
Download
ORIGINAL ARTICLE
Momordica charantia L. improves airway hyperresponsiveness and suppresses inflammation in a murine model of allergic asthma
Clara do Prado Lopes Frotaa, Alessa Castro Ribeiroa, Marco Túlio Alvarenga Calil Filhoa, Lucas Junqueira de Freitas Morelb, Leandra Naira Zambelli Ramalhoa, Jéssica Cristina Amaralc, Silvia Helena Taleb Continib, Vanessa Maciel Braulio da Fonsecaa, Marcos de Carvalho Borgesa, Ana Maria Soares Pereirab, d, Fabio Carmonaa*
aRibeirao Preto Medical School, University of Sao Paulo, Ribeirao Preto, Brazil
bDepartment of Plant Biotechnology, University of Ribeirao Preto, Ribeirao Preto, Brazil
cDepartment of Chemistry, Federal University of Sao Carlos, São Carlos, Brazil
dBotanical Garden of Medicinal Plants Ordem e Progresso, Jardinopolis, Brazil
Abstract
Objective To evaluate the effect of a hydroethanolic extract of Momordica charantia L. (“bitter melon”, Cucurbitaceae) leaves (MCHA) on ovalbumin (OVA)-induced asthma model. Balb/c mice were sensitized twice and challenged for 4 alternate days with OVA and then treated with MCHA (500 mg/kg) for 7 consecutive days.
Methods Control groups received treatment with normal saline or dexamethasone (2 mg/kg) on the same day. We assessed in vivo bronchial hyperresponsiveness and ex-vivo inflammation and mucus production in bronchoalveolar lavage (BAL), lung homogenates, and lung tissue.
Results MCHA significantly improved airway hyperresponsiveness near baseline levels. MCHA administration significantly improved airway and lung inflammation, demonstrated by decreased total and inflammatory cells in BAL, lower levels of IL-5 and IL-13 in lung homogenate, and fewer inflammatory cells in lung tissue. Additionally, MCHA significantly diminished goblet cells in lung tissue.
Conclusions Administration of a hydroethanolic extract of M. charantia leaves was effective in treating OVA-induced asthma in an animal model.
Key words: allergy, Cucurbitaceae, herbal medicine, inflammation, ovalbumin
*Corresponding author: Fabio Carmona. Departamento de Puericultura e Pediatria FMRP-USP, Avenida dos Bandeirantes S/N, Monte Alegre, Campus Universitario, Ribeirão Preto, SP 14049-900, Brazil. Email address: [email protected]
Received 1 January 2022; Accepted 27 January 2022; Available online 1 March 2022
Copyright: Clara do Prado Lopes Frota, et al.
License: This open access article is licensed under Creative Commons Attribution 4.0 International (CC BY 4.0). http://creativecommons.org/licenses/by/4.0/
Introduction
Asthma is one of the most common human chronic diseases, affecting 1–18% of adults and children in different countries and about 300 million people worldwide.1 In 2011, the Brazilian Public Health System’s national database registered asthma as the fourth leading cause of hospital admissions across all ages.2
Inflammation is the hallmark of asthma, being present even in asymptomatic patients. Consequently, treatment with anti-inflammatory drugs, such as inhaled corticosteroids, is frequently used. The response to asthma treatment is widely variable among patients and some do not achieve adequate disease control.1 Therefore, new and safer drugs with different therapeutic targets are still needed.
Momordica charantia L. (bitter melon, Cucurbitaceae), listed on The Plant List,3 is a medicinal plant with anti-inflammatory properties that could benefit asthma treatment. This species grows in tropical areas, including parts of the Amazon, eastern Africa, Asia, and the Caribbean. Populations have extensively used this plant for diabetes and inflammatory diseases but not for asthma.4,5 Therefore, the objective of this study was to evaluate the effect of a M. charantia extract in asthma treatment in a murine model.
Materials and Methods
Animals
Balb/c-specific pathogen-free male mice (6–8 weeks old) with free access to food and water were used. The local Committee for Ethics in Animal Experiments, Ribeirao Preto Medical School, University of Sao Paulo approved the protocol (#072/2013).
Preparation of the M. charantia leaf hydroethanolic extract
The species is cultivated in the Botanical Garden of Medicinal Plants Ordem e Progresso, in Jardinopolis, Sao Paulo, Brazil (21°4'33″ S, 47°44'48″ W), with access permission from the Brazilian government (SisGen protocol #A03C96C). The leaves were harvested at 9 a.m., weighted, and washed with water. Excess water was removed using a filter paper for 2 h. The specimen was identified by Jorge Yoshio Tamashiro, from the Department of Botany, Institute of Biology, State University of Campinas (UNICAMP), Sao Paulo, Brazil. A voucher specimen was deposited at the Herbarium of the University of Ribeirao Preto (UNAERP) with identification code HPMU2844. The leaves were then dried in an oven with circulating air at 45°C for 24 h. They were then ground in a knife mill and sieved to standardize the particle size (40 mesh). The hydroethanolic extract (MCHA) was prepared by adding 100 g of powdered leaves to 1 L of 70% ethanol (v/v in water), macerating for 10 days, filtering, and rota-evaporating it. Before use, the extract was re-suspended in normal saline plus 5% dimethyl sulfoxide (DMSO).
Chromatographic analysis of M. charantia leaf hydroethanolic extract
Chromatographic analysis was done at the Department of Chemistry, Federal University of Sao Carlos (UFSCAR), Sao Paulo, Brazil. Liquid chromatography–high-resolution mass spectrometry (LC-HRMS) analysis of the M. charantia leaf hydroethanolic extract was performed using an LC-HRMS Q-TOF Agilent-6545 equipped with an Agilent, Zorbax Eclipse Plus C-18 (2,1 × 50 mm2) –1,8 μm column. The chromatographic method consisted of a gradient elution of H2O + 0,1% formic acid:CH3CN + 0.1% formic acid as follows: 95:5 to 0:100 in 25 min. The flow rate and the injection volume were 0.35 mL.min‒1 and 2 μL, respectively. The column temperature was 40°C. Mass spectra were acquired in negative and positive modes over the range m/z 100–1700. The capillary voltage was set at 3000 V; nozzle voltage at 500 V; gas temperature at 350°C; drying gas,13 L min‒1; nebulizer, 60 psi; sheaf gas, 300°C; fragmentor, 140 V; skimmer, 80 V; and collision energy voltage 10–at 30 V. The LC-MS data were processed using the software Qualitative Navigator B.08.00 and Qualitative Workflows B.08.00.6–8
Allergen sensitization and challenge protocol
Mice were sensitized with 10 µg of ovalbumin (OVA, Sigma grade V, Sigma-Aldrich, Dorset, UK) and 1 mg of Al(OH)3 (Sigma-Aldrich, Dorset, UK) intraperitoneally (i.p.) twice, 7 days apart. One week later, mice were challenged with 10 µg of intranasal OVA on 4 alternate days under light anesthesia (ketamine and xylazine). Control groups received normal saline (SAL) on the same day.
Treatment with extract of M. charantia
Six groups were studied (challenged with/treated with): SAL/SAL, OVA/SAL, OVA/MCHA, OVA/DEXA, and SAL/MCHA. MCHA (500 mg/kg i.p.) was administered for 7 consecutive days, beginning with the first challenge. Animals from the control groups received dexamethasone (DEXA, 2 mg/kg) or SAL on the same day. Experiments were performed 24 h after the last challenge.
In vivo assessment of respiratory mechanics
Respiratory parameters were assessed as previously described.9 Briefly, 24 h after the last challenge, animals were anesthetized (xylazine and ketamine). A tracheostomy was performed, and they were connected via a tracheal cannula to a ventilator for small animals (FlexiVent®, Scireq, Canada). The animals were ventilated with 150 breaths/min and positive end-expiratory pressure (PEEP) of 3 cm H2O. Pulmonary mechanics were measured under anesthesia and muscular blockade (pancuronium). Bronchial hyperresponsiveness was measured at baseline and increasing concentrations of methacholine aerosol (6.25, 12.5, 25, and 50 mg/mL), for 10 s each, delivered by an ultrasonic nebulizer (Hudson RCI, Teleflex Medical, USA). Only curves with a coefficient of determination (COD) of 0.85 or more were analyzed. The following parameters were analyzed: total resistance (RRS), total elastance (ERS), tissue resistance (G), and tissue elastance (H).
Bronchoalveolar lavage (BAL) collection and lung preparation
After respiratory mechanics measurements, BAL was collected as previously described.10 After, the animals were sacrificed for lung collection. A sample (50 mg) of right lung tissue was homogenized in PBS with antiprotease (Complete, EDTA-free, Roche, USA; 50 mg/mL) until completely dissolved. The sample was then centrifuged, and the supernatant was stored at ‒80°C for cytokine analyses. The left lung was insufflated with 10% neutral buffered formalin (NBF) solution, fixed in NBF for 3 days, and embedded in paraffin for histological analysis.
Assessment of inflammation, oxidative stress, and mucus production
Total and differential cell counts in BAL
Total cell count in BAL was performed using trypan blue exclusion. BAL cytospin slides were fixed and stained with Panotico Rapido LB (Laborclin, Pinhais, PR, Brazil). Differential cell counts were determined by counting 300 cells in each sample.
Assessment of inflammation in lung homogenates
Interleukins (ILs)-4, -5, -10, and -13 were measured by ELISA (BD Biosciences BD OptEIATM, USA, and eBiosciences, USA) in lung homogenates, according to the manufacturer’s instructions.
Oxidative stress evaluation in lung homogenates
The total antioxidant capacity (TAC) was assessed in the lung homogenate by reducing radical cation 2,2'-azino- bis(3-ethylbenzothiazoline-6-sulfonic acid) (ABTS+) (Sigma-Aldrich, Dorset, UK), as previously described.11
Histological assessment of lung inflammation and mucus
Paraffin-embedded lung tissue was sectioned (5 µm) and stained with hematoxylin and eosin (H&E) to assess inflammatory cell infiltration and goblet cells. The sections were photographed, and two researchers analyzed the photos, one of them blinded to treatment. Inflammation was quantified on five airways with intact epithelial lining and a minimum-to-maximum internal diameter ratio greater or equal to 0.5. An inflammation score for peribronchial and perivascular inflammation was used:12 0, 1, 2, 3, and 4, corresponding to none, mild, moderate, marked, or severe inflammation, respectively.13 The sum of scores defined total lung inflammation. A similar score was used for the quantification of goblet cells.
Statistical analysis
Results are expressed as means ± standard deviations or counts (proportions). Comparisons between groups were made with repeated-measure one-way or two-way ANOVA. A p-value of 0.05 or less was considered statistically significant. GraphPad Prism® 6 (LaJolla, CA) was used.
Results
Phytochemical characterization of M. charantia leaf extract
In MCHA, the following compounds were identified: 4- hydroxybenzoic acid, 4-O-glucoside, gentisic acid, esculetin, protocatechuic aldehyde, naringenin, (25ξ)-26-hydroxy-momordicoside L, momordicoside D, momordicoside L, momordicine II, momordicine I, karaviloside IV, and (23S*)-3β-hydroxy-7β,23-dimethoxycucurbita-5,24-dien-19-al (Table 1).
Table 1 Compounds identified in Momordica charantia leaves by liquid chromatography-electrospray ionization–Tandem mass spectrometry (LC-ESI-MS/MS).
| Peak | Compound | Rta (min) | MSb (m/z) obs. |
MSb (m/z) calc. |
MS/MSc Product ions |
Molecular formula | Error | Ref.d |
|---|---|---|---|---|---|---|---|---|
| 1 | 4-Hydroxybenzoic acid 4-O-glucoside |
3.07 | 299.0778 [M‒H]‒ | 299.0767 [M‒H]‒ | 299.0778→ 174.9566, 160.8427, 154.9743, 137.0246 | C13H16O8 | 3.68 [M‒H]‒ | Chanda et al., 2018 |
| 2 | Protocatechuic aldehyde | 6.54 | 137.0246 [M‒H]‒ | 137.0239 [M‒H]‒ | 137.0246 | C7H6O3 | 5.11 [M‒H]‒ | Chanda et al., 2018 |
| 3 | Naringenin | 9.33 | 271.0614 [M‒H]‒ 273.0765 [M+H]+ |
271.0606 [M‒H]‒ 273.0763 [M+H]+ |
271.0614→227.0711, 177.0194, 165.0196, 151.0040, 145.0301, 119.0507, 107.0143 273.0765→153.0183, 147.0442, 119.0482 |
C15H12O5 | –1.46 [M+H]+ 2.95 [M‒H]‒ |
Chanda et al., 2018 |
| 4 | (25ξ)-26-Hydroxy momordicoside L |
9.88 | 673.3932 [M+Na]+ | 673.3928 [M+Na]+ | 673.3932→493.3299 | C36H58O10 | 0.59 [M+Na]+ | Zhang et al., 2012 |
| 5 | Momordicoside D | 11.76 | 805.4721 [M+Na]+ | 805.4714 [M+Na]+ | 805.4721 | C42H70O13 | 0,87 [M+Na]+ | Ma et al. 2012 |
| 6 | Momordicoside L | 12.28 | 657.3983 [M+Na]+ | 657.3979 [M+Na]+ | 657.3983→477.3344, 203.0519 | C36H58O9 | 0.61 [M+Na]+ | Ma et al., 2012 |
| 7 | Momordicine II | 12.85 | 657.3987 [M+Na]+ | 657.3979 [M+Na]+ | 657.3987→479.3502, 203.0530 | C36H58O9 | 1.22 [M+Na]+ | Ma et al., 2012 |
| 8 | Momordicine I | 15.85 | 471.3481 [M‒H]‒ 495.3453 [M+Na]+ |
471.3474 [M‒H]‒ 495.3450 [M+Na]+ |
471.3481→430.9876, 306.9829 495.3453→477.3342 |
C30H48O4 | 1.49 [M‒H]‒ 0.61[M+Na]+ |
Ma et al., 2012 |
| 9 | Karaviloside IV | 15.78 | 649.4313 [M‒H]‒ | 649.4316 [M‒H]‒ | 649.4313→631.0042, 112.9859 | C37H62O9 | –0.46 [M‒H]‒ | Zhang et al., 2012 |
| 10 | (23S*)-3β-Hydroxy-7β, 23-dimethoxycucurbita-5, 24-dien-19-al | 19.22 | 523.3761 [M+Na]+ | 523.3763 [M+Na]+ | 523.3761 | C32H52O4 | –0.38 [M+Na]+ | Zhang et al., 2012 |
aRt, retention time; bMS, mass spectrometry; cMS/MS, Tandem mass spectrometry; dRef, reference.
M. charantia leaf extract decreases total cell and eosinophil counts in BAL
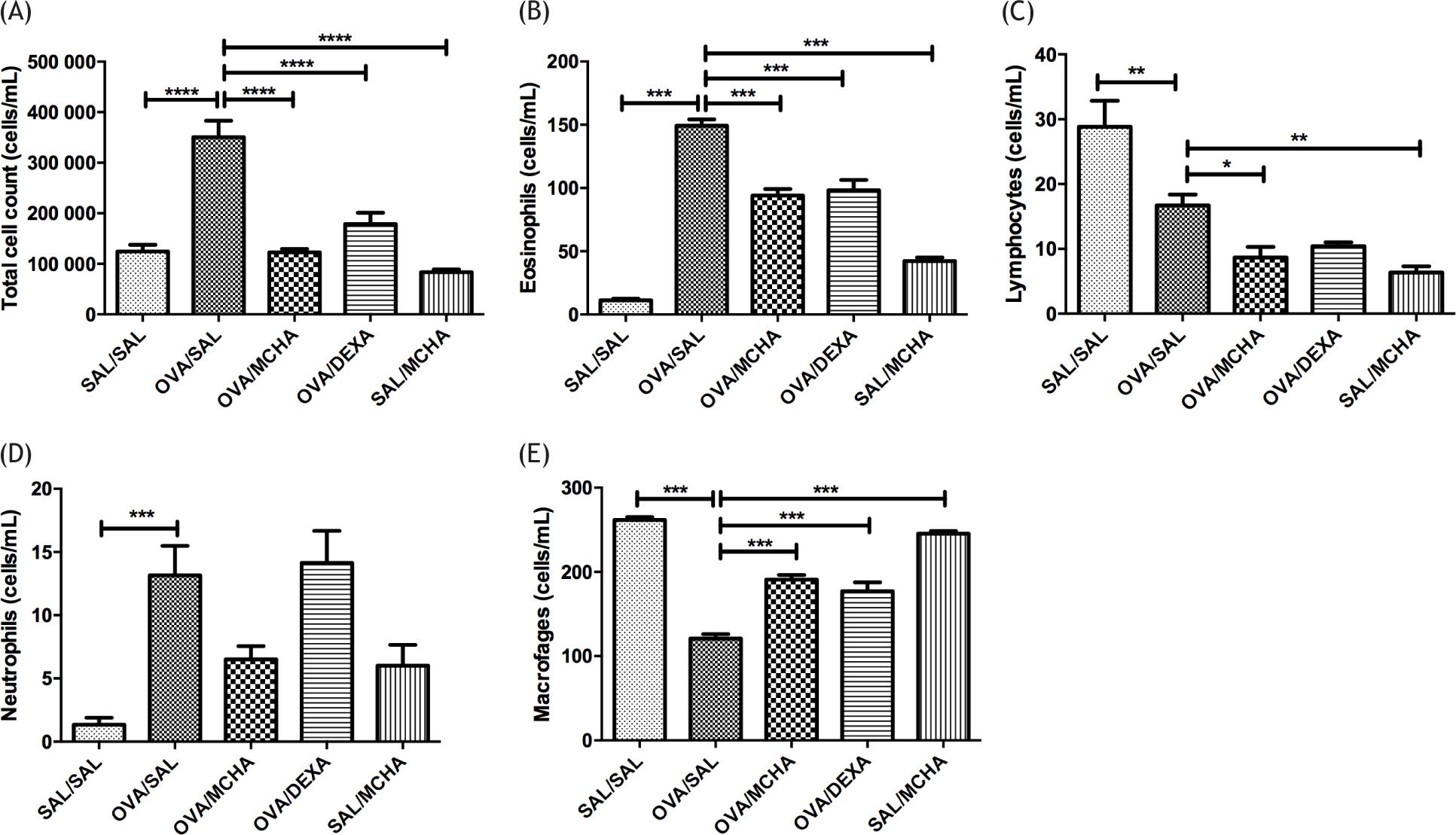
The total number of cells, eosinophils, and neutrophils were significantly higher in OVA-challenged mice compared to saline-challenged mice (p<0.001), while the numbers of lymphocytes and macrophages were significantly lower (Figure 1). Treatment with MCHA extract or dexamethasone significantly decreased the numbers of total cells and eosinophils (p<0.001) and increased macrophage counts (p<0.001) when compared to OVA-challenged mice. Of note, the effects of MCHA extract and dexamethasone on airway inflammation were similar.
Figure 1 Momordica charantia L. leaf hydroethanolic extract (MCHA) administration (500 mg/kg) reduced total cell (A), eosinophil (B), and lymphocyte (C) counts in bronchoalveolar lavage (BAL) fluid from ovalbumin (OVA)-challenged mice; whereas increased macrophages (E) counts. Values are shown as the mean and standard error of the mean (SEM). Legend: SAL, normal saline; DEXA, dexamethasone (2 mg/kg); *P<0.05; **P<0.01; ***P<0.001; ****P<0.0001, all compared with OVA-SAL group.
M. charantia leaf extract abolishes airway hyperresponsiveness
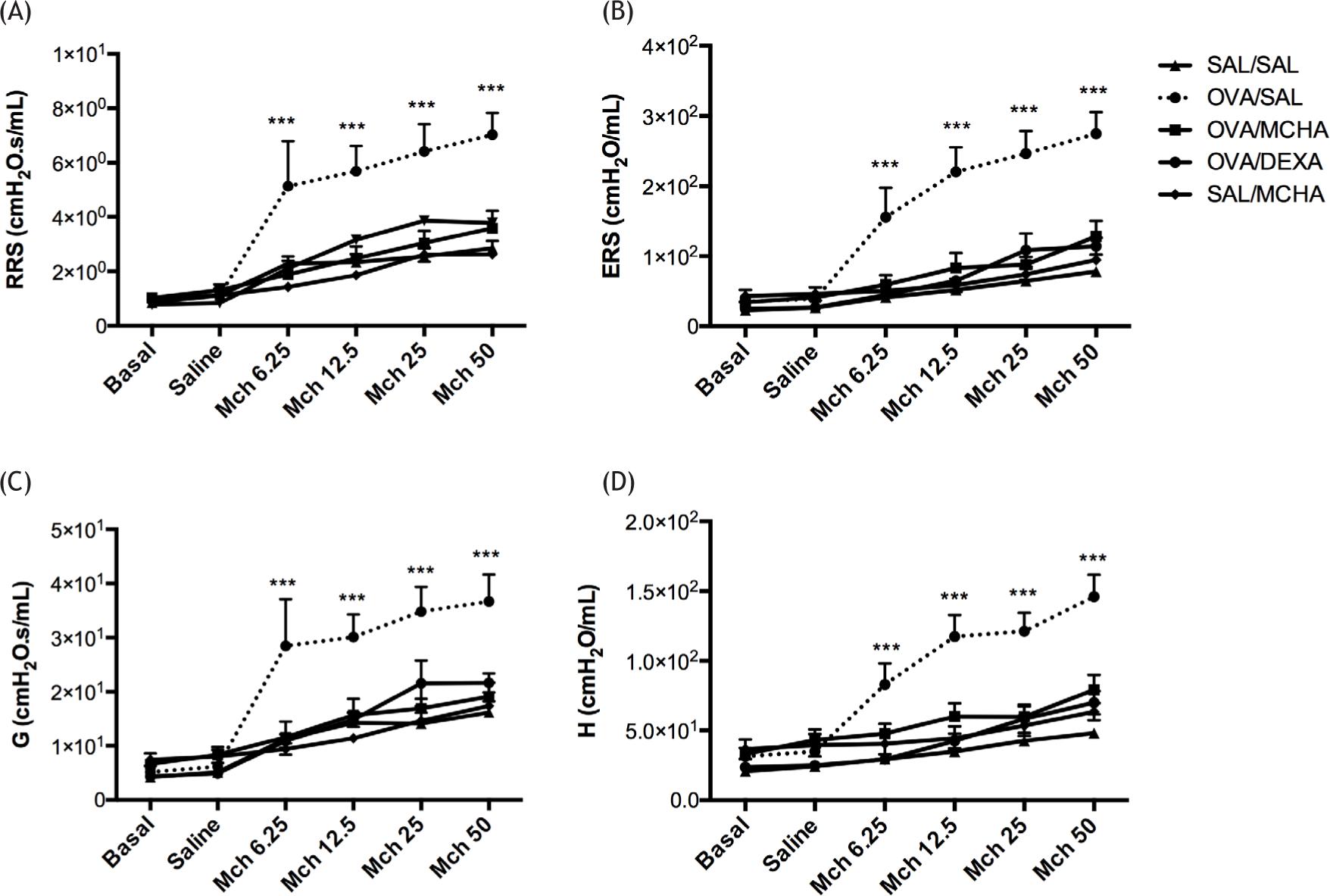
Lung mechanics is shown in Figure 2. Mice in the OVA-challenged group had a significant increase in airway hyperresponsiveness when compared to saline-challenged mice in all parameters RRS, ERS, G, and H (P<0.001). The administration of extract of M. charantia or dexamethasone significantly decreased airway hyperresponsiveness, measured by RRS (P<0.001), ERS (P<0.001), G (P<0.001), and H (P<0.001). Of note, MCHA extract and dexamethasone administration similarly improved airway hyperresponsiveness to basal levels (Figure 2).
Figure 2 Momordica charantia L. leaf hydroethanolic extract (MCHA) administration (500 mg/kg) reduced airway hyperresponsiveness in ovalbumin (OVA)-challenged mice. Airway responsiveness was measured with increasing concentration of aerosolized methacholine between 6.25 and 50 mg/mL, following sensitization and challenge. (A) Total resistance (RRS), (B) total elastance (ERS), (C) tissue resistance (G), and (D) tissue elastance (H). Values are shown as the mean and standard error of the mean (SEM). SAL, normal saline; DEXA, dexamethasone (2 mg/kg); ***all groups were significantly different (P<0.001) from OVA/SAL.
M. charantia leaf extract decreases IL-5 and IL-13 levels in lung homogenate
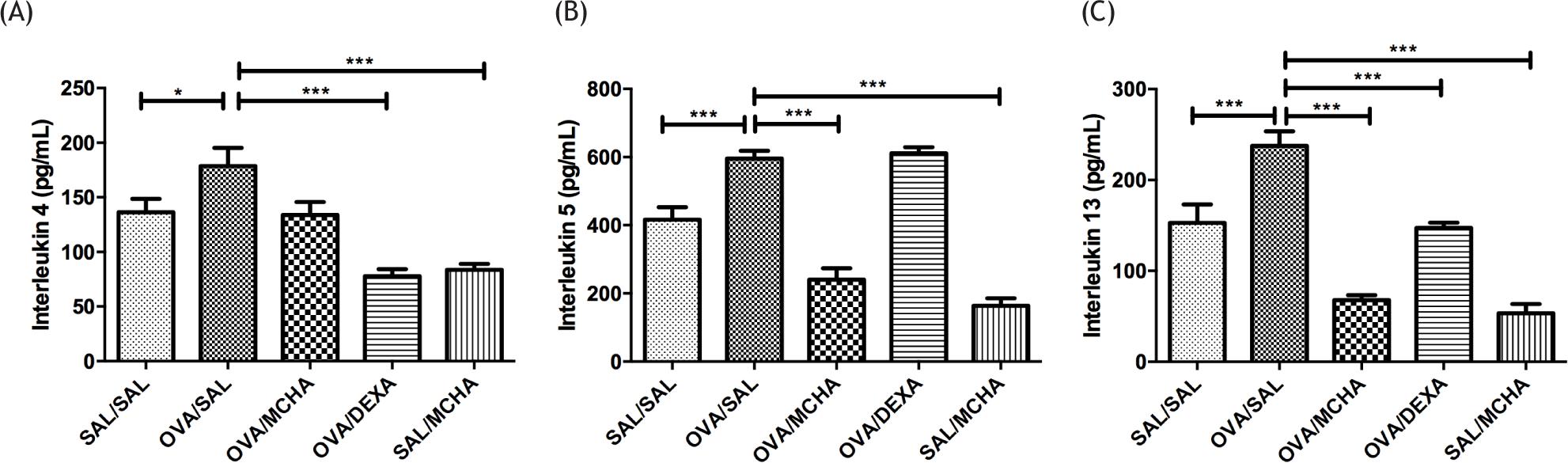
The concentrations of cytokines in lung homogenates are shown in Figure 3. After OVA challenges, IL-4, IL-5, and IL-13 were significantly higher than those of the control group. When compared to OVA-challenged mice, treatment with MCHA extract diminished considerably the levels of IL-5 and IL-13 (P<0.001), while dexamethasone decreased the levels of IL-4 and IL-13 (P<0.001).
Figure 3 Momordica charantia L. leaf hydroethanolic extract (MCHA) administration (500 mg/kg) reduced the concentrations of IL-5 (B) and IL-13 (C), but not IL-4 (A), in ovalbumin (OVA)-challenged mice. SAL normal saline; DEXA dexamethasone (2 mg/kg); *P<0.05; ***P<0.001, all compared with OVA-SAL group.
M. charantia leaf extract does not change total antioxidant capacity (TAC)
TAC levels were not significantly different among groups (data not shown).
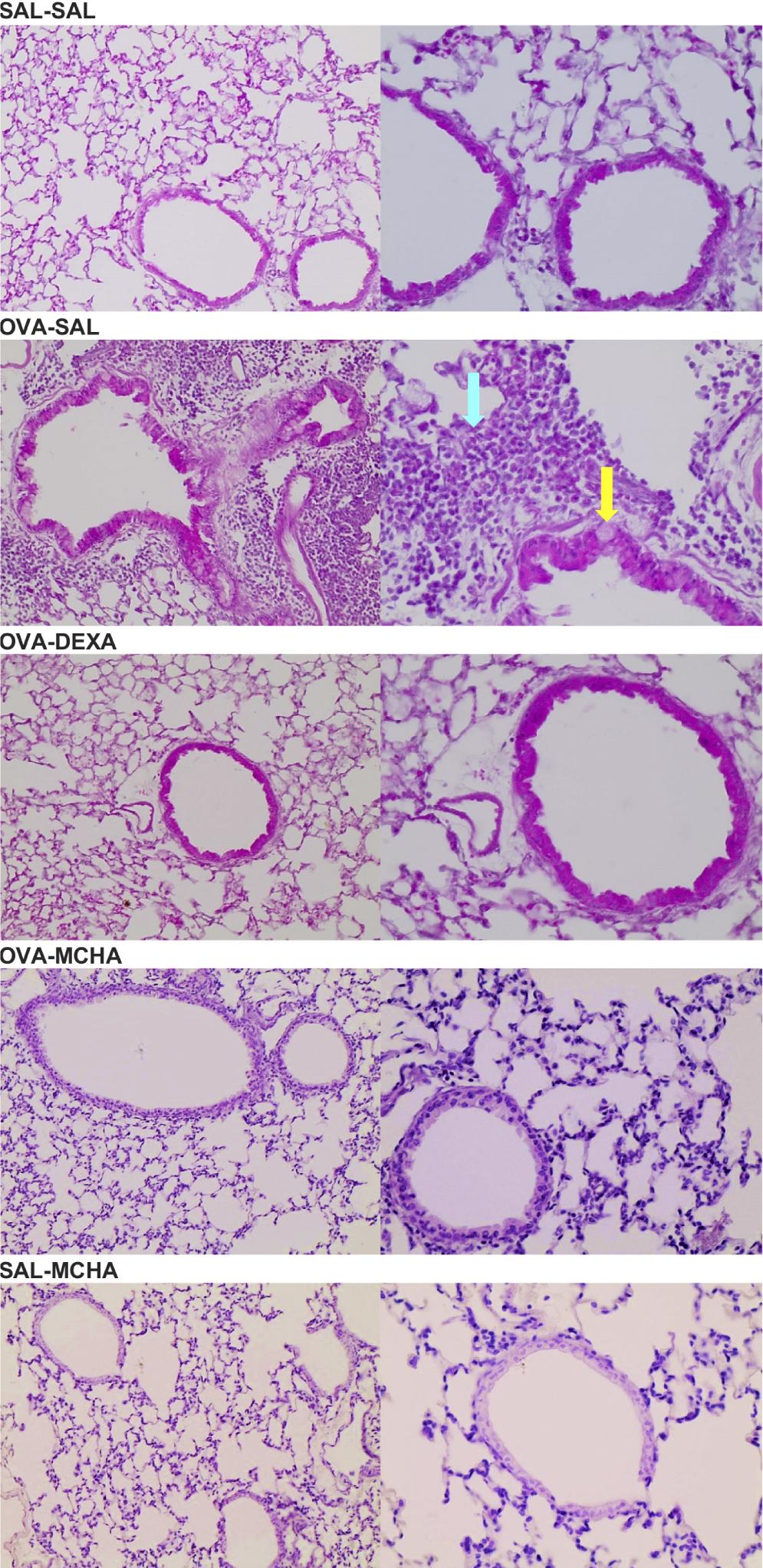
M. charantia leaf extract reduces lung inflammation and mucus production
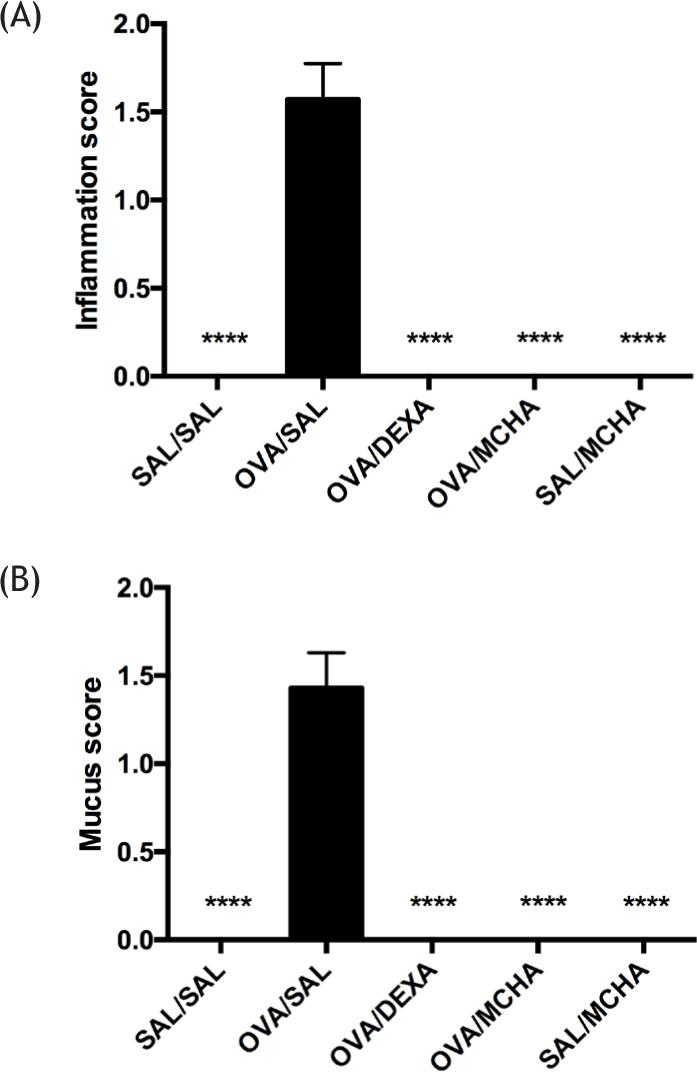
OVA-challenged mice exhibited a significantly higher number of inflammatory cells and mucus production compared to saline-challenged mice (P<0.0001). Peribronchial and perivascular inflammation and mucus production were significantly decreased with MCHA and dexamethasone treatments (Figure 4 and Figure 5).
Figure 4 Momordica charantia L. leaf hydroethanolic extract (MCHA) administration (500 mg/kg) suppressed inflammation (A) and mucus (B) scores in ovalbumin (OVA)-challenged mice. SAL, normal saline; DEXA, dexamethasone (2 mg/kg); ***, all groups were significantly different (P<0.001) from OVA/SAL.
Figure 5 Representative photomicrographs (H&E, ×20 or ×40) of lung tissue of mice challenged with ovalbumin (OVA) or saline solution (SAL) and treated with SAL or of Momordica charantia L. leaf hydroethanolic extract (MCHA) at 500 mg/kg or dexamethasone (DEXA) at 2 mg/kg. The yellow arrow indicates a goblet cell with mucus, and the cyan arrow points to an eosinophilic inflammatory infiltrate. These alterations were only seen in mice from group OVA/SAL.
Discussion
The present study showed, for the first time, that a hydroethanolic extract from M. charantia leaves (MCHA) is effective in treating asthma in an animal model. Treatment with MCHA attenuated bronchial hyperresponsiveness and lung and airway inflammation, demonstrated by less inflammatory cells in BAL, lower levels of IL-5 and IL-13 in lung homogenate, and absence of inflammation and mucus in lung tissue. Additionally, MCHA reduced the influx of inflammatory cells to the airway. Of note, these results were similar to those obtained with dexamethasone.
M. charantia is traditionally used for diabetes.14-20 The benefits of M. charantia for diabetic patients are attributed to its hypoglycemic and lipid-lowering properties and its effect on chronic inflammation.5 Indeed, the anti-inflammatory properties of M. charantia have been previously demonstrated in different animal models4,21 and knee osteoarthritis treatment.22
Regarding the anti-inflammatory mechanisms of M. charantia, previous studies demonstrated that it acts on several critical signaling pathways involved in inflammation, mainly through inhibition of the nuclear factor kappa-B (NF-κB).5 This mechanism is like that of corticosteroids and can decrease both Th1 and Th2 inflammatory responses. More specifically, M. charantia extracts (a) reduced tumor necrosis factor (TNF)-α production induced by LPS in vitro by decreasing the expression of inflammatory genes [IL-1α, IL-1β, and TNF-α]; (b) decreased NF-κB DNA binding activity and phosphorylation of p38, JNKs, ERKs, and MAPKs; (c) reduced NO and prostaglandin E2 production induced by LPS; (d) reduced the expression of cyclooxygenase (Cox)-2; and (e) reduced and activator protein-1 (AP-1) activity via downregulation of ERKs and Akt.5
In an animal model of sepsis, a diet enriched with M. charantia suppressed the inflammatory response, lowering the levels of inflammatory markers C-reactive protein (CRP), IL-1, IL-6, and TNF-α, and increasing levels of IL-10.23,24 IL-1, IL-6, and TNF-α are typical Th1 cytokines. In these studies, M. charantia also diminished NF-κB, inducible nitric oxide synthase (iNOS), and Cox-2 gene expression.23,24 In mice fed with high-fat diet, M. charantia decreased serum CRP, IL-6, and TNF-α and controlled hyperglycemia and hyperlipidemia. More recently, M. charantia extracts reduced intercellular adhesion molecule-1 (ICAM-1) expression and upregulated mir-221/-222 in TNF-α-treated lung tissues in mice, also decreasing PI3K/Akt/NF-κB/IκB.5
The corticosteroid-like anti-inflammatory effect of M. charantia can be attributed to its major chemical constituents: cucurbitane-type triterpenoids, cucurbitane-type triterpene glycosides, phenolic acids, flavonoids, essential oils, fatty acids, amino acids, sterols and saponin constituents, and some proteins.4 These isolated compounds also significantly attenuated lipopolysaccharide (LPS)-induced inflammation, down-regulating the expression of NF-κB, iNOS, and the pro-Th1-inflammatory markers IL-1α, IL-1β, IL-6, IL-1β, TNF-α, and Cox-2 in preclinical models.25–28
Another possible mechanism by which M. charantia extract inhibited the allergen-induced response is decreasing the levels of Th2 cytokines. In our study, the administration of MCHA significantly reduced IL-5 and IL-13 levels in lung homogenate. These cytokines are involved in the recruitment of eosinophils to the airways in asthma.29,30 This fact can explain the reduction of bronchial hyperresponsiveness observed with MCHA, which may be related to the decrease of total cells and eosinophils in BAL and lung tissue. The eosinophilic infiltrates in the lungs contribute to bronchoconstriction, mucus production, airway injury, increased hyper-responsiveness, and vascular permeability.31
Herbal medicines with anti-inflammatory properties, like M. charantia, are potential sources of new drugs. M. charantia is present in the national list of plants of interest for the Brazilian public health system, is found in all continents, and is easily grown and harvested. It is generally regarded as safe, although limited safety data is available.20 Therefore, further studies need to be encouraged.
One limitation of our study was the short treatment course that we used. Longer treatment courses could have resulted in more expressive results regarding inflammatory markers, such as IL-4. Another limitation is that we studied a single dose (500 mg/kg), and it is not clear whether smaller doses could be as effective. Future studies may determine the best dose.
Conclusions
The administration of a hydroethanolic extract from leaves of M. charantia L., at a dose of 500 mg/kg, decreased airway hyper-responsiveness and attenuated lung and airway inflammation in an ovalbumin-induced murine asthma model.
Authors’ Contributions
-
Clara do Prado Lopes Frota, Alessa Castro Ribeiro, Marco Túlio Alvarenga Calil Filho, Lucas Junqueira de Freitas Morel, and Vanessa Maciel Braulio da Fonseca were involved in the animal and lab experiments.
-
Leandra Naira Zambelli Ramalho was involved in histological analyses of lung specimens.
-
Jéssica Cristina Amaral and Silvia Helena Taleb Contini were involved with phytochemical analysis.
-
Marcos de Carvalho Borges and Fabio Carmona were involved in study conceptualization and design, statistical analysis, writing, and revising the final manuscript.
-
Ana Maria Soares Pereira was involved in study conceptualization and design, extract production, phytochemical analysis, statistical analysis, writing, and revising the final manuscript.
Conflict of interest
The authors declare no potential conflicts of interest with respect to research, authorship and/or publication of this article.
Funding
This work was supported by the National Council of Technological and Scientific Development (CNPq), grant # 473261/2013-8. This study was financed in part by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior - Brasil (CAPES) - Finance Code 001.
REFERENCES
1. Global Initiative for Asthma. Global strategy for asthma management and prevention. [cited
2. Cruz AA, Pizzichini ALGFE, Fiterman J, Jones LFFPMMMPM, Lima MA, Stirbulo PAMCR. Diretrizes da sociedade brasileira de pneumologia e tisiologia para o manejo da asma-2012. J Bras Pneumol Tisiol. 2012;38:S1-46.
3. The Plant List. [cited 2021]. Available from: http://www.theplantlist.org
4. Dandawate PR, Subramaniam D, Padhye SB, Anant S. Bitter melon: a panacea for inflammation and cancer. Chin J Nat Med. 2016;14(2):81–100. 10.1016/S1875-5364(16) 60002-X
5. Bortolotti M, Mercatelli D, Polito L. Momordica charantia, a nutraceutical approach for inflammatory related diseases. Front Pharmacol. 2019;10:1–9. 10.3389/fphar.2019.00486
6. Zhang J, Huang Y, Kikuchi T, Tokuda H, Suzuki N, Inafuku K, et al. Cucurbitane triterpenoids from the leaves of momordica charantia, and their cancer chemopreventive effects and cytotoxicities. Chem Biodivers. 2012;9(2):428–40. http://doi.wiley.com/10.1002/cbdv.201100142
7. Ma J, Krynitsky AJ, Grundel E, Rader JI. Quantitative determination of cucurbitane-type triterpenes and Triterpene Glycosides in dietary supplements containing bitter melon (Momordica charantia) by HPLC-MS/MS. J AOAC Int. 2012;95 (6):1597–1608. https://academic.oup.com/jaoac/article/95/6/1597-1608/5655273
8. Chanda J, Mukherjee PK, Biswas R, Malakar D, Pillai M. Study of pancreatic lipase inhibition kinetics and LC-QTOF-MS based identification of bioactive constituents of Momordica charantia fruits. Biomed Chromatogr. 2018;e4463. https://onlinelibrary.wiley.com/doi/abs/10.1002/bmc.4463
9. Azevedo BC, Morel LJF, Carmona F, Cunha TM, Contini SHT, Delprete PG, et al. Aqueous extracts from Uncaria tomentosa (Willd. ex Schult.) DC. reduce bronchial hyperresponsiveness and inflammation in a murine model of asthma. J Ethnopharmacol. 2018;218:76–89. http://linkinghub.elsevier.com/retrieve/pii/S0378874117334566
10. de Freitas Morel LJ, de Azevedo BC, Carmona F, Contini SHT, Teles AM, Ramalho FS, et al. A standardized methanol extract of Eclipta prostrata (L.) L. (Asteraceae) reduces bronchial hyperresponsiveness and production of Th2 cytokines in a murine model of asthma. J Ethnopharmacol. 2017;198:226–34. http://linkinghub.elsevier.com/retrieve/pii/S0378874116323121
11. Erel O. A novel automated direct measurement method for total antioxidant capacity using a new generation, more stable ABTS radical cation. Clin Biochem. 2004;37(4):277–85. https://pubmed.ncbi.nlm.nih.gov/15003729/
12. Sur S, Wild JS, Choudhury BK, Sur N, Alam R, Klinman DM. Long term prevention of allergic lung inflammation in a mouse model of asthma by CpG oligodeoxynucleotides. J Immunol. 1999;162(10):6284–93. https://www.jimmunol.org/content/162/10/6284.short
13. Wills-Karp M, Luyimbazi J, Xu X, Schofield B, Neben TY, Karp CL, et al. Interleukin-13: central mediator of allergic asthma. Science. 1998;282(5397):2258–61. https://www.science.org/doi/abs/10.1126/science.282.5397.2258
14. Krawinkel MB, Keding GB. Bitter gourd (Momordica charantia): a dietary approach to hyperglycemia. Nutr Rev. 2006;64:331–7. http://www.ncbi.nlm.nih.gov/pubmed/16910221
15. El Batran SAES, El-Gengaihi SE, El Shabrawy OA. Some toxicological studies of Momordica charantia L. on albino rats in normal and alloxan diabetic rats. J Ethnopharmacol. 2006;108(2):236–42. http://www.ncbi.nlm.nih.gov/pubmed/16815658
16. Yang SJ, Choi JM, Park SE, Rhee EJ, Lee WY, Oh KW, et al. Preventive effects of bitter melon (Momordica charantia) against insulin resistance and diabetes are associated with the inhibition of NF-κB and JNK pathways in high-fat-fed OLETF rats. J Nutr Biochem. 2015;26(3):234–40. http://www.sciencedirect.com/science/article/pii/S095528631400237X
17. Rahman IU, Khan RU, Rahman KU, Bashir M. Lower hypoglycemic but higher antiatherogenic effects of bitter melon than glibenclamide in type 2 diabetic patients. Nutr J. 2015;14(1): 13. http://ezproxy.think.edu.au/login?url=http://search.ebscohost.com/login.aspx?direct=true&db=mdc&AN=25504465&site=ehost-live&scope=site&custid=s8454151
18. Kim SK, Jung J, Jung JH, Yoon N, Kang SS, Roh GS, et al. Hypoglycemic efficacy and safety of Momordica charantia (bitter melon) in patients with type 2 diabetes mellitus. Complement Ther Med. 2020;52:102524. https://linkinghub.elsevier.com/retrieve/pii/S0965229920302491
19. Cortez-Navarrete M, Martínez-Abundis E, Pérez-Rubio KG, González-Ortiz M, Méndez-del Villar M. Momordica charantia administration improves insulin secretion in type 2 diabetes mellitus. J Med Food. 2018;21(7):672–77. http://www.liebertpub.com/doi/10.1089/jmf.2017.0114
20. Peter EL, Kasali FM, Deyno S, Mtewa A, Nagendrappa PB, Tolo CU, et al. Momordica charantia L. lowers elevated glycaemia in type 2 diabetes mellitus patients: systematic review and meta-analysis. J Ethnopharmacol. 2019;231:311–24. 10.1016/j.jep.2018.10.033
21. Jia S, Shen M, Zhang F, Xie J. Recent advances in momordica charantia: functional components and biological activities. Int J Mol Sci. 2017;18(12). 2555 https://www.mdpi.com/1422-0067/18/12/2555
22. Soo May L, Sanip Z, Ahmed Shokri A, Abdul Kadir A, Md Lazin MR. The effects of Momordica charantia (bitter melon) supplementation in patients with primary knee osteoarthritis: a single-blinded, randomized controlled trial. Complement Ther Clin Pract. 2018;32:181–6. https://linkinghub.elsevier.com/retrieve/pii/S1744388118301701
23. Liaw C, Huang H, Hsiao P, Zhang L, Lin Z, Hwang S, et al. 5β,19-Epoxycucurbitane Triterpenoids from Momordica charantia and their anti-inflammatory and cytotoxic activity. Planta Med. 2015;81(1):62–70. https://www.thieme-connect.com/products/ejournals/abstract/10.1055/s-0034-1383307
24. Chao C-Y, Sung P-J, Wang W-H, Kuo Y-H. Anti-inflammatory effect of momordica charantia in sepsis Mice. Molecules. 2014; 19:12777–88. http://www.mdpi.com/1420-3049/19/8/12777/
25. Shivanagoudra SR, Perera WH, Perez JL, Athrey G, Sun Y, Wu CS, et al. In vitro and in silico elucidation of antidiabetic and anti-inflammatory activities of bioactive compounds from Momordica charantia L. Bioorganic Med Chem. 2019;27(14):3097–109. 10.1016/j.bmc.2019.05.035
26. Kobori M, Nakayama H, Fukushima K, Ohnishi-Kameyama M, Ono H, Fukushima T, et al. Bitter gourd suppresses lipopolysaccharide-induced inflammatory responses. J Agric Food Chem. 2008;56(11):4004–11. http://www.ncbi.nlm.nih.gov/pubmed/18489106
27. Lii C-K, Chen H-W, Yun W-T, Liu K-L. Suppressive effects of wild bitter gourd (Momordica charantia Linn. var. abbreviata ser.) fruit extracts on inflammatory responses in RAW 264.7 macrophages. J Ethnopharmacol. 2009;122(2):227–3. http://www.ncbi.nlm.nih.gov/pubmed/19330915
28. Nhiem NX, Yen PH, Ngan NTT, Quang TH, Kiem P Van, Minh C Van, et al. Inhibition of nuclear transcription factor-κB and activation of peroxisome proliferator-activated receptors in HepG2 cells by cucurbitane-type triterpene glycosides from Momordica charantia. J Med Food. 2012;15(4):369–377. http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=3308713&tool=pmcentrez&rendertype=abstract
29. Hogan SP. Recent advances in eosinophil biology. Int Arch Allergy Immunol. 2007;143:3–14. https://www.karger.com/Article/Abstract/101398
30. van Rensen ELJ, Evertse CE, van Schadewijk WAAM, van Wijngaarden S, Ayre G, Mauad T, et al. Eosinophils in bronchial mucosa of asthmatics after allergen challenge: effect of anti-IgE treatment. Allergy. 2009;64(1):72–80. https://onlinelibrary.wiley.com/doi/abs/10.1111/j.1398-9995.2008.01881.x
31. Davoine F, Lacy P. Eosinophil cytokines, chemokines, and growth factors: emerging roles in immunity. Front Immunol. 2014;5:1–17. 10.3389/fimmu.2014.00570