
Download






ORIGINAL ARTICLE
Conditioned medium from the bone marrow mesenchymal stem cells modulates immune response via signal transduction and activator of transcription 6 signaling pathway in an allergic rhinitis mouse model
Wentao Zoua, Pei Zoub, Jiaxiong Zhanga, Xiaojing Caia, Xueying Maoc, Guangpeng Liub*
aDepartment of Otolaryngology, Shanghai Tenth People’s Hospital, School of Medicine, Tongji University, Shanghai, China
bDepartment of Plastic and Reconstructive Surgery, Shanghai Tenth People’s Hospital, School of Medicine, Tongji University, Shanghai, China
cDepartment of Otolaryngology, Shanghai Tenth People’s Hospital Chongming Branch, Shanghai, China
Abstract
Background Allergic rhinitis (AR) is a common immune disease of the nasal mucosa characterized with immunoglobulin E (IgE)-mediated allergic inflammation after exposure to allergens in susceptible population. Previous reports have demonstrated that the bone marrow mesenchymal stem cells (BMSCs) could reduce allergic inflammation. However, there is little knowledge about whether the culture supernatant of BMSCs (conditioned medium, CM) has similar anti- inflammatory potential in treating AR.
Objective The study aimed to evaluate the immunoregulatory effects of conditioned medium derived from BMSCs (BMSC-CM) on allergic inflammation in an AR mouse model.
Material and Methods The AR murine model was induced by repeated sensitization and challenges with ovalbumin (OVA). Subsequently the allergic symptoms of AR mice, cytokine levels, the histopathological features of the nasal mucosa and T helper 1 (Th1) : T helper 2 (Th2) cells ratio were evaluated.
Results Treatment with BMSC-CM was found as effective as BMSCs in reducing allergic symptoms and inhibiting eosinophilic infiltration in the nasal mucosa. After BMSC-CM or BMSCs administration, the OVA-specific IgE and interleukin 4 levels in serum decreased and interferon gamma level increased compared with AR mice treated with uncultured fresh medium. Flow cytometry analysis revealed a decrease in Th1:Th2 cells ratio after OVA-sensitization and the ratio was reversed by BMSC-CM and BMSCs treatments. Furthermore, the data revealed that BMSC-CM suppressed the production of signal transduction and activator of transcription 6 (STAT6) at messenger RNA and protein levels in the nasal mucosa.
Conclusion BMSC-CM could ameliorate allergic inflammation and regulate the balance of Th cells, and the underlying mechanism was closely related to STAT6 signaling pathway. The immunoregulatory effects of BMSCs could be achieved through paracrine function, and nasal dripping of BMSC-CM might be a novel approach for the treatment of AR.
Key words: allergic rhinitis, bone marrow mesenchymal stem cells, conditioned medium, immune regulation, signal transduction and activator of transcription 6
*Corresponding author: Guangpeng Liu, Department of Plastic and Reconstructive Surgery, Shanghai Tenth People’s Hospital, School of Medicine, Tongji University, Shanghai, China. Email address: Guangpeng Liu: [email protected]
Received 26 December 2021; Accepted 2 May 2022; Available online 1 July 2022
Copyright: Zou W, et al.
License: This open access article is licensed under Creative Commons Attribution 4.0 International (CC BY 4.0). http://creativecommons.org/licenses/by/4.0/
Introduction
Allergic rhinitis (AR) is a common chronic noninfectious inflammatory disorder characterized by allergic symptoms of the nasal mucosa, such as rhinorrhea, sneezing, itchiness, and nasal congestion. AR is caused by the immunoglobulin E (IgE)-mediated T helper 1 (Th1) and T helper 2 (Th2) (Th1–Th2) immune imbalance, among which the superiority of Th2 immune response is recognized as the key factor.1 Large numbers of immune cytokines play an important role in inducing AR, and many signaling pathways are related to this process. Signal transducers and activators of transcription (STAT) are a family of proteins involved in the regulation of gene transcription and the activation of many immune inflammatory mediators.2 Previous studies have reported that STAT6 signaling pathway, a subtype of STAT, participates in the pathogenesis of AR. Interleukin 4 (IL-4), one essential Th2-associated cytokine which leads to allergic inflammation, can regulate the differentiation of immature Th cells into Th2 cells through STAT6 signaling pathway.3,4 Furthermore, IL-4 inhibits Th1 cell responses by upregulating the transcription repressors of interferon gamma (IFN-γ), an important Th1-associated cytokine in balancing Th1:Th2 cells ratio.5 Therefore, recovering the Th1–Th2 immunological balance is required when considering treatment modalities of AR.
The bone marrow mesenchymal stem cells (BMSCs), multipotent progenitor cells derived from the bone marrow, have demonstrated their capabilities in various tissue repairs and regeneration.6,7 On the other hand, BMSCs have demonstrated potent immunomodulatory properties to mitigate allergic responses in airway conditions.8–10 For example, Zhao et al. reported that intravenous injection of BMSCs could effectively decrease allergic symptoms and reduce inflammatory parameters in an AR mouse model.11 However, the transplanted BMSCs usually have a short life span in vivo and their utilization rate is not high. Intravascular administration of BMSCs may even cause vascular occlusion.12,13
Many researchers have identified that paracrine components, including soluble factors and extracellular vesicles (EVs), secreted by BMSCs contribute to wound-healing and tissue-regeneration upon stem cell transplantation.14–16 The paracrine effects also include immunomodulatory and anti-inflammatory properties, demonstrating unique potential in suppressing immunological responses and treating immune diseases.13 Compared with conventional stem cell-based therapies, the cell-free application can not only maintain the therapeutic effects of stem cells but also overcome the above shortcomings. Transplantation of the conditioned medium (CM) from BMSCs, which contains paracrine components, may present considerable advantages over cells in terms of manufacturing, storage, safety, and potential of being a ready-to-access biological product.17–19
The aim of the present study was to explore whether conditioned medium derived from BMSCs (BMSC-CM) have immunomodulatory effects to alleviate allergic sympotoms in an AR mouse model. We hypothesized that BMSC-CM can reduce inflammatory response by regulating Th1–Th2 immune imbalance through STAT6 signaling pathway, and this regulatory efficacy is achieved through paracrine mechanism of BMSCs.
Materials and Methods
Preparation of BMSC-CM
BALB/c mice were purchased from National Rodent Laboratory Animal Resources (Shanghai, China). All animal care and experi-mental procedures were ethically approved by Animal Care and Use Committee of Shanghai Tenth People’s Hospital (approval number: SHDSYY-2020-1581). The government and international animal experiment guidelines were strictly abided. BMSCs were isolated from the bone marrow of 4-week-old male BALB/c mice and in vitro expanded as described previously.20 The characterization of BMSCs was confirmed by cell surface marker expression (CD19, CD45, CD90, and CD105) and multilineage (osteoblast and adipocyte) differentiation capabilities. Cells were expanded in low-glucose Dulbecco’s modified Eagle’s medium (LG-DMEM; Gibco, Grand Island, NY, US) containing 10% fetal bovine serum (FBS; HyClone, Logan, UT, US), 100-U/mL penicillin, and 100-mg/mL streptomycin (both from Sigma-Aldrich, St. Louis, MO, US). When BMSCs of passage 2 grew to nearly 80% confluence, the medium was discarded. Cells were washed thrice with phosphate-buffered saline (PBS) and further incubated in fresh LG-DMEM without serum. The culture supernatant was collected after 48 h, filtered through a 0.2-µm syringe filter, centrifuged at 3000 g for 15 min to remove cell debris, and stored at -20°C before use. Meanwhile, BMSCs were trypsinized and counted by trypan blue staining. After labeled with 2-µM CellTracker CM-Dil (Invitrogen, Carlsbad, CA, US), 1.0 × 106 BMSCs were resuspended in 0.2-mL PBS. In all, 10 mL of supernatant from the same amount of cells was concentrated to approximately 50-fold to yield 0.2 mL of BMSC-CM using ultrafiltration method.21 The protein concentration of BMSC-CM was calculated using Bicinchoninic Acid (BCA) assay (BCA protein assay kit PC0020; Solarbio, Beijing, China) according to the manufacturer’s instructions. LG-DMEM kept for 48 h without culturing cells was similarly concentrated and used as a control conditioned medium (con-CM).
AR mouse model and intervention
A total of 20 healthy male 10-week-old BALB/c mice were used with minor modifications to establish the AR model as described previously.22 Briefly, 15 mice were first sensitized intraperitoneally with 50 µg of ovalbumin (OVA; Sigma-Aldrich, St. Louis, MO, US) and 2.5 mg of aluminum hydroxide (Al(OH)3) in 200-µL PBS (n = 15) every other day for seven times. The mice were intranasally challenged from day 15 to day 19 by daily dropping with 100 µg of OVA in 50-µL PBS to stimulate AR symptoms. From day 20 to day 24, 15 AR mice were randomly divided into three groups for different treatments (5 mice in each group). BMSC-CM group: administrated intranasally with 0.2 mL of BMSC-CM; BMSCs group: transfused with 1.0 × 106 labeled BMSCs (0.2 mL) via the tail vein; and con-CM group: administrated intranasally with 0.2 mL of con-CM. Meanwhile, the other five mice sensitized and challenged with PBS (n = 5) were also administrated intranasally with 0.2 mL of BMSC-CM as a control group (Figure 1). The intervention was performed once a day for 5 consecutive days. After the final intervention on day 24, the frequencies of sneezing and nasal scratching for each mouse were measured during a 15-min period.
Figure 1 Analysis of surface markers and differentiation capacity of BMSCs. BMSCs were positive for (A) CD90 and (B) CD105, and negative for (C) CD19 and (D) CD45. Osteogenic differentiation of BMSCs was verified by the formation of (E) calcified nodule (Von Kossa staining, original magnification ×40). (F) Adipogenic differentiation was revealed by positive staining of Oil Red (original magnification ×200).
Determination of OVA-specific IgE, IL-4, and IFN-γ in serum
The mice were sacrificed and samples of the vein blood, nasal mucosa, and spleen were harvested 24 h after the final intervention (day 25). Part of the nasal mucosa was taken from the front of the inferior turbinate and the anterior segment of the nasal septum, and immediately placed in liquid nitrogen for evaluation of STAT6 signaling pathway. The posterior part of the mouse head was prepared for histopathological evaluation. The levels of OVA-specific IgE, IL-4, and IFN-γ in serum were determined by enzyme linked immunosorbent assay (ELISA; Yifeibiotech, Shanghai, China) according to the manufacturer’s instructions.
Histopathological analysis of nasal mucosa
The posterior part of the mouse head was fixed in 4% paraformaldehyde and decalcified in 10% ethylenedi-aminetetraacetic acid (EDTA; pH 7.0) for 6–8 weeks. After being embedded in paraffin, samples were cut into 4-µm cross sections and stained with hematoxylin and eosin (H&E) for histopathological evaluation. In order to assess the degree of allergic inflammation, two blinded observers calculated the number of eosinophils in the nasal submucosa. The results were expressed as the number of eosinophils around the basal lamina calculated under a light microscope (magnification ×200). Degree of allergic inflammation in the nasal mucosa was evaluated by observing epithelial injury, loss of cilia, gland hyperplasia, and vascular dilatation. The parameter of each aspect was evaluated semi-quantitatively (0 = no change, 1 = mild change, 2 = moderate change, and 3 = severe change; Table 1), and the inflammation score was calculated by cumulative scores and compared statistically as reported previously.23,24 In the BMSCs group, CM-Dil-labelled BMSCs were transplanted through the tail vein, and the specimens were analyzed by fluorescence microscope (Leica Microsystems, Wetzlar, Germany).
Table 1 Degree of allergic inflammation in the nasal mucosa.
| Epithelial injury | Loss of cilia | Gland hyperplasia | Vascular dilatation | |
|---|---|---|---|---|
| 0 (No change) |
Integral epithelial structure | No damage or shedding of the cilia | No glandular hyperplasia | No vasodilation |
| 1 (Mild change) |
Slight damage to epithelial cells, still continuous structure | Mild damage to cilia | Mild hyperplasia of mucous glands | Mild dilation of submucosal vessels |
| 2 (Moderate change) |
Moderate damage to epithelial cells, partial discontinuous structure | Damage to the cilia, visible shedding | Mucous gland hyperplasia with a small amount of mucus secretion | Moderate dilation to submucosal vessels with interstitial edema |
| 3 (Severe change) |
Obvious damage to epithelial cells, disordered structure | Massive shedding of the cilia | Obvious mucous gland hyperplasia with a large amount of mucus secretion | Obvious dilation of submucosal vessels with severe interstitial edema |
Flow cytometry for detecting Th1/Th2/Regulatory T (Treg) cells
The spleen tissue was dissociated, resuspended in cell washing solution, and the lymphocyte suspension was adjusted to 1×106 cells/mL. Single-cell suspensions of lymphocytes (1×106 cells/well in 24-well culture plates) were cultured in RPMI 1640 medium (Gibco), and restimulated with OVA (100 ng/mL). After 3 days of culture, flow cytometry analysis was performed to identify Th1/Th2/Treg cells in the mouse spleen. Th1 cells were identified as CD4+IFN-γ+ lymphocytes; Th2 cells were identified as CD4+IL-4+ lymphocytes; and Treg cells were identified as CD4+CD25+FoxP3+ lymphocytes.
Then fluorescein isothiocyanate (FITC) anti-mouse CD4 antibody (85-11-0041-81, eBioscience, San Diego, CA, US) was added and reacted at 4°C for 30 min in dark. After being fixed and permeabilized using a fix/perm solution (GAS-003; Caltag, Carlsbad, CA, US) according to the manufacturer’s instructions, the CD4+ T cells were stained with purified anti-phycoerythrin (PE) anti-mouse IL-4 antibody (85-12-7041-81, eBioscience) and PE anti-mouse IFN-γ antibody (85-12-7311-82, eBioscience), and incubated at 4°C for 30 min in dark; finally washed with pre-cold PBS to remove unbound antibody.
Treg cells were stained with FITC anti-mouse CD4 antibody and allophycocyanin (APC) anti-mouse CD25 antibody (85-17-0251-81, eBioscience) and fixed and permeabilized using a fix/perm solution. The cells were incubated with PE anti-mouse FoxP3 antibody (85-12-5773-82, eBioscience). Flow cytometric analysis was performed on an BD LSR II flow cytometry system (Becton, Dickinson and Company, Lake Franklin, NJ, US).
Messenger RNA (mRNA) and protein expression measurement for STAT6 signaling pathway
Quantitative real-time polymerase chain reaction (PCR) was performed to determine the expression level of STAT6 mRNA in the mouse nasal mucosa. The total RNA was isolated using TrizolTM reagent (Invitrogen) following the manufacturer’s instructions. Complementary DNA (cDNA) was synthesized using a cDNA synthesis kit (SuperRT cDNA Synthesis Kit; CoWin Biosciences, Jiangsu, China) and subjected to PCR assay (7300 Real-time PCR System; Applied Biosystems, Foster City, CA, US). The PCR primer sequences of mouse genes of STAT6 signaling pathway and glyceraldehyde 3-phosphate dehydrogenase (GAPDH) are listed in Table 2.
Table 2 Primers of STAT6 and GAPDH.
| Gene | Primer sequence |
|---|---|
| STAT6-F | GAGCTACTGGTCAGATCGGC |
| STAT6-R | CTCTGGAGTAGGAAGGGGCT |
| GAPDH-F | GGTGAAGGTCGGTGTGAACG |
| GAPDH-R | CTCGCTCCTGGAAGATGGTG |
For Western blot analysis, the nasal mucosa of each group was homogenized in protein lysis buffer (PBS containing 0.1% Triton X-100), and BCA protein assay kit (PC0020; Solarbio) was used to detect protein concentration quantitatively. Equal amount of protein was separated by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to nitrocellulose filter. The membranes were blocked with 1% bovine serum albumin (BSA), washed and then probed with a 1:1000 diluted anti-STAT6 antibody (ab32520; Abcam, Cambridge, UK) at room temperature. After three additional washings, the membranes were incubated with horseradish peroxidase– conjugated secondary antibody (1:5000, IRDye 800CW; LI-COR Biosciences, Lincoln, NE, US) for 1 h at room temperature. Immunoreactive bands were visualized by Odyssey CLx Imager Dual-color infrared imaging system (LI-COR Biosciences). GAPDH (BA2913; Boster Preston, CA, US) was detected as internal control.25
Statistical analysis
Data obtained from three independent experiments were expressed as mean ± standard error of mean (SEM). Statistical analysis was assessed by ANOVA using the SPSS software package version 19.0, with statistical significance set at P < 0.05.
Results
Immunophenotypic and differentiation characterization of BMSCs
The BMSC cultures were analyzed for expression of cell-surface markers. BMSCs were positive for CD90 and CD105 and negative for CD19 and CD45 (Figures 1A–D). In order to examine the multilineage capacity of BMSCs, cells were differentiated as osteogenic and adipogenic lineages using lineage-specific induction factors. Osteogenic differentiation of BMSCs was verified by the formation of calcified nodule (Von Kossa staining; Figure 1E). Adipogenic differentiation was revealed by positive staining of Oil Red (Figure 1F).
A total of 0.2-mL BMSC-CM was administered intranasally to each mouse; it was obtained from 1.0 × 106 BMSCs after concentration. The protein concentration of BMSC-CM was 1414.23 ± 336.07 µg/mL.
Treatment with BMSC-CM alleviated nasal allergic symptoms in AR mice
Mice were sensitized on days 1, 3, 5, 7, 9, 11, and 13 by intraperitoneal injection of OVA + Al(OH)3, and challenged by intranasal dropping with OVA from day 15 to day 19 (Figure 2A). Compared to the control group (mice sensitized and challenged with PBS), the frequency of both sneezing and nasal scratching was dramatically increased in OVA-induced mice, confirming the successful establishment of AR model. After different interventions for the next 5 days, the incidence of sneezing (Figure 2B) and nasal scratching (Figure 2C) was significantly decreased in both BMSC-CM and BMSCs groups compared to the con-CM group, indicating that both BMSCs and BMSC-CM treatments could relieve nasal symptoms in AR mice.
Figure 2 (A) Timeline of the AR mouse model and different intervention groups. The AR symptoms were evaluated on day 24. (B)Number of sneezing per 15 min for each group. (C) Number of nasal scratching per 15 min for each group (*P < 0.05; **P < 0.01; and ***P < 0.001). BMSC-CM group: administrated intranasally with BMSC-CM; BMSCs group: transfused with 1.0 × 106 BMSCs through the tail vein; con-CM group: administrated intranasally with con-CM; control group: mice sensitized and challenged with PBS, administrated intranasally with BMSC-CM.
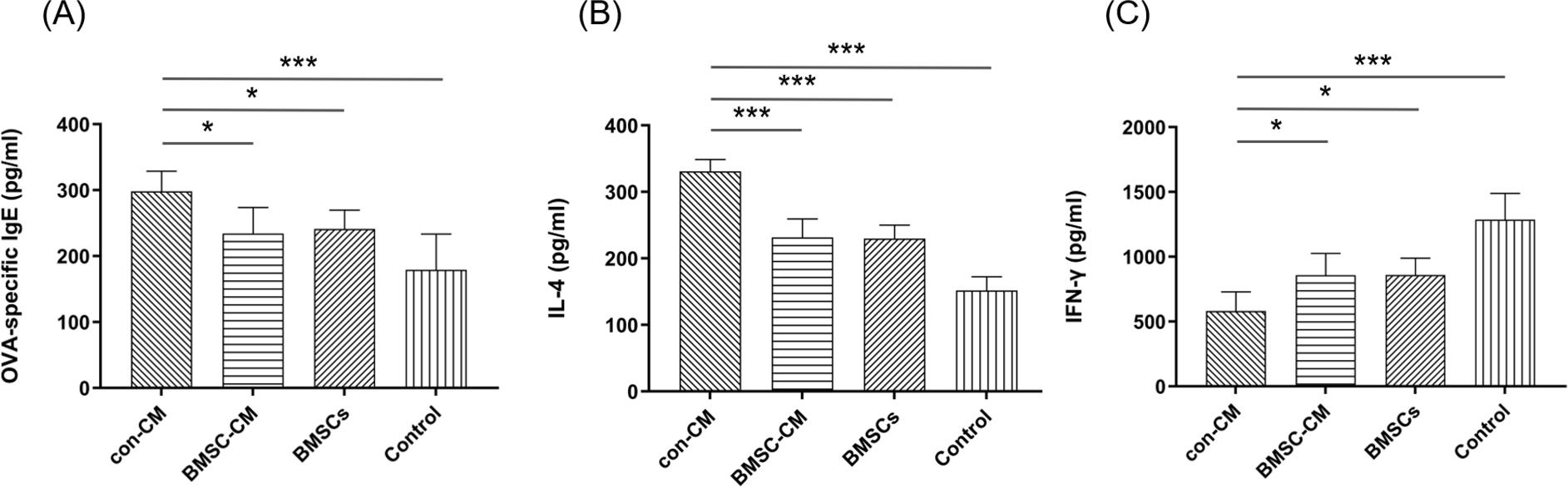
Detection of OVA-specific IgE, IL-4, and IFN-γ levels in mouse serum
According to ELISA measurement, the levels of OVA-specific IgE and IL-4 (Th2-associated cytokine) in murine serum were highest in the con-CM group, and decreased significantly after treatment with BMSC-CM or BMSCs (Figures 3A and B). Regarding the serum IFN-γ (Th1-associated cytokine) level, it was lower in mice sensitized with OVA than in the control group (Figure 3C). Statistically no difference was found between BMSC-CM and BMSCs groups, but both groups had higher values than values of the con-CM group. These results implied that BMSC-CM and BMSCs treatments restored Th1–Th2 immune balance in AR.
Figure 3 Detection of (A) OVA-specific IgE, (B) IL-4, and (C) IFN-γ levels in mouse serum. After treatment with BMSCs-CM or BMSCs, the levels of OVA-specific IgE and IL-4 decreased significantly compared to the con-CM group, with increased IFN-γ level (*P < 0.05; **P < 0.01; and ***P < 0.001).
Histological analysis of nasal mucosa
The cross section of murine nasal cavity was observed with H&E staining after sample decalcification. It was found that histological score of the nasal mucosa was higher in the OVA-induced groups than in the control group, and the extent of inflammatory cell infiltration in the submucosa tissue also increased accordingly (Figure 4A). Both eosinophil infiltration and inflammation scores were significantly increased in the nasal mucosa of AR mice. When treated with BMSC-CM or BMSCs, they decreased to over 50%, suggesting that both BMSC-CM and BMSCs treatments could reduce allergic inflammation of the nasal mucosa caused by OVA induction (Figures 4B and C).
Figure 4 (A) Histological observation of the nasal mucosa tissue. In the con-CM group, hematoxylin and eosin (H&E) staining revealed high number of eosinophil infiltration and increased epithelial injury, cilia damage, gland hyperplasia, and vascular dilatation compared to the control group. When treated with BMSC-CM or BMSCs, histological changes in the nasal mucosa improved significantly. (B) Numbers of eosinophils around the basal lamina were counted. (C) Inflammatory scores of each group were compared. (D) Migration of CM-Dil-labeled BMSCs (red marker) in the nasal mocosa was observed under fluorescence microscope (*P < 0.05; **P < 0.01; and ***P < 0.001).
For the BMSC-treated mice, 1.0 × 106 CM-Dil-labeled BMSC (0.2 mL) was transplanted through the tail vein everyday for 5 days. Cells with red fluorescence were detected under fluorescence microscope, confirming that labeled BMSCs migrated to the nasal mucosa with allergic inflammation (Figure 4D).
Changes of Th1/Th2/Treg cell proportions in the spleen
In order to further investigate the immunomodulatory effects of BMSC-CM on Th cell phenotype, changes in Th1/Th2/Treg cells in single-cell suspensions of the spleen lymphocytes (restimulated with OVA) were measured using flow cytometric analysis. The Th1:Th2 ratio was higher in the control group than in the AR groups after OVA sensitization, which established increased proportion of Th2 cells. The proportions of Th1 and Treg cells were significantly elevated whereas the proportion of Th2 cells decreased after treatment with BMSCs-CM or BMSCs. An increased Th1:Th2 ratio was observed in BMSC-CM and BMSCs groups compared to the con-CM group (Figures 5 and 6), indicating that both treatments were able to regulate the balance of Th cells and modulate the generation of Treg cells in AR mice.
Figure 5 Flow cytometric analysis of Th1/Th2 cell changes in single-cell suspensions of spleen lymphocytes (restimulated with OVA) for each group. An increased Th1:Th2 ratio was observed in BMSC-CM and BMSCs groups compared to the con-CM group, indicating that both treatments were able to regulate the balance of Th cells in AR mice. (A) Th1 (CD4+IFN-γ+) cells. (B) Th2 (CD4+IL-4+) cells (*P < 0.05; **P < 0.01; and ***P < 0.001).
Figure 6 Regulation of Treg cell (CD4+CD25+FoxP3+) proliferation by different treatments. After treatment with BMSCs-CM or BMSCs, the proportion of Treg cells was significantly elevated, indicating that both treatments could modulate the generation of Treg cells in AR mice. (A) Flow cytometric analysis. (B) Statistical comparison (*P < 0.05; **P < 0.01; and ***P < 0.001).
Changes of STAT6 signaling pathway in the nasal mucosa
In order to evaluate the role of STAT6 signaling pathway in BMSC-CM administration on allergic inflammation, changes in the expression of STAT6 mRNA and protein in the nasal mucosa were measured using real-time PCR and Western blot analysis. As shown in Figure 7, the highest expression levels of both STAT6 mRNA and protein were found in con-CM group, followed by BMSCs, BMSC-CM, and control groups, suggesting that conditioned medium derived from the stem cells could suppress expression of STAT6 signaling pathway under allergic inflammation conditions. No significant difference was found between BMSC-CM and BMSCs groups (P > 0.05).
Figure 7 Measurement of (A) STAT6 mRNA and (B and C) protein expression levels in the nasal mucosa (*P < 0.05; **P < 0.01; and ***P < 0.001). The data suggested that conditioned medium derived from the stem cells could suppress expression of STAT6 signaling pathway under allergic inflammation conditions.
Discussion
As a global health problem, AR has affected over 10–20% of adults and 20–40% of children globally.26 Present treatments, including medical drugs, immunotherapy, intranasal irrigation, and surgery, have specific limitations, such as high recurrence rate and adverse drug effects. Therefore, it is imperative to explore new treatment options for AR.
The BMSCs are multipotent adult stem cells derived from the bone marrow. In addtion to their capability of tissue regeneration and wound healing, BMSCs also demonstrate the effects of anti-inflammation and immunomodulation in treating airway allergic diseases, such as asthma and AR.27,28 However, transplantation of BMSCs is expensive and time-consuming, and the cells usually have a shorter in vivo life span. Recently, conditioned medium containing the paracrine factors secreted by BMSCs is reported to have immunomodulatory effects similar to the stem cells.29 In the present study, we found that BMSC-CM could modulate immune response and suppress Th2 cytokine production through STAT6 signal pathway in an AR murine model.
The immune imbalance of Th1–Th2 and immunodeficiency of Treg cells are the major factors of AR pathogenesis. Th2-associated cytokines, including IL-4, IL-5, and IL-13, are involved in delayed type-1 hypersensitivity mediated by IgE and contribute to the recruitment of eosinophil cells, which can start and maintain inflammation responses and further lead to a series of allergic symptoms.30–32 IL-4 is the main immunoregulatory cytokine secreted by activated Th2 cells in allergic reponses. It is also a vital cellular cytokine in the Janus kinase-signal transducer and activator of transcription (JAK/STAT) protein pathway. IL-4 is phosphorylated by binding to the JAK family receptors and initiates STAT phosphorylation. The IL-4 receptor site is remodeled and demethylated by the activation of STAT6 signaling pathway and antigen stimulation; it then promotes IgE production and enhances Th2 cells differentiation. On the contrary, IFN-γ, the principal Th1 effector cytokine, triggers the production of macrophages and inhibits Th2 cells proliferation. Th1 and Th2 cytokines are mutually antagonistic, and selective suppression of Th2 responses may alleviate allergic symptoms.33–35
In this study, we demonstrated the Th1–Th2 immune imbalance and Treg cell immunodeficiency in an AR mouse model, as confirmed by increased OVA-specific IgE, IL-4, and STAT6 levels in serum and eosinophil infiltration in the nasal mucosa. The OVA-specific IgE level was reduced and the symptoms of AR were alleviated in mice after treatment with BMSC-CM. Furthermore, the pathological observation of the nasal mucosa demonstrated that eosinophil infiltration, loss of cilia, and interstitial edema were ameliorated, indicating the ability of conditioned medium to suppress allergic airway inflammation. The detailed molecular mechanism was further identified by ELISA, flow cytometry, and proteomics analysis. Our results demonstrated that BMSC-CM could significantly reduce OVA-specific IgE, IL-4, and STAT6 formation and increase IFN-γ level in AR mice. By down-regulating the STAT6 signaling pathway, the over-activation of Th2 cells was inhibited whereas the proportion of Th1 cells was recovered.
Although many studies have reported the immunomodulatory effects of BMSCs in alleviating allergic symptoms, there are few reports about the immunosuppresive effects of BMSC-CM in treating AR. In this study, both BMSC-CM and BMSCs have demonstrated similar immunomodulation ability in reducing Th2 cytokine (IL-4) and increasing Th1 cytokine (IFN-γ) in an AR model. The culture supernatant of BMSCs, known as “conditioned medium,” contains a large number of bioactive components secreted by BMSCs, including soluble proteins, free nucleic acids, and EVs.36 It is speculated that BMSCs perform immunosuppressive effects on AR partly through the bioactive components produced by the paracrine pathway. Compared to the direct application of stem cells, BMSC-CM could be a promising cell-free option for AR treatment. In clinical practice, nasal dripping of conditioned medium is more effective and convenient than BMSC injection. Specifically, the cell-free therapy could avoid issues of immune compatibility and tumorigenicity associated with use of stem cells.
There are some limitations to this study. First, detailed mechanism of conditioned medium paracrine factors is complex and further in-depth experimental research is required. Second, as the BMSC-CM composition may change depending on culture conditions, standardization and isolation of conditioned medium collection is required. Still, we found that BMSC-CM had the immunoregulatory effects as effective as BMSCs in an AR mouse model, and could be used as a new option for AR therapy.37,38
Conclusion
We established in this study that treatment with BMSC-CM could reduce the production of IL-4, promote IFN-γ level, and regulate the ratio of Th1:Th2 cells through the STAT6 signaling pathway in an AR mouse model. BMSC-CM has many advantages over BMSCs therapy, including ease of preparing and storing, and convenience and safety of administration. Moreover, the immunoregulation of BMSCs is partly realized by paracrine function, representing a novel approach for the treatment of AR.
Acknowledgments
This work was supported financially by grants from the National Natural Science Foundation of China (31870974 and 51873101) and the research funding of “Action Plan for Sustainable Development of Science and Technology Innovation” of Chongming district, Shanghai, China (CKY2020-25).
Conflict of interest
The authors declared no potential conflict of interest.
Author contributions
Wentao Zou designed and carried out the experiments as well as wrote the first draft of the manuscript. Pei Zou collected and analyzed the data, and Guangpeng Liu interpreted the data and modified and confirmed the final version. Jiaxiong Zhang, Xiaojing Cai, and Xueying Mao also participated and carried out the experiment. All authors read and approved the final manuscript.
REFERENCES
1. Wheatley LM, Togias A. Clinical practice. Allergic rhinitis. N Engl J Med. 2015;372(5):456–63. 10.1056/nejmcp044141
2. Kuperman DA, Schleimer RP. Interleukin-4, interleukin-13, signal transducer and activator of transcription factor 6, and allergic asthma. Curr Mol Med. 2016;8(5):384–92. 10.2174/156652408785161032
3. Kim SH, Hong JH, Lee JE, Lee YC. 18β-Glycyrrhetinic acid, the major bioactive component of glycyrrhizae radix, attenuates airway inflammation by modulating Th2 cytokines, GATA-3, STAT6, and Foxp3 transcription factors in an asthmatic mouse model. Environ Toxicol Pharmacol. 2017;52:99–113. 10.1016/j.etap.2017.03.011
4. Seumois G, Zapardiel-Gonzalo J, White B, Singh D, Schulten V, Vijayanand P, et al. Transcriptional profiling of Th2 cells identifies pathogenic features associated with asthma. J Immunol. 2016;197(2):655–64. 10.4049/jimmunol.1600397
5. Li Y, Rui X, Ma B, Jiang F, Chen J. Early-life environmental factors, IFN-γ methylation patterns, and childhood allergic rhinitis. Int Arch Allergy Immunol. 2019;178(4):323–32. 10.1159/000495304
6. Hng CH, Camp E, Anderson P, Breen J, Zannettino A, Gronthos S. HOPX regulates bone marrow-derived mesenchymal stromal cell fate determination via suppression of adipogenic gene pathways. Sci Rep. 2020;10(1):11345. 10.1038/s41598-020-68261-2
7. Luby AO, Ranganathan K, Lynn JV, Buchman SR, Donneys A, Buchman SR. Stem cells for bone regeneration: Current state and future directions. J Craniofac Surg. 2019;30(3):730–5. 10.1097/SCS.0000000000005250
8. Dai YY, Ni SY, Ma K, Ma YS, Wang ZS, Zhao XL. Stem cells from human exfoliated deciduous teeth correct the immune imbalance of allergic rhinitis via Treg cells in vivo and in vitro. Stem Cell Res Ther. 2019;10(1):39. 10.1186/s13287-019-1134-z
9. Cho KS, Park HK, Park HY, Roh HJ, Jung JS, Roh HJ, et al. IFATS collection: Immunomodulatory effects of adipose tissue-derived stem cells in an allergic rhinitis mouse model. Stem Cells. 2009;27(1):259–65. 10.1634/stemcells.2008-0283
10. Ebrahim N, Mandour Y, Farid AS, Nafie E, Mohamed AZ, Refae A, et al. Adipose tissue-derived mesenchymal stem cell modulates the immune response of allergic rhinitis in a rat model. Int J Mol Sci. 2019;20(4):873. 10.3390/ijms20040873
11. Zhao N, Liu Y, Liang H, Jiang X. Bone marrow-derived mesenchymal stem cells reduce immune reaction in a mouse model of allergic rhinitis. Am J Transl Res. 2016;8(12):5628–36. eCollection 2016
12. Ankrum JA, Ong JF, Karp JM. Mesenchymal stem cells: Immune evasive, not immune privileged. Nat Biotechnol. 2014;32(3):252–60. 10.1038/nbt.2816
13. Abreu SC, Weiss DJ, Rocco PR. Extracellular vesicles derived from mesenchymal stromal cells: A therapeutic option in respiratory diseases? Stem Cell Res Ther. 2016;7(1):53. 10.1186/s13287-016-0317-0
14. Heo JS, Choi Y, Kim HS, Kim HO. Comparison of molecular profiles of human mesenchymal stem cells derived from bone marrow, umbilical cord blood, placenta and adipose tissue. Int J Mol Med. 2016;37(1):115–25. 10.3892/ijmm.2015.2413
15. Gesundheit B, Ashwood P, Keating A, Naor D, Melamed M, Rosenzweig JP. Therapeutic properties of mesenchymal stem cells for autism spectrum disorders. Med Hypotheses. 2015;84(3):169–77. 10.1016/j.mehy.2014.12.016
16. Basu J, Ludlow JW. Exosomes for repair, regeneration and rejuvenation. Expert Opin Biol Ther. 2016;16(4):489–506. 10.1517/14712598.2016.1131976
17. Goolaerts A, Pellan-Randrianarison N, Larghero J, Vanneaux V, Uzunhan Y, Clerici C, et al. Conditioned media from mesenchymal stromal cells restore sodium transport and preserve epithelial permeability in an in vitro model of acute alveolar injury. Am J Physiol Lung Cell Mol Physiol. 2014;306(11):975–85. 10.1152/ajplung.00242.2013
18. Park IS, Kim JH, Bae JS, Kim DK, Mo JH. The supernatant of tonsil-derived mesenchymal stem cell has antiallergic effects in allergic rhinitis mouse model. Mediators Inflamm. 2020;2020:6982438. 10.1155/2020/6982438
19. Nagata M, Iwasaki K, Akazawa K, Komaki M, Yokoyama N, Morita I, et al. Conditioned medium from periodontal ligament stem cells enhances periodontal regeneration. Tissue Eng A. 2017;23(9–10):367–77. 10.1089/ten.tea.2016.0274
20. Menicanin D, Bartold PM, Zannettino AC, Gronthos S. Identification of a common gene expression signature associated with immature clonal mesenchymal cell populations derived from bone marrow and dental tissues. Stem Cells Dev. 2010;19(10):1501–10. 10.1089/scd.2009.0492
21. Yu HS, Park MK, Kang SA, Cho KS, Mun SJ, Roh HJ. Culture supernatant of adipose stem cells can ameliorate allergic airway inflammation via recruitment of CD4(+) CD25(+) Foxp3 T cells. Stem Cell Res Ther. 2017;8(1):8. 10.1186/s13287-016-0462-5
22. Hellings PW, Hessel EM, Van Den Oord JJ, Kasran A, Van Hecke P, Ceuppens JL. Eosinophilic rhinitis accompanies the development of lower airway inflammation and hyper-reactivity in sensitized mice exposed to aerosolized allergen. Clin Exp Allergy. 2001;31:782–90. 10.1046/j.1365-2222.2001.01081.x
23. Isik S, Karaman M, Adan A, Kiray M, Bagriyanik H, Uzuner N, et al. Intraperitoneal mesenchymal stem cell administration ameliorates allergic rhinitis in the murine model. Eur Arch Otorhinolaryngol. 2017;274(1):197–207. 10.1007/s00405-016-4166-3
24. Sobkowiak P, Narozna B, Wojsyk-Banaszak I, Breborowicz A, Szczepankiewicz A. Expression of proteins associated with airway fibrosis differs between children with allergic asthma and allergic rhinitis. Int J Immunopathol Pharmacol. 2021;35: 2058738421990493. 10.1177/2058738421990493
25. Zhao C, Yu S, Li J, Ge R, Xu W, Ge R. Changes in IL-4 and IL-13 expression in allergic-rhinitis treated with hydrogen-rich saline in guinea-pig model. Allergol Immunopathol (Madr). 2017;45(4):350–5. 10.1016/j.aller.2016.10.007
26. Cheng L, Chen J, Fu Q, He S, Li H, Liu Z, et al. Chinese society of allergy guidelines for diagnosis and treatment of allergic rhinitis. Allergy Asthma Immunol Res. 2018;10(4):300–53. 10.4168/aair.2018.10.4.300
27. Marinas-Pardo L, Mirones I, Amor-Carro O, Fraga-Iriso R, Lema-Costa B, Ramos-Barbon D, et al. Mesenchymal stem cells regulate airway contractile tissue remodeling in murine experimental asthma. Allergy. 2014;69(6):730–40. 10.1111/all.12392
28. Li C, Fu Y, Wang Y, Kong Y, Li M, Wang Y, et al. Mesenchymal stromal cells ameliorate acute allergic rhinitis in rat. Cell Biochem Funct. 2017;35(7):420–5. 10.1002/cbf.3291
29. Borg ZD, Goodwin M, Sokocevic D, Wagner DE, Weiss DJ, et al. Systemic administration of human bone marrow-derived mesenchymal stromal cell extracellular vesicles ameliorates aspergillus hyphal extract-induced allergic airway inflammation in immunocompetent mice. Stem Cells Transl Med. 2015;4(11):1302–16. 10.5966/sctm.2014-0280
30. Small P, Keith PK, Kim H. Allergic rhinitis. Allergy Asthma Clin Immunol. 2018;14(Suppl 2):51. 10.1186/s13223-018-0280-7
31. Michaud B, Gouvis-Echraghi R, Candon S, Couderc R, Jais JP, Just J, et al. Quantification of circulating house dust mite-specific IL-4-and IL-13-secreting T cells correlates with rhinitis severity in asthmatic children and varies with the seasons. Clin Exp Allergy. 2014;44(2):222–30. 10.1111/cea.12222
32. Folci M, Ramponi G, Arcari I, Zumbo A, Brunetta E. Eosinophils as major player in type 2 inflammation: Autoimmunity and beyond. Adv Exp Med Biol. 2021;1347:197–219 10.1007/5584_2021_640
33. McCormick SM, Heller NM. Commentary: IL-4 and IL-13 receptors and signaling. Cytokine. 2015;75(1):38–50. 10.1016/j.cyto.2015.05.023
34. Howell MD, Fitzsimons C, Smith PA. JAK/STAT inhibitors and other small molecule cytokine antagonists for the treatment of allergic disease. Ann Allergy Asthma Immunol. 2018;120(4):367–75. 10.1016/j.anai.2018.02.012
35. Antczak A, Domańska-Senderowska D, Górski P, Pastuszak-Lewandoska D, Nielepkowicz-Gozdzinska A, Brzezianska-Lasota E, et al. Analysis of changes in expression of IL-4/IL-13/STAT6 pathway and correlation with the selected clinical parameters in patients with atopic asthma. Int J Immunopathol Pharmacol. 2016;29(2):195–204. 10.1177/0394632015623794
36. Börger V, Bremer M, Ferrer-Tur R, Gockeln L, Stambouli O, Giebel B, et al. Mesenchymal stem/stromal cell-derived extracellular vesicles and their potential as novel immunomodulatory therapeutic agents. Int J Mol Sci. 2017;18(7):1450. 10.3390/ijms18071450
37. Haddad R, Saldanha-Araujo F. Mechanisms of T-cell immuno-suppression by mesenchymal stromal cells: What do we know so far? Biomed Res Int. 2014;(2):216806. 10.1155/2014/216806
38. Yu QN, Guo YB, Li X, Li CL, Tan WP, Fu QL, et al. ILC2 frequency and activity are inhibited by glucocorticoid treatment via STAT pathway in patients with asthma. Allergy. 2018;73(9):1860–70. 10.1111/all.13438