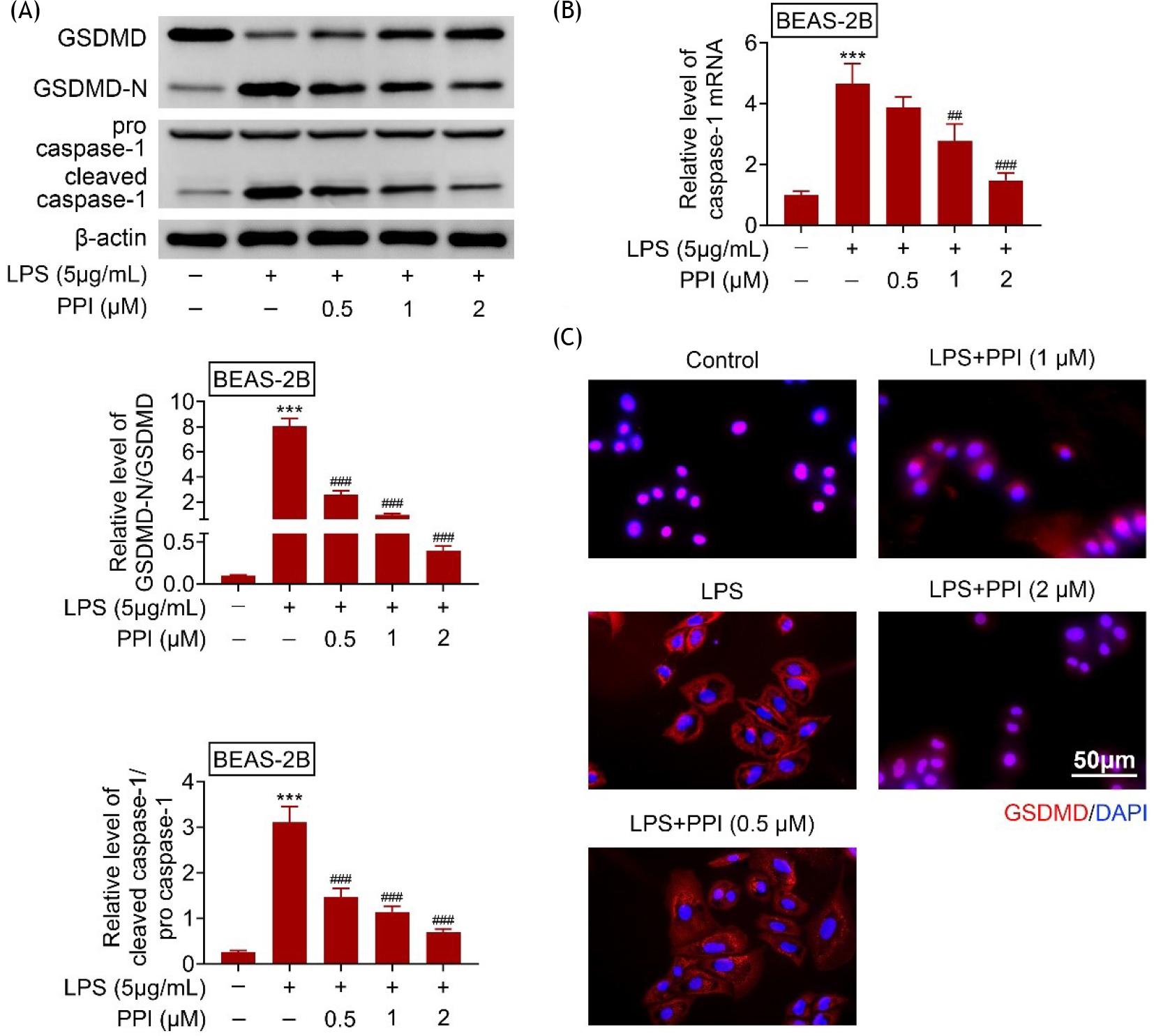
Incubation with lipopolysaccharides reduced the protein expression of GSDMD, enhanced GSDMD-N, pro caspase-1, and cleaved caspase-1 in BEAS-2B (Figure 4A). Polyphyllin I increased GSDMD, and decreased GSDMD-N, pro caspase-1 and cleaved caspase-1 in lipopolysaccharide-treated BEAS-2B (Figure 4A). Moreover, polyphyllin I attenuated the lipopolysaccharide-induced increase in caspase-1 mRNA in BEAS-2B (Figure 4B). Lipopolysaccharides also induced the translocation of GSDMD from cytoplasm to plasma membrane (Figure 4C). However, polyphyllin I suppressed the translocation of GSDMD from cytoplasm to plasma membrane (Figure 4C), thus reducing the pyroptosis of lipopolysaccharide-treated BEAS-2B.
Figure 4 Polyphyllin I reduced pyroptosis of lipopolysaccharide-treated BEAS-2B; (A) Polyphyllin I increased GSDMD, while decreased GSDMD-N, pro caspase-1 and cleaved caspase-1 in lipopolysaccharide-treated BEAS-2B; (B) Polyphyllin I reduced caspase-1 mRNA in lipopolysaccharide-treated BEAS-2B; (C) Polyphyllin I suppressed the translocation of GSDMD from cytoplasm to plasma membrane. *** vs. BEAS-2B without lipopolysaccharide and polyphyllin I treatment, p ˂ 0.001. #, ##, ### vs. BEAS-2B with lipopolysaccharide treatment, p ˂ 0.05, p ˂ 0.01, p ˂ 0.001.
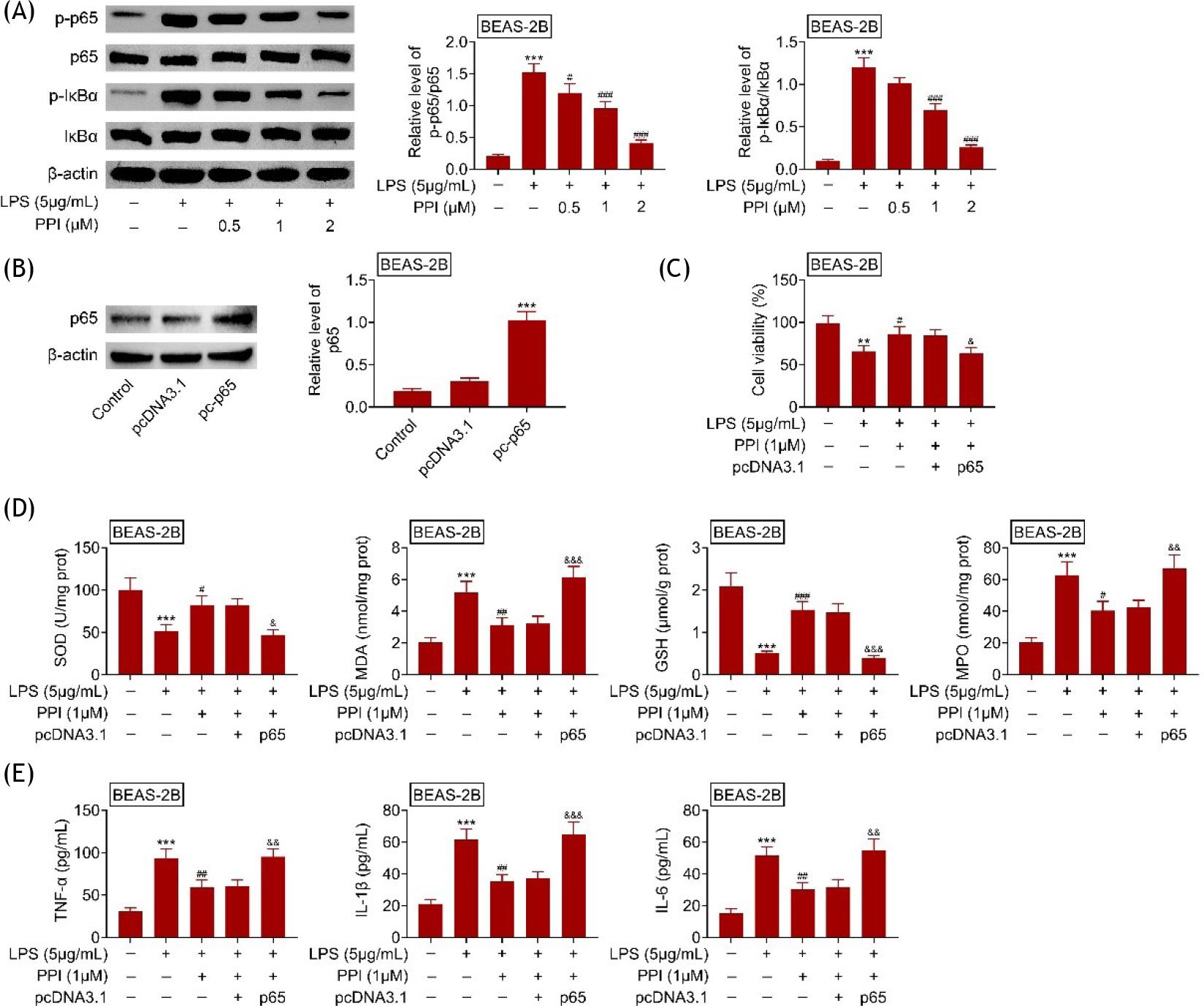
Polyphyllin I inhibited NF-κB in lipopolysaccharide-treated BEAS-2B
Incubation with lipopolysaccharides enhanced protein expression of p-IκBα and p-p65 in BEAS-2B (Figure 5A). However, polyphyllin I reduced the expression of p-IκBα and p-p65 in lipopolysaccharide-treated BEAS-2B (Figure 5A) to inhibit the activation of NF-κB signaling. BEAS-2B was transfected with pcDNA-p65 for the up-regulation of p65 (Figure 5B). The over-expression of p65 attenuated the polyphyllin I-induced decrease in cell viability in lipopolysaccharide-treated BEAS-2B (Figure 5C). Moreover, the up-regulation of SOD and GSH, and down-regulation of MDA and MPO in lipopolysaccharide-treated BEAS-2B were reversed by the over-expression of p65 (Figure 5D). The over-expression of p65 counteracted with the suppressive effects of polyphyllin I on the expression of TNF-α, IL-1β, and IL-6 in BEAS-2B (Figure 5E). These results indicated that Polyphyllin I inhibited the oxidative stress and inflammation of lipopolysaccharide-treated BEAS-2B through inactivation of NF-κB signaling.
Figure 5 Polyphyllin I inhibited NF-κB in lipopolysaccharide-treated BEAS-2B; (A) Polyphyllin I reduced the expression of p-IκBα and p-p65 in lipopolysaccharide-treated BEAS-2B; (B) BEAS-2B was transfected with pcDNA-p65 for the up-regulation of p65; (C) Over-expression of p65 attenuated polyphyllin I-induced decrease of cell viability in lipopolysaccharide-treated BEAS-2B; (D) Over-expression of p65 attenuated polyphyllin I-induced increase of SOD and GSH, decrease of MDA and MPO in lipopolysaccharide-treated BEAS-2B; (E) Over-expression of p65 attenuated polyphyllin I-induced decrease of TNF-α, IL-1β, and IL-6 in lipopolysaccharide-treated BEAS-2B. **, *** vs. BEAS-2B without lipopolysaccharide and polyphyllin I treatment, p ˂ 0.01, p ˂ 0.001. #, ## vs. BEAS-2B with lipopolysaccharide treatment, p ˂ 0.05, p ˂ 0.01. &, &&, &&& vs. BEAS-2B with lipopolysaccharide and polyphyllin I treatment, p ˂ 0.05, p ˂ 0.01, p ˂ 0.001
Discussion
This study for the first time revealed that polyphyllin I reduced the inflammation and oxidative stress in lipopolysaccharide-treated BEAS-2B/HPAEC cells, exhibiting anti-inflammatory and anti-oxidant effects against acute lung injury.
The lipopolysaccharide-treated bronchial epithelial cell was widely used as a model of acute lung injury.22 In this study, the lipopolysaccharide was also used to induce injury in BEAS-2B and HPAEC cells through decreasing cell viability and increasing inflammation. Polyphyllin I protected BEAS-2B and HPAEC cells against lipopolysaccharide-induced cytotoxicity by increasing the cell viability. Lipopolysaccharide has been shown to bind with toll-like receptor 4, resulting in the production of inflammatory mediators such as TNF-α, IL-1β, and IL-6, thus contributing to the lung tissue damage during the development of acute lung injury.23 The suppression of lipopolysaccharide-induced inflammation attenuated the acute lung injury.23 Polyphyllin I reduced the secretion of IL-1β, TNF-α, IL-6, and iNOS in lipopolysaccharide/IFN-γ activated macrophages.21 Here, polyphyllin I also decreased the levels of TNF-α, IL-1β, and IL-6 in lipopolysaccharide-treated BEAS-2B/HPAEC cells, indicating the anti-inflammatory property against acute lung injury.
A previous study has shown that lipopolysaccharide stimulates the activation of residential macrophages and recruited neutrophils, promoting the accumulation of reactive oxygen species in lung tissues, and induces extensive tissue damage and destruction, leading to pulmonary fibrosis.24 Therefore, the oxidative stress has implicated in the pathogenesis of acute lung injury.25 Polyphyllin I enhanced the levels of antioxidant, SOD, and GSH, and reduced the levels of oxidant, ROS, and MDA, to attenuate myocardial ischemia/reperfusion injury in rats.18 Here, polyphyllin I increased the SOD and GSH, and decreased MDA and MPO in lipopolysaccharide-treated BEAS-2B/HPAEC cells to suppress the oxidative stress in acute lung injury.
Pyroptosis is induced by inflammatory caspases such as caspase-1, 4, 5, and 11, and it promotes the secretion of inflammatory cytokines and leads to programmed cell death.26 Pyroptosis and the associated inflammatory caspases have been reported to be involved in the development of acute lung injury.26 Lipopolysaccharide activates caspase-1, cleaves GSDMD, and induces the translocation of GSDMD from cytoplasm to plasma membrane to promote the release of cytokines, such as IL-1β and IL-18, thus leading to cell death and pyroptosis of alveolar macrophages and endothelial cells.26,27 Inhibition of lipopolysaccharide-induced pyroptosis ameliorated the acute lung injury.28 Our results showed that Polyphyllin I enhanced the level of GSDMD, reduced the levels of caspase-1 and GSDMD-N in lipopolysaccharide-induced BEAS-2B. Moreover, polyphyllin I also suppressed the translocation of GSDMD from cytoplasm to plasma membrane, thus inhibiting pyroptosis to protect against acute lung injury.
NF-κB signaling which is essential for the secretion of pro-inflammatory factors, is activated in lipopolysaccharide--induced acute lung injury.29 the interference of NF-κB has been used for the treatment of acute lung injury.30 Polyphyllin I has been shown to reduce the phosphorylation activity of NF-κB.18-20 The over-expression of p65 eliminated the inhibitory effect of polyphyllin I on collagen-induced arthritis.21 Here, polyphyllin I reduced the expression of p-IκBα and p-p65 in lipopolysaccharide-treated BEAS-2B. Moreover, the over-expression of p65 attenuated polyphyllin I-induced decrease in cell viability, and increase in oxidative stress and inflammation in lipopolysaccharide-treated BEAS-2B, revealing that polyphyllin I inhibited the activation of NF-κB signaling protection against acute lung injury.
In conclusion, polyphyllin I reduced pyroptosis of lipopolysaccharide-treated BEAS-2B, and exerted anti-oxidant and anti-inflammatory effects against lipopolysaccharide--treated BEAS-2B/HPAEC through the inactivation of NF-κB signaling. Therefore, polyphyllin I might be a promising strategy for the prevention of acute lung injury and the associated complications. However, the effect of polyphyllin I on acute lung injury in vivo animal model should be investigated in further research.
Acknowledgements
Not applicable.
Funding
Not applicable.
Availability of Data and Materials
All data generated or analyzed during this study are included in this published article.
Competing Interests
The authors state that there are no conflicts of interest to disclose.
Ethics Approval
Not applicable.
Contribution of Authors
Fangli Mao designed the study and supervised the data collection, Aiping Wu analyzed the data and interpreted the data, Fangli Mao and Aiping Wu prepared the manuscript for publication and reviewed the draft of the manuscript. All authors have read and approved the manuscript.
REFERENCES
1. Ragaller M, Richter T. Acute lung injury and acute respiratory distress syndrome. J Emerg Trauma Shock. 2010;3(1):43–51. 10.4103/0974-2700.58663
2. Hotchkiss RS, Colston E, Yende S, Angus DC, Moldawer LL, Crouser ED, et al. Immune checkpoint inhibition in sepsis: a Phase 1b randomized, placebo-controlled, single ascending dose study of anti-PD-L1 (BMS-936559). Critical care medicine. 2019;47(5):632. 10.1097/CCM.0000000000003685
3. Sazonov V, Abylkassov R, Tobylbayeva Z, Saparov A, Mironova O, Poddighe D. Case Series: Efficacy and Safety of Hemoadsorption With HA-330 Adsorber in Septic Pediatric Patients With Cancer. Frontiers in Pediatrics. 2021;9. 10.3389/fped.2021.672260
4. Kojicic M, Li G, Hanson AC, Lee K-M, Thakur L, Vedre J, et al. Risk factors for the development of acute lung injury in patients with infectious pneumonia. Critical Care. 2012;16(2):R46. 10.1186/cc11247
5. Herrero R, Sanchez G, Lorente JA. New insights into the mechanisms of pulmonary edema in acute lung injury. Ann Transl Med. 2018;6(2):32. 10.21037/atm.2017.12.18
6. Hai Zhong LH, Xiang Li, Chunjing Wang, Xu Wu. Anti-inflammatory Role of Trilobatin on Lipopolysaccharide-induced Acute Lung Injury through Activation of AMPK/GSK3β-Nrf2 Pathway. Signa Vitae. 2020;16(2):160–6.
7. Metz C, Sibbald WJ. Anti-inflammatory Therapy for Acute Lung Injury: A Review of Animal and Clinical Studies. CHEST. 1991;100(4):1110–9. 10.1378/chest.100.4.1110
8. Brooks D, Barr LC, Wiscombe S, McAuley DF, Simpson AJ, Rostron AJ. Human lipopolysaccharide models provide mechanistic and therapeutic insights into systemic and pulmonary inflammation. European Respiratory Journal. 2020;56(1):1901298. 10.1183/13993003.01298-2019
9. Rittirsch D, Flierl MA, Day DE, Nadeau BA, McGuire SR, Hoesel LM, et al. Acute lung injury induced by lipopolysaccharide is independent of complement activation. J Immunol. 2008;180(11):7664–72. 10.4049/jimmunol.180.11.7664
10. Liu T-Y, Zhao L-L, Chen S-B, Hou B-C, Huang J, Hong X, et al. Polygonatum sibiricum polysaccharides prevent LPS-induced acute lung injury by inhibiting inflammation via the TLR4/Myd88/NF-κB pathway. Exp Ther Med. 2020;20(4):3733–9. 10.3892/etm.2020.9097
11. Zhang Y, He H, Zhang B, Chen Q, Yao S, Gui P. Amelioration of Lipopolysaccharide-Induced Acute Lung Injury in Rats by Na-H Exchanger-1 Inhibitor Amiloride Is Associated with Reversal of ERK Mitogen-Activated Protein Kinase. BioMed Research International. 2018;2018:3560234. 10.1155/2018/3560234
12. Rahmati M, Keshvari M, Mirnasouri R, Chehelcheraghi F. Exercise and Urtica dioica extract ameliorate hippocampal insulin signaling, oxidative stress, neuroinflammation, and cognitive function in STZ-induced diabetic rats. Biomedicine & Pharmacotherapy. 2021;139:111577. 10.1016/j.biopha.2021.111577
13. Rodriguez-Ramiro I, Vauzour D, Minihane A. Polyphenols and non-alcoholic fatty liver disease: impact and mechanisms. Proceedings of the Nutrition Society. 2016;75(1):47–60. 10.1017/S0029665115004218
14. Liu X, Sun Z, Deng J, Liu J, Ma K, Si Y, et al. Polyphyllin I inhibits invasion and epithelial-mesenchymal transition via CIP2A/PP2A/ERK signaling in prostate cancer. Int J Oncol. 2018;53(3):1279–88. 10.3892/ijo.2018.4464
15. Tian Y, Gong G-Y, Ma L-L, Wang Z-Q, Song D, Fang M-Y. Anti-cancer effects of Polyphyllin I: An update in 5 years. Chem Biol Interact. 2020;316:108936. 10.1016/j.cbi.2019.108936
16. Han W, Hou G, Liu L. Polyphyllin I (PPI) increased the sensitivity of hepatocellular carcinoma HepG2 cells to chemotherapy. Int J Clin Exp Med. 2015;8(11):20664–9.
17. Yang S, Jiang Y, Yu X, Zhu L, Wang L, Mao J, et al. Polyphyllin I Inhibits Propionibacterium acnes-Induced IL-8 Secretion in HaCaT Cells by Downregulating the CD36/NOX1/ROS/NLRP3/IL-1β Pathway. Evidence-Based Complementary and Alternative Medicine. 2021;2021:1–11. 10.1155/2021/8455709
18. Huang R, Shu J, Dai X, Liu Y, Yu F, Shi G. The protective effect of polyphyllin I on myocardial ischemia/reperfusion injury in rats. Ann Transl Med. 2020;8(10):644.10.21037/atm-20-3371
19. Negi J, Bisht V, Bhandari A, Vp B, Negi P, Singh N. Paris polyphylla: Chemical and Biological Prospectives. Anti-cancer agents in medicinal chemistry. 2014;14:833–9. 10.2174/1871520614666140611101040
20. Zhu T, Wu W, Yang S, Li D, Sun D, He L. Polyphyllin I Inhibits Propionibacterium acnes-Induced Inflammation In Vitro. Inflammation. 2019;42(1):35–44. 10.1007/s10753-018-0870-z
21. Wang Q, Zhou X, Zhao Y, Xiao J, Lu Y, Shi Q, et al. Polyphyllin I Ameliorates Collagen-Induced Arthritis by Suppressing the Inflammation Response in Macrophages Through the NF-κB Pathway. Front Immunol. 2018;9:2091.10.3389/fimmu.2018.02091
22. Liu P, Feng Y, Li H, Chen X, Wang G, Xu S, et al. Ferrostatin-1 alleviates lipopolysaccharide-induced acute lung injury via inhibiting ferroptosis. Cell Mol Biol Lett. 2020;25:10. 10.1186/s11658-020-00205-0
23. Huang C-Y, Deng J-S, Huang W-C, Jiang W-P, Huang G-J. Attenuation of Lipopolysaccharide-Induced Acute Lung Injury by Hispolon in Mice, Through Regulating the TLR4/PI3K/Akt/mTOR and Keap1/Nrf2/HO-1 Pathways, and Suppressing Oxidative Stress-Mediated ER Stress-Induced Apoptosis and Autophagy. Nutrients. 2020;12(6):1742. 10.3390/nu12061742
24. Ward PA. Oxidative stress: acute and progressive lung injury. Annals of the New York Academy of Sciences. 2010;1203(1):53. 10.1111/j.1749-6632.2010.05552.x
25. Chow C-W, Herrera Abreu MT, Suzuki T, Downey G. Oxidative Stress and Acute Lung Injury. American journal of respiratory cell and molecular biology. 2003;29:427–31. 10.1165/rcmb.F278
26. Liu B, He R, Zhang L, Hao B, Jiang W, Wang W, et al. Inflammatory Caspases Drive Pyroptosis in Acute Lung Injury. Frontiers in Pharmacology. 2021;12. 10.3389/fphar.2021.631256
27. Burdette BE, Esparza AN, Zhu H, Wang S. Gasdermin D in pyroptosis. Acta Pharm Sin B. 2021;11(9):2768–82. 10.1016/j.apsb.2021.02.006
28. Wu D, Pan P, Su X, Zhang L, Qin Q, Tan H, et al. Interferon Regulatory Factor-1 Mediates Alveolar Macrophage Pyroptosis During LPS-Induced Acute Lung Injury in Mice. Shock. 2016;46(3):329–38. 10.1097/SHK.0000000000000595
29. Kan X, Chen Y, Huang B, Fu S, Guo W, Ran X, et al. Effect of Palrnatine on lipopolysaccharide-induced acute lung injury by inhibiting activation of the Akt/NF-κB pathway. J Zhejiang Univ Sci B. 2021;22(11):929–40. 10.1631/jzus.B2000583
30. Li N, Song Y, Zhao W, Han T, Lin S, Ramirez O, et al. Small interfering RNA targeting NF-κB attenuates lipopolysaccharide-induced acute lung injury in rats. BMC Physiol. 2016;16(1):7. 10.1186/s12899-016-0027-y