
Download




ORIGINAL ARTICLE
Tricin attenuates the progression of LPS-induced severe pneumonia in bronchial epithelial cells by regulating AKT and MAPK signaling pathways
Fan Yanga, Wenming Liub*
aDepartment of Emergency, The Affiliated Changzhou No. 2 People’s Hospital of Nanjing Medical University, Changzhou, China
bDepartment of Critical Care Medicine, The Affiliated Changzhou No. 2 People’s Hospital of Nanjing Medical University, Changzhou, China
Abstract
Background Pneumonia is a continuous and widespread disease with higher incidence, the effects of it on human life can be fearful. Tricin has been demonstrated to take part in the progression and development of diseases. However, the function of Tricin and its related regulatory pathways remain unclear. This study was planned to investigate the effects of Tricin on severe pneumonia.
Methods The cell viability was detected through CCK-8 assay. The TNF-α, IL-1β and IL-6 levels were assessed through ELISA and RT-qPCR. The levels of MDA, SOD and GSH were tested through corresponding commercial kits. The protein expressions were examined through western blot.
Results In our study, the lipopolysaccharide (LPS) was firstly used to stimulate cell model for severe pneumonia. We discovered that Tricin had no toxic effects on BEAS-2B cells and the decreased cell viability induced by LPS was relieved by a dose-dependent Tricin treatment. Additionally, through ELISA and RT-qPCR, it was uncovered that Tricin reduced the LPS-induced inflammation through regulating TNF-α, IL-1β and IL-6. Furthermore, Tricin relieved LPS-induced oxidative stress through reducing MDA level and enhancing SOD and GSH levels. Finally, it was demonstrated that Tricin retarded LPS-activated AKT and MAPK pathways.
Conclusion Our findings revealed that Tricin attenuated the progression of LPS induced severe pneumonia through modulating AKT and MAPK signaling pathways. This discovery might afford one novel sight for the treatment of severe pneumonia.
Key words: AKT and MAPK pathways, LPS, severe pneumonia, tricin
*Corresponding author: Wenming Liu, Department of Critical Care Medicine, The Affiliated Changzhou No. 2 People’s Hospital of Nanjing Medical University, No. 68, Middle Gehu Road, Wujin District, Changzhou, Jiangsu 213000, China. Email address: [email protected]
Received 20 January 2022; Accepted 25 February 2022; Available online 1 May 2022
Copyright: Yang F and Liu W
License: This open access article is licensed under Creative Commons Attribution 4.0 International (CC BY 4.0). http://creativecommons.org/licenses/by/4.0/
Introduction
Inflammatory lung diseases including acute lung injury, pneumonia, and chronic obstructive pulmonary disease result in serious morbidity and mortality worldwide and pose a major risk to public health.1,2 Pneumonia is a chronic and recurrent disease. Patients are diagnosed with severe pneumonia if ventilation support, circulatory support, and intensive monitoring, as well as treatment, are required.3,4 One in five patients hospitalized with pneumonia is admitted to the intensive care unit, and one-third of them require mechanical ventilation with high mortality rates.4 Factors that cause severe pneumonia include age, antibiotic resistance, septic shock, and acute respiratory failure.5 Lipopolysaccharide (LPS) is a strong stimulator that can stimulate the production of proinflammatory factors, including TNF-α, IL-1β, and IL-6, thereby triggering a systemic inflammatory response.6,7 Studies on severe pneumonia have gained increasing attention from basic and clinical researchers, and it is essential to ameliorate the outcomes of severe pneumonia.
Tricin is the main flavonoid component of barley, wheat, oats, and rice. Tricin confers multiple health effects, such as antiviral, anti-inflammation, antioxidant, and antitumor properties, cardiovascular protection, and so on.8–10 Tricin has been shown to suppress inflammatory response in human PBMC cells through modulating p38/MAPK and PI3K pathways.11 Besides, tricin can relieve inflammatory response of raw 264.7 cells mediated by LPS.12 In addition, tricin can act on the Lyn/Syk pathway to inhibit the hypersensitivity of mast cells.13 Moreover, tricin may affect retinal diseases through its antioxidative effects.14 However, there is no relevant study on tricin in severe pneumonia.
Therefore, this study was designed to investigate the effects and the related mechanisms of tricin on severe pneumonia. In the current study, tricin was found to inactivate AKT and MAPK signaling pathways to attenuate the progression of LPS-induced severe pneumonia in bronchial epithelial cells. This finding may offer a useful therapeutic drug for severe pneumonia.
Materials and Methods
Cell culture
BEAS-2B cells were acquired from American Type Culture Collection (ATCC, Manassas, VA, USA). Cells were maintained in RPMI-1640 medium containing 10% fetal bovine serum (FBS, Gibco, NY, USA) in a humidified atmosphere (5% CO2 and 95% air) at 37°C. Tricin (Tokyo Chemical Industry Co., Ltd.; 0, 5, 10, 20, or 50 μM) or LPS (from Escherichia coli 055:B5, Solarbio, Beijing, China; 10 μg/mL) was used to treat the cells. The concentration of tricin used in this study was in accordance with previous publications11,15,16 and based on the results of our preliminary experiments.
MTT assay
To evaluate cell viability, BEAS-2B cells (1×104 cells/well) were plated into 96-well plates following tricin or LPS treatment. At 24 h post-treatment, MTT solution (20 μL; 5 mg/mL; Sigma-Aldrich) was added into each well and incubated for another 4 h. Then, DMSO (200 µL, Sigma-Aldrich) was added, and the absorbance was measured at 490 nm using a microplate reader (BioTek Instruments, Inc.).
RT-qPCR
The TRIzol reagent (Thermo Fisher Scientific, Inc.) was used to isolate total RNA from BEAS-2B cells. Reverse transcription of RNA into cDNA was carried out using a PrimeScript RT reagent kit (Takara Biotechnology Co., Ltd.). Quantitative polymerase chain reaction (qPCR) was performed using the SYBR Premix Ex Taq (Takara Biotechnology Co., Ltd.). The relative expression was calculated using the 2−ΔΔCt method. The expression of TNF-α, IL-1β, and IL-6 was normalized to the internal reference gene, GAPDH. The primers used in this study are as follows:
TNF-α:
Forward: 5’-CCGGGAGAAGAGGGATAGCTT-3’,
Reverse: 5’-TCGGACAGTCACTCACCAAGT-3’;
IL-6:
Forward: 5’-TAGTCCTTCCTACCCCAATTTCC-3’,
Reverse: 5’-TTGGTCCTTAGCCACTCCTTC-3’;
IL-1β:
Forward: 5’-GAAATGCCACCTTTTGACAGTG-3’,
Reverse: CTGGATGCTCTCATCAGGACA-3’;
GAPDH:
Forward: 5’-CTTTGGTATCGTGGAAGGACTC-3’,
Reverse: 5’-GTAGAGGCAGGGATGATGTTCT-3’.
Western blot
Total protein from BEAS-2B cells was isolated using RIPA buffer (Beyotime, Shanghai, China). Proteins were separated on 10% SDS- PAGE gel (Beyotime, Shanghai, China) and transferred onto poly(vinylidene difluoride) (PVDF) membranes (Millipore, MA, USA). After blocking with 5% skim milk, membranes were incubated with primary antibodies for 12 h at 4°C. The primary antibodies used in this study are as follows: p-AKT (ab38449, 1:1000, Abcam), AKT (ab8805, 1:1000, Abcam), p-p38 (ab178867, 1:1000, Abcam), p38 (ab45136, 1:1000, Abcam), p-JNK (ab76572, 1:1000, Abcam), JNK (ab179461, 1:1000, Abcam), p-ERK (ab214036, 1:1000, Abcam), ERK (ab184699, 1:1000, Abcam), and GAPDH (ab9484, 1:1000, Abcam). After washing, membranes were incubated with secondary antibodies (ab6721, 1:2000, Abcam) at room temperature for 2 h. Next, protein bands were visualized using the ECL detection kit (Beyotime Institute of Biotechnology) and analyzed using the ImageJ software.
ELISA
Human TNF-α/IL-1β/IL-6 enzyme-linked immunosorbent assay (ELISA) Kit (RayBiotech) was used to assess the expression levels of TNF-α/IL-1β/IL-6 in the culture medium of BEAS-2B cells, according to the manufacturer’s instructions. The levels of TNF-α/IL-6 and IL-1β are presented as ng/mL and pg/mL, respectively.
Detection of MDA, SOD, and GSH
The malondialdehyde (MDA, A003-1), superoxide dismutase (SOD, A001-3), and glutathione (GSH, A006-2) commercial kits were purchased from Jiancheng Biotechnology Research Institute (Nanjing, China). The levels of MDA, SOD, and GSH were assessed according to the manufacturer’s instructions.
Statistical analysis
SPSS 20.0 (IBM Corp., Armonk, NY, USA) was used to perform the statistical analyses. All data are presented as mean ± standard deviation (SD). All experiments were repeated three times. Student’s t-test or one-way analysis of variance (ANOVA) with Dunnett’s multiple comparison test was employed to analyze the difference between the two or multiple groups, respectively. Data were tested for normal distribution and homogeneity of variance. p<0.05 was considered statistically significant.
Results
Effects of Tricin/LPS on the cell viability of BEAS-2B cells
MTT assay was performed to investigate the effect of tricin treatment on the cell viability of BEAS-2B cells. The results showed that tricin did not affect cell viability at any concentration tested, indicating that tricin had no toxic effects on BEAS-2B cells (Figure 1A). This was in contrast with LPS treatment that led to significant cell cytotoxicity. Interestingly, LPS-induced cell cytotoxicity was relieved by tricin treatment in a dose-dependent manner (Figure 1B).
Figure 1 The effects of Tricin/LPS on the cell viability of BEAS-2B cells. (A) Cell viability following treatment with 0, 5, 10, 20 or 50 μM Tricin was assessed using MTT assay. (B) Cell viability was assessed using MTT assay in the control, LPS + 0 μM, LPS + 5 μM, LPS + 10 μM, LPS + 20 μM and LPS + 50 μM groups. @@P<0.01 vs control; $P<0.05, $$P<0.01 vs LPS + 0 μM.
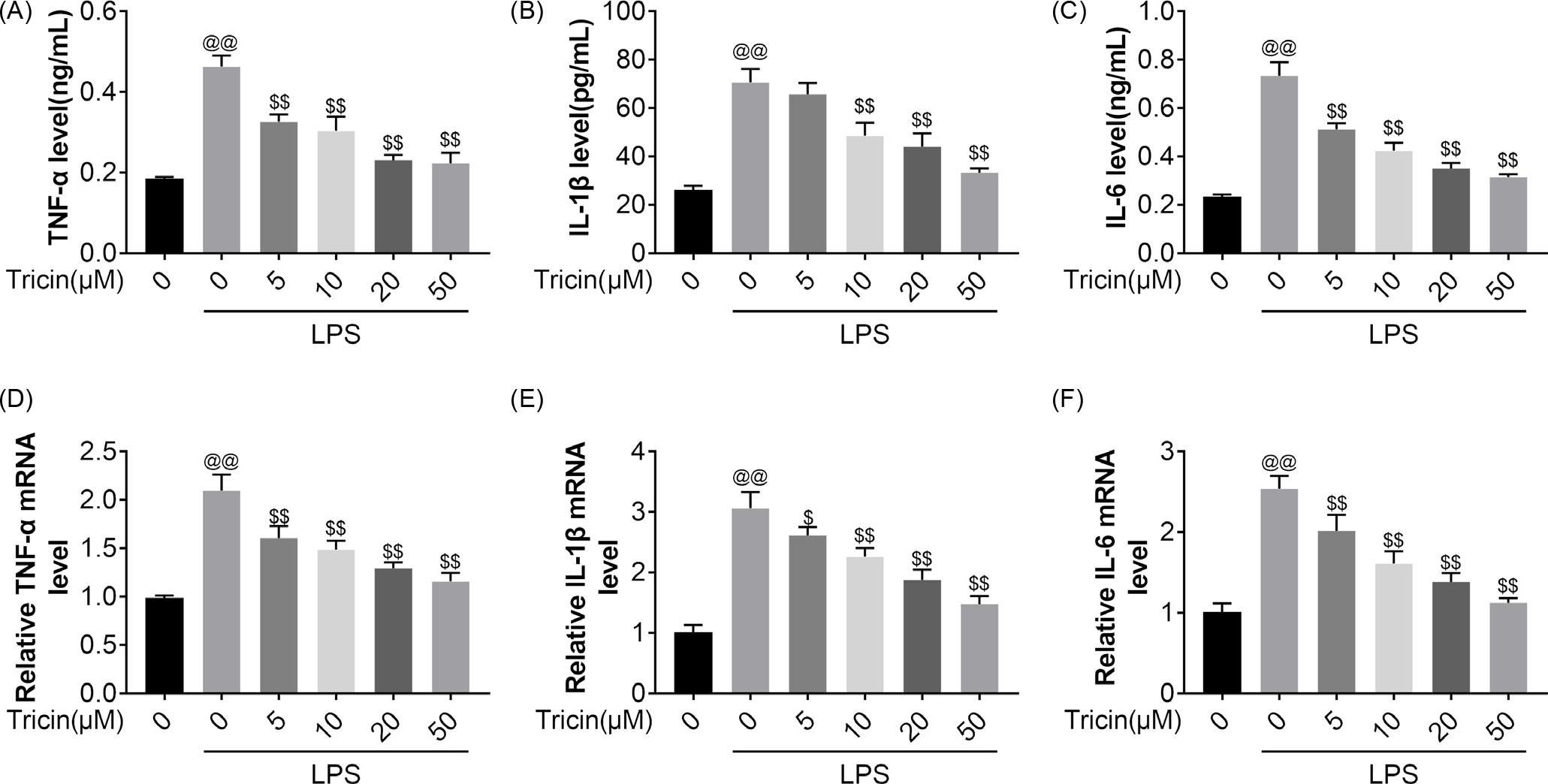
Tricin reduces LPS-induced inflammation
As shown in Figure 2A–C, LPS-mediated increase in TNF-α, IL-1β, and IL-6 levels was offset by tricin treatment. This was consistent with the results from quantitative reverse transcription PCR (RT-qPCR), which showed the same trend in the transcript levels (Figure 2D–F). Taken together, these data indicate that tricin reduces LPS-induced inflammation.
Figure 2 The effects of Tricin on LPS-induced inflammation. Treatment groups include control, LPS + 0 μM, LPS + 5 μM, LPS + 10 μM, LPS + 20 μM, and LPS + 50 μM group. (A–C) The levels of TNF-α, IL-1β and IL-6 were measured using ELISA. (D–F) The mRNA expression of TNF-α, IL-1β and IL-6 was detected using RT-qPCR. @@P<0.01 vs control; $P<0.05, $$P<0.01 vs LPS + 0 μM.
Tricin weakens LPS-induced oxidative stress
The MDA level was enhanced after LPS treatment, which was reversed by adding tricin (Figure 3A). Besides, LPS-mediated reduction in SOD and GSH levels was also neutralized by tricin treatment (Figure 3B,C). Thus, these results suggest that tricin weakens LPS-induced oxidative stress.
Figure 3 The effects of Tricin on LPS-induced oxidative stress. Treatment groups include control, LPS + 0 μM, LPS + 5 μM, LPS + 10 μM, LPS + 20 μM, and LPS + 50 μM group. (A–C) The levels of MDA, SOD and GSH were tested using corresponding commercial kits. @@P<0.01 vs control; $P<0.05, $$P<0.01 vs LPS + 0 μM.
Tricin attenuates LPS-activated AKT and MAPK pathways
The AKT and MAPK pathways play a key role in cancer progression. Thus, in order to explore whether tricin has any effect on LPS-mediated activation of AKT and MAPK pathways, western blot was performed to assess the levels of AKT and MAPK pathway components following tricin treatment. We demonstrated that the protein expression of p-AKT/AKT, p-p38/p38, p-JNK/JNK, and p-ERK/ERK was upregulated after LPS treatment, but this was reversed by adding tricin (Figure 4). Altogether, these data indicate that tricin attenuates LPS-mediated activation of AKT and MAPK pathways.
Figure 4 The effects of Tricin on LPS-activated AKT and MAPK pathways. Treatment groups include control, LPS + 0 μM, LPS + 5 μM, LPS + 10 μM, LPS + 20 μM, and LPS + 50 μM group. The protein expression of p-AKT, AKT, p-p38, p38, p-JNK, JNK, p-ERK and ERK was assessed using western blot. @@P<0.01 vs control; $$P<0.01 vs LPS + 0 μM.
Figure 5
Discussion
In this study, our results have demonstrated that tricin treatment relieved LPS-induced cell cytotoxicity, inflammation, and oxidative stress. Furthermore, our findings have shown that tricin attenuated LPS-mediated activation of AKT and MAPK pathways. Thus, altogether, our data suggest that tricin attenuated the progression of LPS-induced severe pneumonia through modulating AKT and MAPK signaling pathways.
Recently, an increasing number of drugs have been discovered to confer protective effects against different lung diseases. For example, rutin provides a protective effect against LPS-mediated acute lung injury.17 Icariin modulates the Smad and MAPK pathways to relieve TGF-β1-stimulated epithelial-mesenchymal transition.18 Arenaria kansuensis activates the Nrf2 pathway and attenuates the NF-κB/TGF-β1/Smad2/3 pathway to ameliorate pulmonary fibrosis in mice.19 Baicalin regulates the PDK1/AKT pathway to suppress EMT progression in non-small cell lung cancer.20 Ginsenoside Rg1 attenuates inflammation and endoplasmic reticulum stress to relieve sepsis-mediated lung injury through regulating SIRT1.21 In the current study, LPS was used to treat cells to simulate a cell model for severe pneumonia. We have demonstrated that tricin had no toxic effects on BEAS-2B cells and LPS-induced cell cytotoxicity was reversed by tricin treatment in a dose-dependent manner.
In addition, inflammation and oxidative stress have been reported to play a key role in driving lung disease progression. For instance, NLRP9b knockdown attenuates inflammation and oxidative stress to ameliorate acute lung injury.22 Furthermore, CD28 modulates the PI3K/Akt/FoxO1 pathway to affect blast exposure-stimulated lung inflammation, oxidative stress, and T cell accumulation.23 In addition, zingerone affects TGF-β1 and iNOS expression in bleomycin-mediated pulmonary fibrosis to improve oxidative stress and inflammation.24 Pterostilbene weakens oxidative stress and inflammation to resist LPS-stimulated early pulmonary fibrosis in vivo.25 Besides, N-acetylcysteine ameliorates inflammatory response and oxidative stress in community-acquired pneumonia.26 In this study, using ELISA and RT-qPCR, we have shown that tricin reduced LPS-induced inflammation through modulating TNF-α, IL-1β, and IL-6 at both the protein and mRNA levels. Furthermore, tricin relieved LPS-induced oxidative stress through reducing the MDA level and enhancing SOD and GSH levels.
It is widely known that AKT and MAPK pathways are the key regulators of cell survival, cell apoptosis, inflammation, and oxidative stress. For instance, gallic acid modulates the MAPK/NF-κB and Akt/AMPK/Nrf2 pathways to relieve LPS-triggered inflammation and oxidative stress.27 Moreover, unripe Carica papaya regulates the Akt/MAPK/NF-κB pathway to reduce methylglyoxal-stimulated endothelial cell apoptosis and inflammation.28 Schisandrin A modulates the NF-κB, MAPKs, and PI3K/Akt pathways in RAW 264.7 macrophages to inhibit LPS-evoked inflammation and oxidative stress.29 However, it remains unknown whether tricin can regulate AKT and MAPK pathways to achieve anti-inflammatory and antioxidative stress effects in severe pneumonia. In this study, our findings have demonstrated that tricin attenuated LPS-activated AKT and MAPK pathways.
In conclusion, this study has demonstrated for the first time the role of tricin in severe pneumonia and the pathways regulated by tricin. Our findings uncover that tricin attenuates the progression of LPS-induced severe pneumonia through modulating AKT and MAPK signaling pathways. However, there are limitations in this study, and we aim to elucidate further in our future works.
Competing interests
The authors declare no conflicts of interest with respect to research, authorship and/or publication of this article.
Contribution of authors
Fan Yang and Wenming Liu designed the experiments, carried them out, analyzed and interpreted the data, and prepared the manuscript with contributions from all co-authors.
REFERENCES
1. Barba T, Mainbourg S, Nasser M, Lega JC, Cottin V. Lung diseases in inflammatory myopathies. Semin Respir Crit Care Med. 2019;40(2):255–70. 10.1055/s-0039-1685187
2. Liu Y, Gao H, Wang X, Zeng Y. Methylation of inflammatory cells in lung diseases. Adv Exp Med Biol. 2020;1255:63–72. 10.1007/978-981-15-4494-1_5
3. Cunha BA. Pneumonia in the elderly. Clin Microbiol Infect. 2001;7(11):581–8. 10.1046/j.1198-743x.2001.00328.x
4. Mizgerd JP. Pathogenesis of severe pneumonia: advances and knowledge gaps. Curr Opin Pulm Med. 2017;23(3):193–7. 10.1097/MCP.0000000000000365
5. Wang J, Song YL. Advances in severe community-acquired pneumonia. Chin Med J. 2019;132(16):1891–3. 10.1097/CM9.0000000000000366
6. Gao W, Yang H. MicroRNA-124-3p attenuates severe community-acquired pneumonia progression in macrophages by targeting tumor necrosis factor receptor-associated factor 6. Int J Mol Med. 2019;43(2):1003–10. 10.3892/ijmm.2018.4011
7. Sun W, Cheng Z, Chen H, Lin G, Chen H. Tetrahydropyrimidines, ZL-5015 alleviated lipopolysaccharide (LPS)-induced acute pneumonia in rats by activating the NRF-2/HO-1 pathway. Med Sci Monit. 2020;26:e924482. 10.12659/MSM.924482
8. Lam PY, Lui ACW, Wang L, et al. Tricin biosynthesis and bioengineering. Front Plant Sci. 2021;12:733198. 10.3389/fpls.2021.733198
9. Jiang B, Song J, Jin Y. A flavonoid monomer tricin in Gramineous plants: metabolism, bio/chemosynthesis, biological properties, and toxicology. Food Chem. 2020;320:126617. 10.1016/j.foodchem.2020.126617
10. Chang CL, Wang GJ, Zhang LJ, et al. Cardiovascular protective flavonolignans and flavonoids from Calamus quiquesetinervius. Phytochemistry. 2010;71(2–3):271–9. 10.1016/j.phytochem.2009.09.025
11. Shalini V, Pushpan CK, Sindhu G, Jayalekshmy A, Helen A. Tricin, flavonoid from Njavara reduces inflammatory responses in hPBMCs by modulating the p38MAPK and PI3K/Akt pathways and prevents inflammation associated endothelial dysfunction in HUVECs. Immunobiology. 2016;221(2):137–44. 10.1016/j.imbio.2015.09.016
12. Kang BM, An BK, Jung WS, et al. Anti-inflammatory effect of tricin isolated from alopecurus aequalis sobol. on the LPS-induced inflammatory response in RAW 264.7 cells. Int J Mol Med. 2016;38(5):1614–20. 10.3892/ijmm.2016.2765
13. Lee JY, Park SH, Jhee KH, Yang SA. Tricin isolated from enzyme-treated Zizania latifolia extract inhibits IgE-mediated allergic reactions in RBL-2H3 cells by targeting the Lyn/Syk pathway. Molecules. 2020;25(9):2084. 10.3390/molecules25092084
14. Lee HJ, Kim KA, Kang KD, et al. The compound isolated from the leaves of Phyllostachys nigra protects oxidative stress-induced retinal ganglion cells death. Food Chem Toxicol. 2010;48(6):1721–7. 10.1016/j.fct.2010.03.052
15. Shalini V, Jayalekshmi A, Helen A. Mechanism of anti-inflammatory effect of tricin, a flavonoid isolated from Njavara rice bran in LPS induced hPBMCs and carrageenan induced rats. Mol Immunol. 2015;66(2):229–39. 10.1016/j.molimm.2015.03.004
16. Li X-X, Chen S-G, Yue GG, et al. Natural flavone tricin exerted anti-inflammatory activity in macrophage via NF-κB pathway and ameliorated acute colitis in mice. Phytomedicine. 2021;90:153625. 10.1016/j.phymed.2021.153625
17. Tian C, Shao Y, Jin Z, et al. The protective effect of rutin against lipopolysaccharide induced acute lung injury in mice based on the pharmacokinetic and pharmacodynamic combination model. J Pharm Biomed Anal. 2021;209:114480. 10.1016/j.jpba.2021.114480
18. Li Z, Yuan X, Wang B, Gao F. Icariin alleviates transforming growth factor-β1-induced epithelial-mesenchymal transition by targeting Smad and MAPK signaling pathways. Am J Transl Res. 2020;12(2):343–60.
19. Cui Y, Xin H, Tao Y, Mei L, Wang Z. Arenaria kansuensis attenuates pulmonary fibrosis in mice via the activation of Nrf2 pathway and the inhibition of NF-kB/TGF-beta1/Smad2/3 pathway. Phytother Res. 2021;35(2):974–86. 10.1002/ptr.6857
20. Chen J, Yuan CB, Yang B, Zhou X. Baicalin inhibits EMT through PDK1/AKT signaling in human nonsmall cell lung cancer. J Oncol. 2021;2021:4391581. 10.1155/2021/4391581
21. Wang QL, Yang L, Peng Y, et al. Ginsenoside Rg1 regulates SIRT1 to ameliorate sepsis-induced lung inflammation and injury via inhibiting endoplasmic reticulum stress and inflammation. Mediat Inflamm. 2019;2019:6453296. 10.1155/2019/6453296
22. Yanling Q, Xiaoning C, Fei B, Liyun F, Huizhong H, Daqing S. Inhibition of NLRP9b attenuates acute lung injury through suppressing inflammation, apoptosis and oxidative stress in murine and cell models. Biochem Biophys Res Commun. 2018;503(2):436–43. 10.1016/j.bbrc.2018.04.079
23. Liu Y, Tong C, Xu Y, et al. CD28 Deficiency ameliorates blast exposure-induced lung inflammation, oxidative stress, apoptosis, and t cell accumulation in the lungs via the PI3K/Akt/FoxO1 signaling pathway. Oxid Med Cell Longev. 2019;2019:4848560. 10.1155/2019/4848560
24. Gungor H, Ekici M, Karayigit MO, Turgut NH, Kara H, Arslanbas E. Zingerone ameliorates oxidative stress and inflammation in bleomycin-induced pulmonary fibrosis: modulation of the expression of TGF-β1 and iNOS. Naunyn Schmiedebergs Arch Pharmacol. 2020;393(9):1659–70. 10.1007/s00210-020-01881-7
25. Yang H, Hua C, Yang X, et al. Pterostilbene prevents LPS-induced early pulmonary fibrosis by suppressing oxidative stress, inflammation and apoptosis in vivo. Food Funct. 2020;11(5):4471–84. 10.1039/C9FO02521A
26. Zhang Q, Ju Y, Ma Y, Wang T. N-acetylcysteine improves oxidative stress and inflammatory response in patients with community acquired pneumonia: a randomized controlled trial. Medicine. 2018;97(45):e13087. 10.1097/MD.0000000000013087
27. Tanaka M, Kishimoto Y, Sasaki M, et al. Terminalia bellirica (Gaertn.) roxb. extract and gallic acid attenuate LPS-induced inflammation and oxidative stress via MAPK/NF-κB and Akt/AMPK/Nrf2 pathways.Oxid Med Cell Longev. 2018;2018:9364364. 10.1155/2018/9364364
28. Jarisarapurin W, Kunchana K, Chularojmontri L, Wattanapitayakul SK. Unripe carica papaya protects methylglyoxal-invoked endothelial cell inflammation and apoptosis via the suppression of oxidative stress and Akt/MAPK/NF-κB signals. Antioxidants. 2021;10(8):1158. 10.3390/antiox10081158
29. Kwon DH, Cha HJ, Choi EO, et al. Schisandrin A suppresses lipopolysaccharide-induced inflammation and oxidative stress in RAW 264.7 macrophages by suppressing the NF-κB, MAPKs and PI3K/Akt pathways and activating Nrf2/HO-1 signaling.Int J Mol Med. 2018;41(1):264–74. 10.3892/ijmm.2017.3209