
Download
ORIGINAL RESEARCH
PTPRO activates TLR4/NF-κB signaling to intensify lipopolysaccharide-induced pneumonia cell injury
Yao Chena, Buming Sunb*
aDepartment of Pediatric, The First Affiliated Hospital of Chengdu Medical College, Chengdu, China
bDepartment of Pediatric, Huai’an Second People’s Hospital, Huai’an, China
Abstract
Background Protein tyrosine phosphatase receptor type O (PTPRO) belongs to the PTP (protein tyrosine phosphatase) family and is widely expressed in parenchymal cells, such as breast and lung epithelial cells. PTPRO has been shown to enhance inflammatory responses and has been implicated in the pathogenesis of inflammation-associated diseases. The role of PTPRO in pneumonia was investigated.
Methods Human embryonic lung fibroblasts (HFL1) were treated with increasing concentrations of lipopolysaccharide at 5, 10, or 20 μg/mL to induce inflammatory and apoptotic injuries. Expression of PTPRO was detected by western blot. Inflammation and apoptosis were assessed by ELISA and flow cytometry assays, respectively.
Results Lipopolysaccharide induced decreased cell viability, and increased inflammation and apoptosis in HFL1. PTPRO was upregulated in HFL1 post lipopolysaccharide treatment, and silencing of PTPRO enhanced lipopolysaccharide-induced cell viability of HFL1, and suppressed the inflammation and apoptosis. However, overexpression of PTPRO aggravated lipopolysaccharide-induced cytotoxicity in HFL1. Overexpression of PTPRO upregulated protein expression of TLR4 and p-p65 in lipopolysaccharide-induced HFL1, while knockdown of PTPRO reduced the level of p-IκBα to downregulate levels of TLR4 and p-p65.
Conclusion PTPRO contributed to pro-inflammatory and pro-apoptotic effects on lipopolysaccharide-induced HFL1 through activation of TLR4/NF-κB signaling.
Key words: PTPRO, apoptosis, inflammation, lipopolysaccharide, HFL1, TLR4/NF-κB, pneumonia
*Corresponding author: Buming Sun, Department of Pediatric, Huai’an Second People’s Hospital, No. 62 Huaihai South Road, Huai’an, Jiangsu Province, China. Email address: [email protected]
Received 20 January 2022; Accepted 25 February 2022; Available online 1 May 2022
Copyright: Chen Y and Sun B
License: This open access article is licensed under Creative Commons Attribution 4.0 International (CC BY 4.0). http://creativecommons.org/licenses/by/4.0/
Introduction
Pediatric pneumonia is one of the most common diseases in infants with increasing incidence and morbidity.1 Pediatric pneumonia with the main symptoms of dyspnea, fever, cough, and shortness of breath can lead to long duration and severe extra pulmonary complications in infants and young children.2 Bacterial and viral infections induce distal airway inflammation in infants, and result in alteration of pulmonary circulation, pulmonary cell damage, and interference of normal respiratory mechanism during the development of pediatric pneumonia.3 Antibiotic therapies are the first-line treatment for pediatric pneumonia. However, drug resistance limits the clinical use of the therapies.4 Therefore, novel strategies for the prevention of pediatric pneumonia are urgently needed.
Protein tyrosine phosphatase receptor type O (PTPRO) is a receptor-type PTP that is widely expressed in multiple organs, including breast, liver, lung, brain, and kidney.5 PTPRO with the conserved intracellular PTP domain catalyzes dephosphorylation of tyrosine peptides in target genes, thus participating in biological processes, such as apoptosis, differentiation, and proliferation.6 PTPRO was also found to be associated with immune infiltration in distinct cancers,7,8 and functioned as a tumor suppressor9 through polarization of macrophages into M1-like tumor--associated macrophages.10 Moreover, PTPRO also plays a role in inflammation-associated tissue damage. For example, knockout of PTPRO in mice reduced bile-duct ligation or carbon tetrachloride administration-induced liver fibrosis and tissue remodeling through downregulation of inflammatory factors.11 PTPRO promoted NF-κB activation and contributed to fulminant hepatitis.12 However, the role of PTPRO in pneumonia is unclear.
TLRs function as pattern-recognition receptors to recognize pathogen-associated molecular patterns and activated multiple downstream inflammatory pathways.13 In pneumonia, activation of TLR4/NF-κB signaling was essential for lipopolysaccharide-induced inflammatory and apoptotic injuries,14 and inhibition of TLR4/NF-κB signaling ameliorated pneumonia-associated injuries.15 Therefore, PTPRO might promote progression of pneumonia through activation of TLR4/NF-κB signaling. The effects of PTPRO on cell apoptosis and inflammation in lipopolysaccharide--induced HFL1 were investigated.
Materials and Methods
Cell culture and treatment
Human lung fibroblasts (HFL1) (ATCC, Manassas, VA, USA) were cultured in DMEM medium containing 10% fetal bovine serum (Life Technologies, Gaithersburg, MD, USA). HFL1 cells were treated with 5, 10, or 20 μg/mL lipopolysaccharide (Sigma-Aldrich, St. Louis, MO, USA) for 12 hours to induce injury.
Cell transfection
HFL1 cells were seeded in 96-well plates, and treated with 5 μg/mL lipopolysaccharide. Full length of PTPRO was subcloned into pcDNA vector (Thermo Fisher Scientific, Waltham, MA, USA). shRNA targeting PTPRO (shPTPRO) and the negative control (shNC) were acquired from Genepharma (Suzhou, China). Lipopolysaccharide-induced HFL1 cells were transfected with pcDNA vectors (300 μg) or shRNAs (50 nM) via Lipofectamine 2000 (Thermo Fisher Scientific).
Cell viability and apoptosis assays
Lipopolysaccharide-induced HFL1 with or without transfections were incubated with 5 mg/mL MTT (10 μL) (Beyotime, Beijing, China) for 4 hours. Microplate reader (Thermo Fisher Scientific) was used to detect absorbance at 490 nm. For cell apoptosis analysis, HFL1 cells were resuspended in binding buffer of Annexin Fitc Pi Staining Kit (Thermo Fisher Scientific), and then stained with 5 µL of PI and 5 μL of FITC-labeled annexin V. The apoptotic ratio was evaluated by FACS flow cytometer (Life Technologies).
ELISA
Culture supernatants of HFL1 were harvested, and the levels of TNF-α, IL-1β, and IL-18 were detected by ELISA kits (Pharmingen, San Diego, CA, USA).
qRT-PCR
RNAs were extracted from HFL1 via TRIzol reagent (Invitrogen, Carlsbad, CA, USA) and then synthesized into cDNAs. The mRNA expression of TNF-α, IL-1α, and IL-18 were analyzed under SYBR® premix Ex TaqTM kit (Takara, Dalian, China). The primers are shown in Table 1, and the expression levels were normalized to β-actin using the 2-ΔΔCt method.
Table 1 Primers.
| Genes | Forward | Reverse |
|---|---|---|
| β-actin (NG_007992.1) | 5'-TGTGATGGTGGGAATGGGTCAG-3' | 5'-TTTGATGTCACGCACGATTTCC-3' |
| IL-18 (NG_028143.1) | 5'-GGCATTTGCGAGCTGCCAAATTCC-3' | 5'-CTTGTTGTGTCCTGGAACACG-3' |
| IL-1β (NG_008851.1) | 5'-CCAGCTTCAAATCTCACAGCAG-3' | 5'-CTTCTTTGGGTATTGCTTGGGATC-3' |
| TNF-α (NG_007462.1) | 5'-CCGAGTGACAAGCCTGTAGC-3' | 5'-AGGAGGTTGACCTTGGTCTG-3' |
Western blot
HFL1 cells were lysed in RIPA buffer (Beyotime) and the isolated proteins were separated by 10% SDS-PAGE. Samples were transferred onto nitrocellulose membranes, and membranes were blocked in 5% bovine serum albumin. The membranes were probed with specific antibodies: anti-PTPRO (ab150834) and anti-β-actin (ab179467) (1:2000; Abcam, Cambridge, MA, USA); anti-p-IκBα (ab133462) and anti-IκBα (ab97783) (1:2500, Abcam); anti-p-p65 (ab28856) and anti-p65 (ab16502) (1:3500, Abcam); and anti-TLR4 (ab13556) and anti-GAPDH (ab9485) (1:4000, Abcam). The membranes were then washed and incubated with horseradish peroxidase-conjugated secondary antibody (ab205718 or ab97240) (1:4500, Abcam). Immunoreactivities were visualized using enhanced chemiluminescence (Sigma-Aldrich). Protein signals were quantified using β-actin and GAPDH as references.
Statistical analysis
All the data with at least triple replicates were expressed as mean ± SEM. Shapiro–Wilk and Levene’s tests were used to detect the normality and homogeneity of data. The data were analyzed by student’s t-test or one-way analysis of variance followed by Tukey’s post hoc analysis under SPSS software. A p-value of ˂ 0.05 was considered as statistically significant.
Results
Lipopolysaccharide induced elevation of PTPRO in HFL1
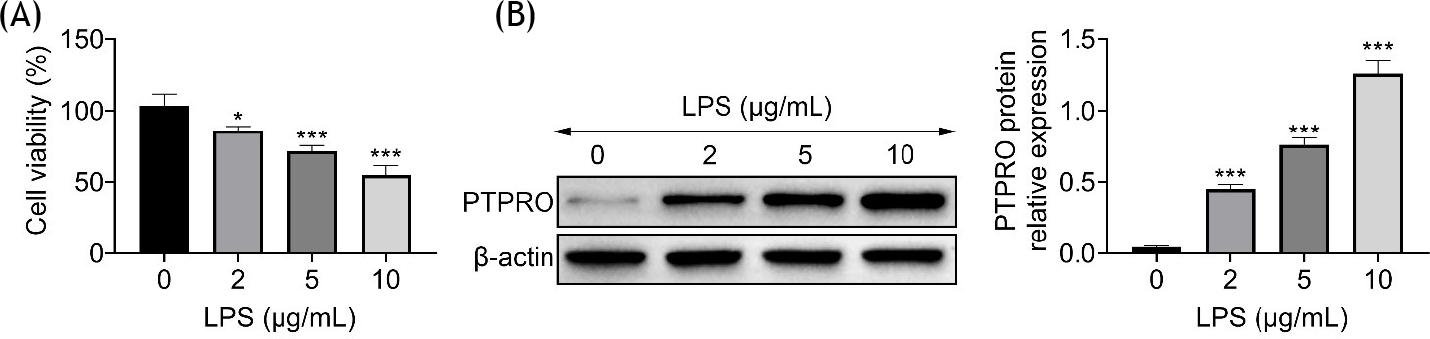
HFL1 was incubated with lipopolysaccharide to induce injury. Results showed that cell viability of HFL1 was decreased by lipopolysaccharide in a dose-dependent manner (Figure 1A). Protein expression of PTPRO was upregulated in HFL1 by lipopolysaccharide in a dose-dependent manner (Figure 1B), demonstrating that PTPRO might be involved in lipopolysaccharide-induced injury in HFL1.
Figure 1 Lipopolysaccharide induced the elevation of PTPRO in HFL1. (A) Incubation with lipopolysaccharide reduced cell viability of HFL1 in a dose-dependent way. (B) Incubation with lipopolysaccharide enhanced protein expression of PTPRO in HFL1 in a dose-dependent way. *p ˂ 0.05, ***p ˂ 0.001.
PTPRO contributed to lipopolysaccharide-induced apoptosis in HFL1
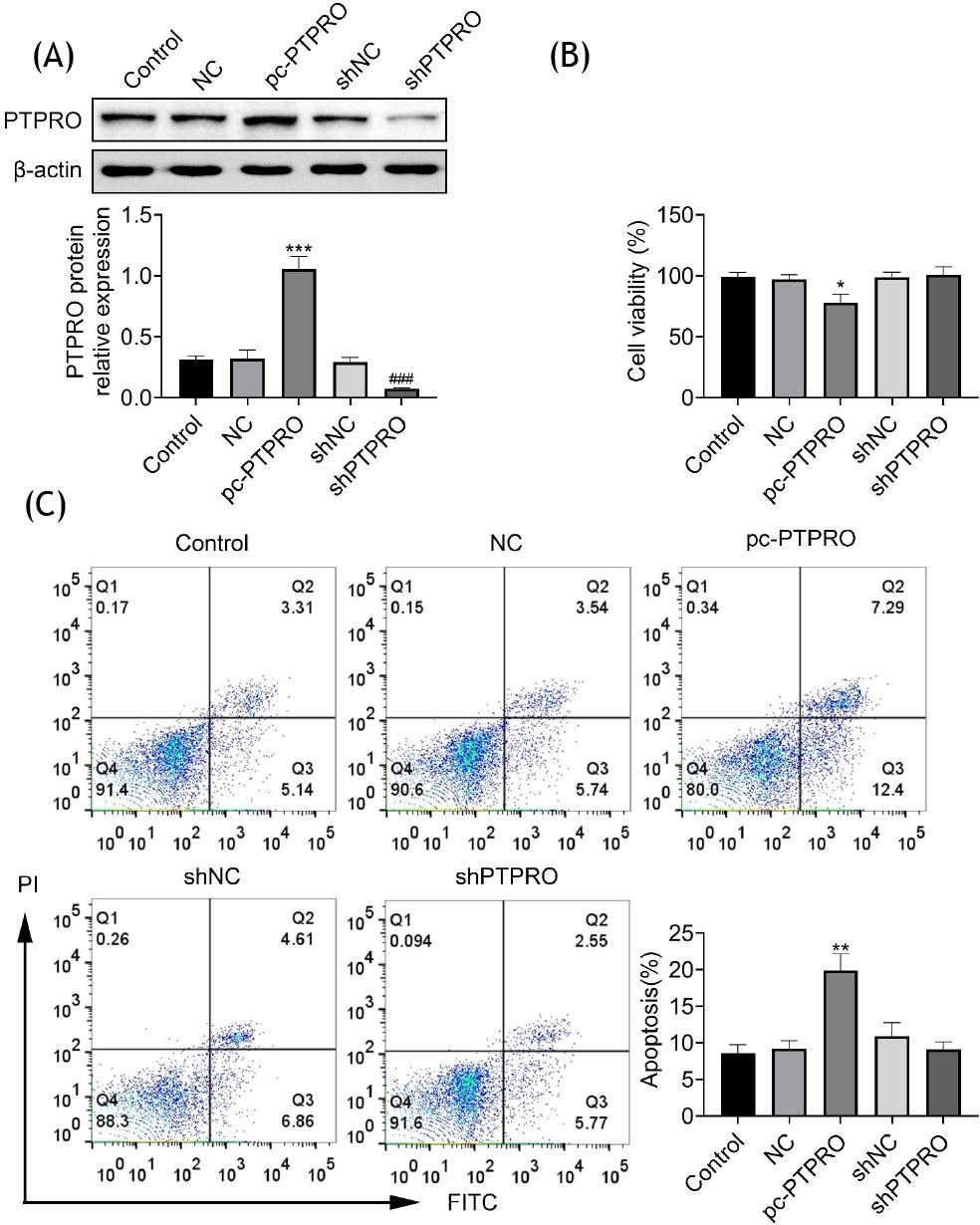
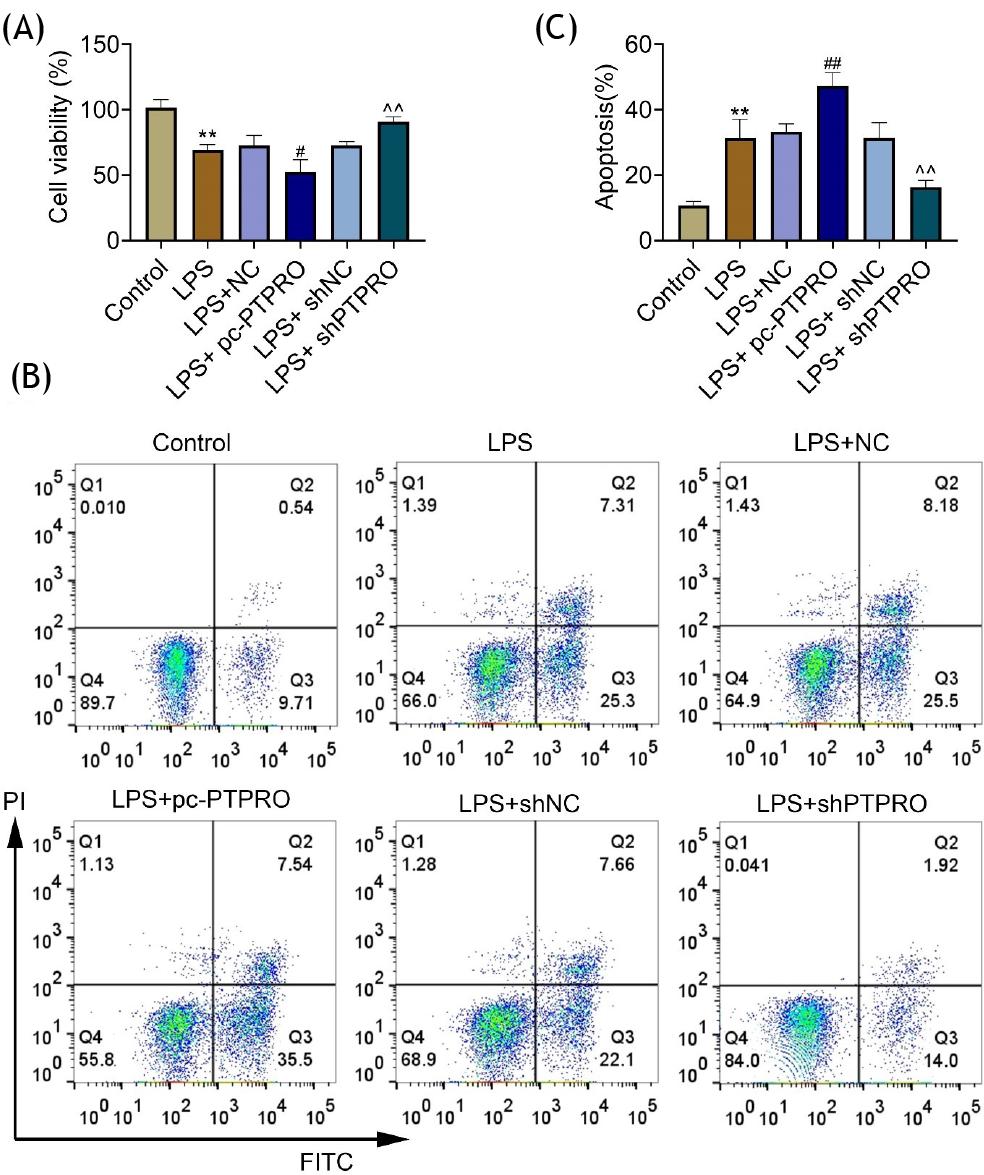
To investigate cytotoxicity of PTPRO, HFL1 cells were transfected with pcDNA-PTPRO or shPTPRO to increase or decrease PTPRO expression, respectively (Figure 2A). Overexpression of PTPRO reduced cell viability of HFL1 (Figure 2B) and promoted cell apoptosis (Figure 2C). However, cell viability (Figure 2B) and apoptosis (Figure 2C) were not affected by the silencing of PTPRO. Lipopolysaccharide-induced HFL1 cells were also transfected with pcDNA-PTPRO or shPTPRO. Transfection with pcDNA-PTPRO reduced the cell viability of lipopolysaccharide-induced HFL1, while transfection with shPTPRO enhanced cell viability (Figure 3A). Moreover, overexpression of PTPRO increased cell apoptosis of lipopolysaccharide-induced HFL1, while transfection with shPTPRO inhibited cell apoptosis (Figure 3B and C). These results showed the anti-apoptotic effect of PTPRO silencing against pneumonia.
Figure 2 Cytotoxicity of PTPRO in HFL1. (A) Transfection with pcDNA-PTPRO or shPTPRO increased or decreased PTPRO expression, respectively. (B) Overexpression of PTPRO reduced cell viability of HFL1, while silencing of PTPRO did not affect cell viability. (C) Overexpression of PTPRO promoted cell apoptosis of HFL1, while silencing of PTPRO did not affect cell apoptosis. *p ˂ 0.05, **p ˂ 0.01.
Figure 3 PTPRO contributed to lipopolysaccharide-induced apoptosis in HFL1. (A) Transfection with pcDNA-PTPRO reduced the cell viability of lipopolysaccharide-induced HFL1, while transfection with shPTPRO enhanced cell viability. (B) Overexpression of PTPRO increased cell apoptosis of lipopolysaccharide-induced HFL1, while transfection with shPTPRO inhibited cell apoptosis. (C) The apoptotic ratio of lipopolysaccharide-induced HFL1 that was transfected with pcDNA-PTPRO or shPTPRO. #p ˂ 0.05; ##,^^p ˂ 0.01; ***,###p ˂ 0.001.
PTPRO contributed to lipopolysaccharide-induced inflammation in HFL1
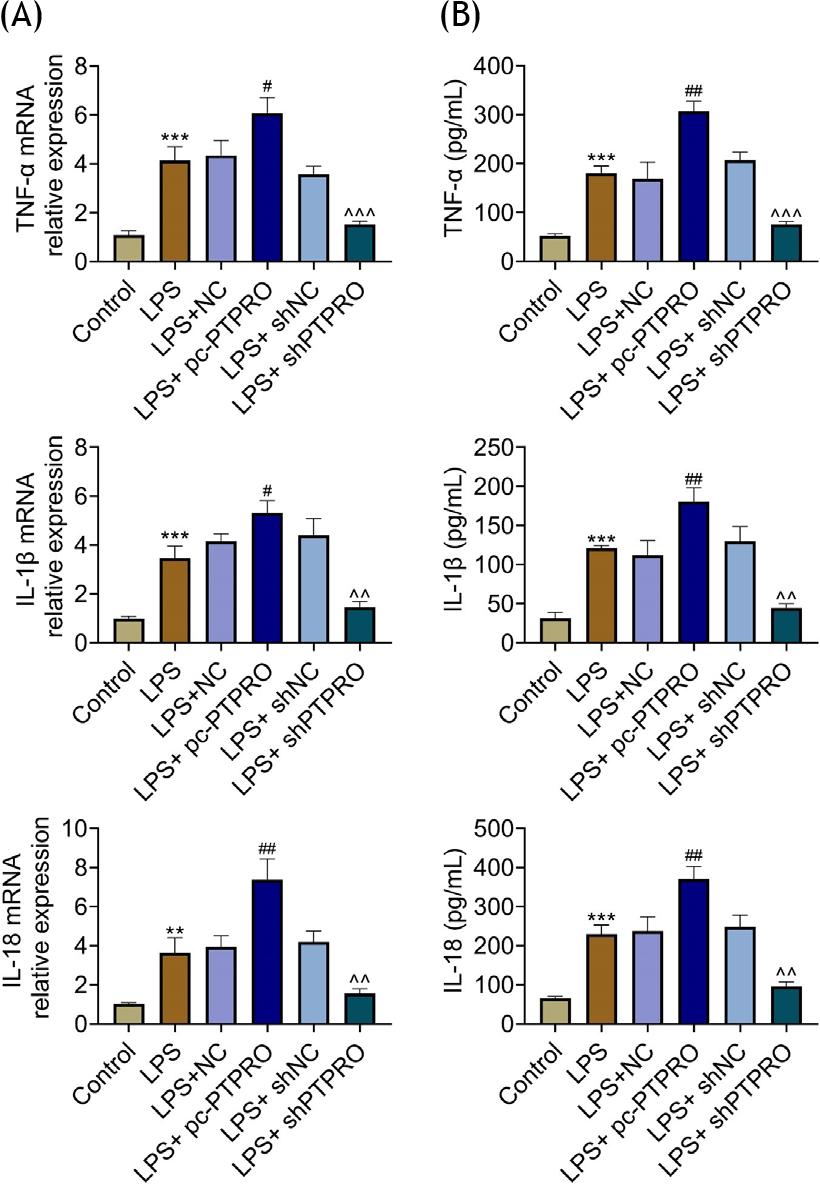
Lipopolysaccharide promoted the expression of TNF-α, IL-1α, and IL-18 in HFL1 (Figure 4A and B). The expressions of TNF-α, IL-1α, and IL-18 in lipopolysaccharide-induced HFL1 were enhanced by the overexpression of PTPRO, and reduced by PTPRO silencing (Figure 4A and B), revealing the anti--inflammatory effect of PTPRO silencing against pneumonia.
Figure 4 PTPRO contributed to lipopolysaccharide-induced inflammation in HFL1. (A) mRNA expressions of TNF-α, IL-1α, and IL-18 in lipopolysaccharide-induced HFL1 were enhanced by PTPRO overexpression, while they were reduced by PTPRO silence. (B) Protein expressions of TNF-α, IL-1α, and IL-18 in lipopolysaccharide-induced HFL1 were enhanced by PTPRO overexpression, while they were reduced by PTPRO silence. #p ˂ 0.05; **,##,^^p ˂ 0.01; ***,^^^p ˂ 0.001.
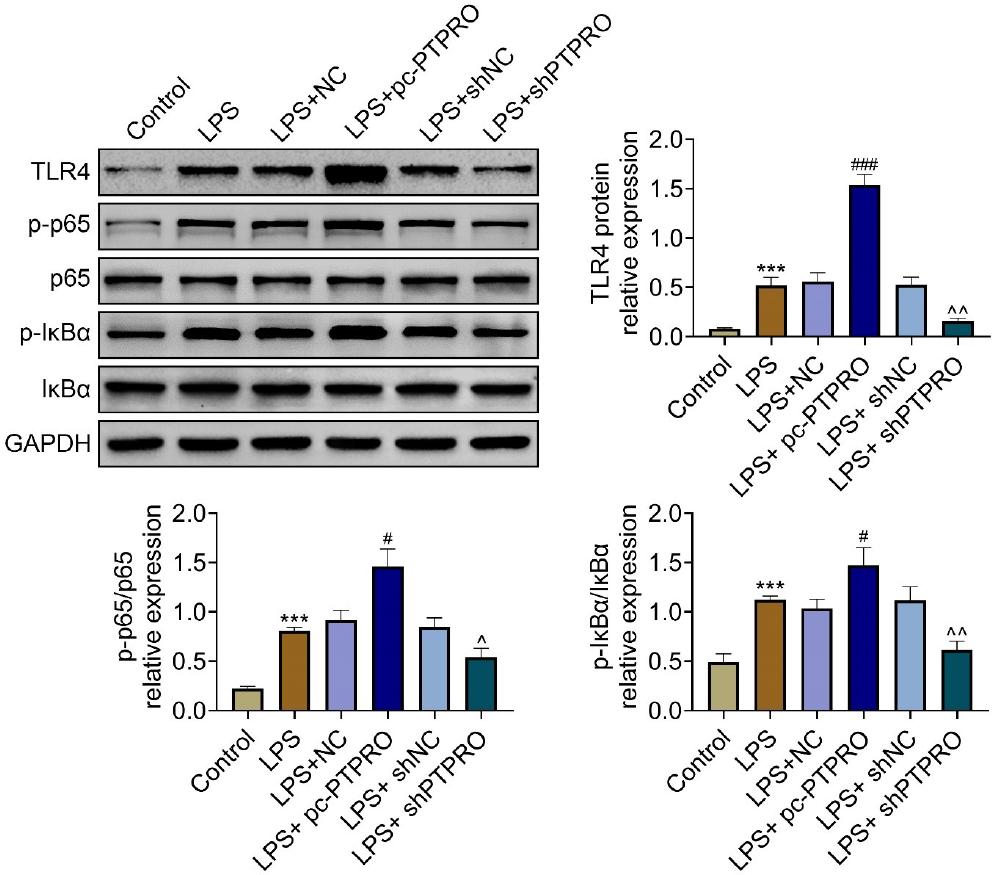
PTPRO contributed to lipopolysaccharide-induced activation of TLR4/NF-κB in HFL1
Protein expressions of TLR4 and p-p65 were upregulated in HFL1 by lipopolysaccharide (Figure 5). Lipopolysaccharide also induced upregulation of p-IκBα in HFL1 (Figure 5). The expressions of TLR4, p-p65, and p-IκBα in lipopolysaccharide-induced HFL1 were increased by PTPRO overexpression, and decreased by PTPRO silencing (Figure 5), indicating that the silence of PTPRO suppressed the activation of TLR4/NF-κB to protect against pneumonia.
Figure 5 PTPRO contributed to lipopolysaccharide-induced activation of TLR4/NF-κB in HFL1. (A) Protein expressions of TLR4, p-p65, and p-IκBα in lipopolysaccharide-induced HFL1 were increased by PTPRO overexpression, while they were decreased by PTPRO silence. #,^p ˂ 0.05; ^^p ˂ 0.01, ***,###p ˂ 0.001.
Discussion
PTPs catalyze the dephosphorylation of target genes, thus being implicated in the pathogenesis of several diseases, such as obesity, diabetes, and cancer.16 PTP, PTP-CPS4B, was identified as a potential drug candidate for the prevention of Streptococcus pneumonia.17 This study identified that PTPRO, a novel target for pneumonia, intensified lipopolysaccharide-induced apoptosis and inflammation in HFL1.
Previous studies have shown that lipopolysaccharide, a potent endotoxin, induced inflammatory response in lung fibroblast (HFL1) through production of TNF-α and IL-1α.18 Lipopolysaccharide-induced HFL1 cells were used as cell models of infantile pneumonia.19 In this study, HFL1 cells were also treated with lipopolysaccharide, and the results showed that the cytotoxicity of lipopolysaccharide on HFL1 was achieved through decreasing of cell viability and increasing of cell apoptosis.
Pathogenic infections, such as bacterial and viral infections, induce cell apoptosis in the initial stage of pneumonia.20 Inhibition of lipopolysaccharide-induced cell apoptosis of HFL1 alleviated the progression of infantile pneumonia.19 PTPRO aggravated oxidized low-density lipoprotein-induced cell apoptosis in RAW264.7 cells.21 This study confirmed that knockdown of PTPRO protected HFL1 against lipoprotein-induced cytotoxicity through increasing of cell viability and decreasing of cell apoptosis.
Patients with Pneumocystis jirovecii pneumonia demonstrated higher expression of pro-inflammatory cytokines, including MCP-1, IL-8, IL-6, IL-1α, and TNF-α than the normal people.22 Bacterial and viral infections induce recruitment of polymorphonuclear neutrophils into the alveoli, and promote cell death and multiorganic failure through the secretion of pro-inflammatory cytokines during the development of pneumonia.23 Therefore, anti-inflammatory options, such as TLR antagonists, macrolides, statins, and corticosteroids, showed promising prevention effects for patients with pneumonia.24 PTPRO has been reported to induce the expression of proinflammatory cytokines, such as TNF-α, IL-1α, and IL-6, in lipopolysaccharide--induced macrophages.25 Similarly, lipopolysaccharide--induced upregulations of TNF-α, IL-1α, and IL-18 in HFL1 were reversed by silencing the PTPRO. Therefore, knockdown of PTPRO protected against lipoprotein-induced cytotoxicity in HFL1 through anti-apoptotic and anti-inflammatory abilities. Moreover, patients with severe pneumonia showed alteration in oxidative stress status with deficit of antioxidants and increased lipid peroxidation.26 Oxidative stress is implicated in the pathogenesis of pneumonia.27 PTPRO aggravated oxidized low-density lipoprotein-induced oxidative stress in RAW264.7 cells through decreasing of SOD, and increasing of ROS and MDA.21 Therefore, knockdown of PTPRO might exert anti-oxidant effect on lipopolysaccharide-induced HFL1.
Lipopolysaccharide has been shown to induce activation of TLR4/NF-κB signaling, and promote inflammatory and apoptotic lung injury.28 Inhibition of TLR4/NF-κB signaling reduced lipopolysaccharide-induced lung inflammation.29 PTPRO promoted the activation of TLR4/NF-κB signaling through increasing of TLR4, p-p65, and IκBα to exaggerate inflammatory response in ulcerative colitis.25 Moreover, PTPRO also interacted with TLR4, and promoted TLR4-mediated carcinogenesis and progression of hepatocellular carcinoma through the activation of NF-κB signaling.30 Here, knockdown of PTPRO reduced the protein expression of TLR4, p-p65, and p-IκBα in lipopolysaccharide-induced HFL1 to suppress the inflammation.
In conclusion, PTPRO promoted lipopolysaccharide--induced apoptosis and inflammation in HFL1. Knockdown of PTPRO inhibited the activation of TLR4/NF-κB signaling to attenuate lipopolysaccharide-induced apoptosis and inflammation in HFL1, revealing that PTPRO might be a novel target for the prevention of pneumonia. However, the effect of PTPRO on animal model of pneumonia should be investigated in further research.
Acknowledgements
Not applicable.
Funding
Not applicable.
Competing Interests
The authors state that there are no conflicts of interest to disclose.
Ethics Approval
Not applicable.
Statement of Human and Animal Rights
Not applicable.
Statement of Informed Consent
Not applicable.
Contribution of Authors
Yao Chen and Buming Sun designed the experiments, carried them out. Yao Chen analyzed and interpreted the data. Buming Sun prepared the manuscript with contributions from all co-authors.
REFERENCES
1. Yang X, Yang Y, Wu Y, Fu M. METTL3 promotes inflammation and cell apoptosis in a pediatric pneumonia model by regulating EZH2. Allergol Immunopathol. 2021;49(5):49–56. 10.15586/aei.v49i5.445
2. Eslamy HK, Newman B. Pneumonia in normal and immunocompromised children: an overview and update. Radiol Clin North Am. 2011;49(5):895–920. 10.1016/j.rcl.2011.06.007
3. Hooven TA, Polin RA. Pneumonia. Semin Fetal Neonatal Med. 2017;22(4):206–213. 10.1016/j.siny.2017.03.002
4. Esposito S, Cohen R, Domingo JD, Pecurariu OF, Greenberg D, Heininger U, et al. Do we know when, what and for how long to treat?: Antibiotic therapy for pediatric community-acquired pneumonia. Pediatr Infect Dis J. 2012;31(6):e78–e85. 10.1097/INF.0b013e318255dc5b
5. Hou J, Xu J, Jiang R, Wang Y, Chen C, Deng L, et al. Estrogen-sensitive PTPRO expression represses hepatocellular carcinoma progression by control of STAT3. Hepatology. 2012;57(2):678–688. 10.1002/hep.25980
6. Ren W, Yi H, Bao Y, Liu Y, Gao X. Oestrogen inhibits PTPRO to prevent the apoptosis of renal podocytes. Exp Ther Med. 2019;17(3):2373–2380. 10.3892/etm.2019.7167
7. Gan J, Zhang H. PTPRO predicts patient prognosis and correlates with immune infiltrates in human clear cell renal cell carcinoma. Transl Cancer Res. 2020;9(8):4800–4810. 10.21037/tcr-19-2808
8. Hou X, Du J, Fang H. PTPRO is a therapeutic target and correlated with immune infiltrates in pancreatic cancer. J Cancer. 2021;12(24):7445–7453. 10.7150/jca.64661
9. Motiwala T, Kutay H, Ghoshal K, Bai S, Seimiya H, Tsuruo T, et al. Protein tyrosine phosphatase receptor-type O (PTPRO) exhibits characteristics of a candidate tumor suppressor in human lung cancer. Proc Natl Acad Sci U S A. 2004;101(38):13844. 10.1073/pnas.0405451101
10. Dong H, Xie C, Jiang Y, Li K, Lin Y, Pang X, et al. Tumor-derived exosomal protein tyrosine phosphatase receptor type O polarizes macrophage to suppress breast tumor cell invasion and migration. Front Cell Dev Biol. 2021;9:703537. 10.3389/fcell.2021.703537
11. Zhang X, Tan Z, Wang Y, Tang J, Jiang R, Hou J, et al. PTPRO-associated hepatic stellate cell activation plays a critical role in liver fibrosis. Cell Physiol Biochem. 2015;35(3):885–898. 10.1159/000369746
12. Jiang R, Chen D, Hou J, Tan Z, Wang Y, Huang X, et al. Survival and inflammation promotion effect of PTPRO in fulminant hepatitis is associated with NF-B activation. J Immunol. 2014;193:5161–5170. 10.4049/jimmunol.1303354
13. Cen X, Liu S, Cheng K. The role of toll-like receptor in inflammation and tumor immunity. Front Pharmacol. 2018;9:878.
14. Nong W. Long non-coding RNA NEAT1/miR-193a-3p regulates LPS-induced apoptosis and inflammatory injury in WI-38 cells through TLR4/NF-κB signaling. Am J Transl Res. 2019;11(9):5944–5955. 10.3389/fphar.2018.00878
15. Fan Y, Wang J, Feng Z, Cao K, Xu H, Liu J. Pinitol attenuates LPS-induced pneumonia in experimental animals: possible role via inhibition of the TLR-4 and NF-κB/IκBα signaling cascade pathway. J Biochem Mol Toxicol. 2020;35:e22622. 10.1002/jbt.22622
16. Hendriks WJAJ, Elson A, Harroch S, Pulido R, Stoker A, den Hertog J. Protein tyrosine phosphatases in health and disease. FEBS J. 2013;280(2):708–730. 10.1111/febs.12000
17. Zaman Z, Khan S, Nouroz F, Farooq U, Urooj A. Targeting protein tyrosine phosphatase to unravel possible inhibitors for Streptococcus pneumoniae using molecular docking, molecular dynamics simulations coupled with free energy calculations. Life Sci. 2021;264:118621. 10.1016/j.lfs.2020.118621
18. Lee C-H, Chen J-C, Hsiang C-Y, Wu S-L, Wu H-C, Ho T-Y. Berberine suppresses inflammatory agents-induced interleukin-1β and tumor necrosis factor-α productions via the inhibition of IκB degradation in human lung cells. Pharmacol Res. 2007;56(3):193–201. 10.1016/j.phrs.2007.06.003
19. Cui J, Wang J, Lv Y, Xu D. LncRNA NEAT1 regulates infantile pneumonia by sponging miR-146b. Mol Biotechnol. 2021;63(8):694–701. 10.1007/s12033-021-00331-w
20. Kazzaz J, Horowitz S, Xu J, Khullar P, Niederman M, Fein AM, et al. Differential patterns of apoptosis in resolving and nonresolving bacterial pneumonia. Am J Respir Crit Care Med. 2000;161:2043–2050. 10.1164/ajrccm.161.6.9806158
21. Liang C, Wang X, Hu J, Lian X, Zhu T, Zhang H, et al. PTPRO promotes oxidized low-density lipoprotein induced oxidative stress and cell apoptosis through toll-like receptor 4/nuclear factor κB pathway. Cell Physiol Biochem. 2017;42:495–505. 10.1159/000477596
22. Chou C-W, Lin F-C, Tsai H-C, Chang S-C. The importance of pro-inflammatory and anti-inflammatory cytokines in Pneumocystis jirovecii pneumonia. Med Mycol. 2013;51(7):704–712. 10.3109/13693786.2013.772689
23. Mizgerd JP. Inflammation and pneumonia: why are some more susceptible than others? Clin Chest Med. 2018;39(4):669–676. 10.1016/j.ccm.2018.07.002
24. Meijvis SCA, van de Garde EMW, Rijkers GT, Bos WJW. Treatment with anti-inflammatory drugs in community-acquired pneumonia. J Intern Med. 2012;272(1):25–35. 10.1111/j.1365-2796.2012.02554.x
25. Zhao J, Yan S, Zhu X, Bai W, Li J, Liang C. PTPRO exaggerates inflammation in ulcerative colitis through TLR4/NF-κB pathway. J Cell Biochem. 2020;121(2):1061–1071. 10.1002/jcb.29343
26. Pincemail J, Cavalier E, Charlier C, Cheramy-Bien J-P, Brevers E, Courtois A, et al. Oxidative stress status in COVID-19 patients hospitalized in intensive care unit for severe pneumonia. A pilot study. Antioxidants (Basel). 2021;10(2):257. 10.3390/antiox10020257
27. Chen Y, Luo G, Yuan J, Wang Y, Yang X, Wang X, et al. Vitamin C mitigates oxidative stress and tumor necrosis factor-alpha in severe community-acquired pneumonia and LPS-induced macrophages. Mediat Inflam. 2014;2014:426740. 10.1155/2014/426740
28. Zhang H, Lang W, Wang S, Li B, Li G, Shi Q. Echinacea polysaccharide alleviates LPS-induced lung injury via inhibiting inflammation, apoptosis and activation of the TLR4/NF-κB signal pathway. Int Immunopharmacol. 2020;88:106974. 10.1016/j.intimp.2020.106974
29. Pan W, Xu X, Wang Y, Song X. Interleukin-35 reduces inflammation in acute lung injury through inhibiting TLR4/NF-κB signaling pathways. Exp Ther Med. 2020;19(3):1695–700. 10.3892/etm.2020.8407
30. Xu D, Wang X, Yan S, Yin Y, Hou J, Wang X, et al. Interaction of PTPRO and TLR4 signaling in hepatocellular carcinoma. Tumor Biol. 2014;35(10):10267–10273. 10.1007/s13277-014-2302-5