
Download
ORIGINAL ARTICLE
Dexmedetomidine reduces dextran sulfate sodium (DSS)-induced NCM460 cell inflammation and barrier damage by inhibiting RhoA/ROCK signaling pathway
Xin Hana, Yini Wua, Yongwei Zhuangb, Jianmei Zhangb*
aDepartment of Anesthesiology, Lishui City People’s Hospital, the Sixth Affiliated Hospital of Wenzhou Medical University, the First Affiliated Hospital of Lishui University, Zhejiang Province, Lishui, China
bDepartment of Gastroenterology, Lishui City People’s Hospital, the Sixth Affiliated Hospital of Wenzhou Medical University, the First Affiliated Hospital of Lishui University, Zhejiang Province, Lishui, China
Abstract
Objective This study investigated the role of dexmedetomidine (DEX) in dextran sulfate sodium (DSS)-induced NCM460 cells.
Material and Methods The viability and apoptosis of NCM460 cells treated with DEX with or without DSS were detected by CCK-8 and terminal deoxynucleotidyl transferase (TdT) dUTP nick-end labeling (TUNEL) assay. The level of inflammatory factors and expression of inflammation-related proteins, tight junction proteins and Ras homolog gene family, member A/Rho-associated coiled-coil containing protein kinase (RhoA/ROCK) signaling-related proteins in NCM460 cells treated with DEX and/or U46619 (RhoA/ROCK agonist) and/or DSS were detected by the respective enzyme-linked immunosorbent assay (ELISA) kits and Western blot analysis. The permeability of NCM460 monolayers was examined with transepithelial electrical resistance (TEER) assay.
Results DEX had no effect on NCM460 cell viability. However, DEX improved the viability and barrier damage and suppressed the apoptosis and inflammation of DSS-induced NCM460 cells. Correspondingly, the expression of inflammation-related proteins was reduced and the expression of tight junction proteins was increased in DSS-induced NCM460 cells after treatment with DEX. In addition, RhoA/ROCK signaling was activated in NCM460 cells induced by DSS, which was suppressed by DEX. The protective effects of DEX on DSS-indued NCM460 cells were reversed by U46619.
Conclusion DEX improved viability and barrier damage while suppressed apoptosis and inflammation in DSS-indued NCM460 cells by inhibiting RhoA/ROCK signaling pathway.
Key words: dexmedetomidine, inflammation, barrier damage, RhoA/ROCK signaling pathway, ulcerative colitis
*Corresponding author: Jianmei Zhang, Department of Gastroenterology, Lishui City People’s Hospital, the Sixth Affiliated Hospital of Wenzhou Medical University, the First Affiliated Hospital of Lishui University, No. 15, Dazhong Street, Liandu District, Zhejiang Province, Lishui 323000, China. Email address: [email protected]
Received 24 December 2021; Accepted 15 February 2022; Available online 1 May 2022
Copyright: Han X, et al.
License: This open access article is licensed under Creative Commons Attribution 4.0 International (CC BY 4.0). http://creativecommons.org/licenses/by/4.0/
Introduction
Ulcerative colitis (UC) is a frequent inflammatory bowel disease that results in inflammatory response of the colon. Disruption in intestinal epithelial barrier and continued production of pro-inflammatory cytokines play a key role in the course of ulcerative colitis.1 The prevalence and morbidity of ulcerative colitis is highest in western developed countries.2 Meanwhile, the morbidity rate of ulcerative colitis has also elevated among Asians and Latin Americans.3 At present, corticosteroid drugs or immunosuppressants are primarily used for symptomatic treatment of inflammatory reactions. Although there are certain curative effects, long-term toxicity and adverse reactions are significant. The increased risk of cancers in patients with ulcerative colitis, which is easy to recur and has a prolonged disease course of 10–20 years, is as high as 28%.4
Dexmedetomidine (DEX) is a highly selective α2-adrenergic receptor agonist possessing anti-anxiety, sedative, sympathetic and analgesic effects, and widely used in intensive care units as an adjunct to sedation and anesthesia (Figure 1A).5 DEX preconditioning effectively prevents intestinal ischemia-reperfusion (I/R) injury and inhibits TLR4/MyD88/nuclear factor kappa B (NF-κB) signaling. Also, DEX could reduce the in vivo and in vitro production of inflammatory cytokines.6 In addition, DEX alleviates intestinal epithelial hyperpermeability caused by burns by reducing inflammation and enhancing the expression and distribution of tight junction proteins zonula occluden-1 (ZO-1) and occludin.7 In addition, DEX has a protective effect on cecum ligation and puncture (CLP)-induced intestinal injury, which may be related to the inhibition of inflammation by regulating TLR4 pathway(.8 However, the function of DEX in ulcerative colitis has not been determined.
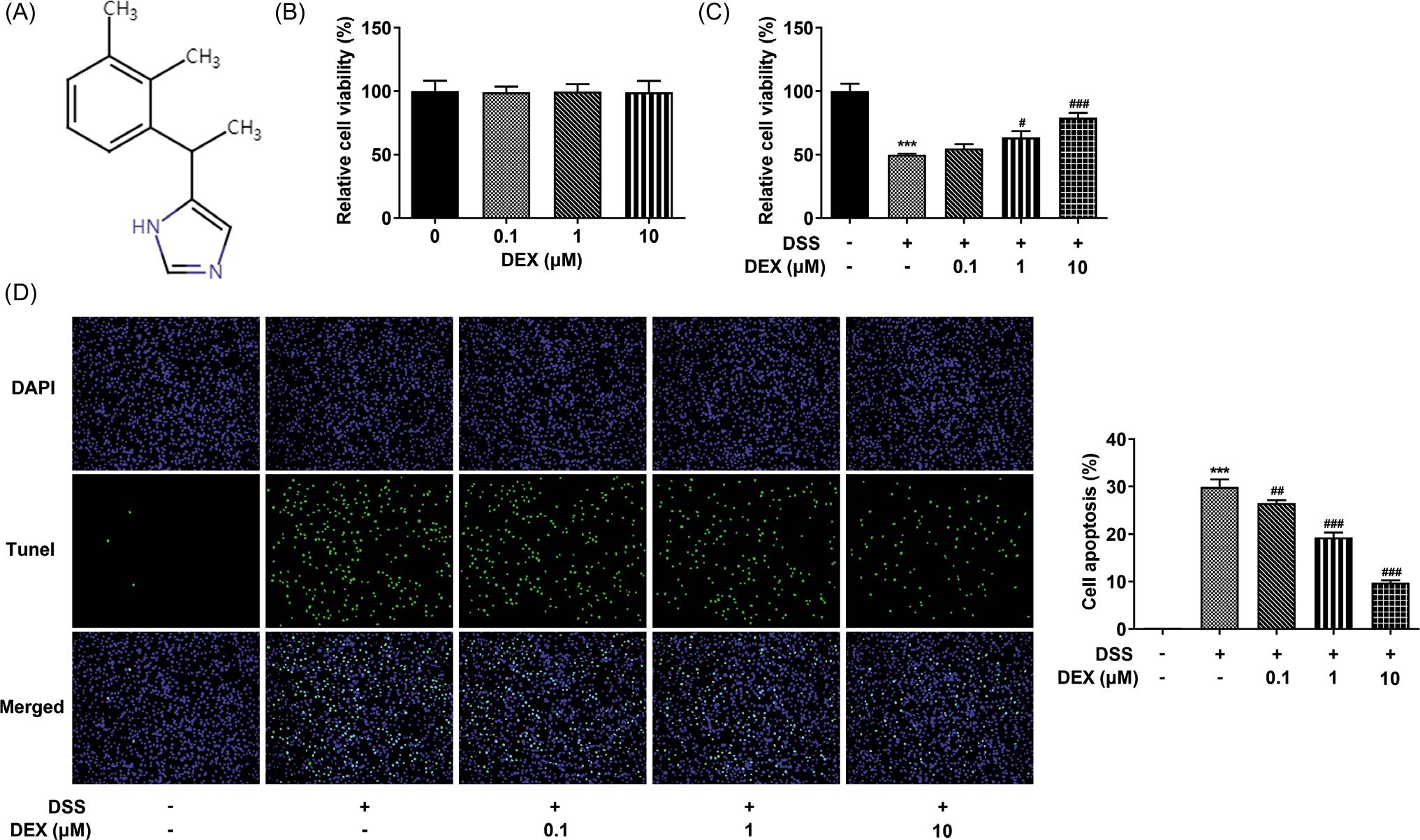
Figure 1 DEX increases viability and inhibits apoptosis of DSS-induced NCM460 cells. (A) Chemical structural formula of DEX. (B) Influence of DEX on the viability of NCM460 cells was appraised by CCK-8 assay. (C) Viability of DSS-induced NCM460 cells treated with DEX was detected by CCK-8 assay. (D) The apoptosis of DSS-induced NCM460 cells treated with DEX was determined by TUNEL assay. ***P < 0.001 vs. control group; #P < 0.05, ##P < 0.01 and ###P < 0.001 vs. DSS group.
Ras homolog gene family, member A/Rho-associated coiled-coil containing protein kinase (RhoA/ ROCK) pathway is closely related to a variety of cell functions, such as generation of reactive oxygen species (ROS), cell migration and differentiation etc.9 ROCK belongs to a 160-kD threonine/serine kinase, primarily containing ROCK1 and ROCK2. ROCK1 and ROCK2 are key effectors of RhoA and regulate a variety of physiological functions through phosphorylation of downstream targets.10–12 In the downstream of ROCK, the activation of NF-κB, a pivotal driving factor in the aetiology of inflammatory bowel disease (IBD), is closely related to colon and intestinal inflammation.13–15 DEX may reduce myocardial apoptosis and improve cardiac function by inhibiting RhoA/ROCK signaling pathway.16
Therefore, this paper is aimed to understand the definite role of DEX in dextran sulfate sodium (DSS)-triggered inflammation and barrier damage in NCM460 cells through RhoA/ROCK signaling pathway.
Materials and methods
Cell culture
The human normal colonic epithelial cell line NCM460 acquired from BeNa Culture Collection (BNCC, Beijing, China) were maintained in modified 1640 medium (HyClone, Logan, USA) supplemented with 10% fetal bovine serum (FBS; Gibco, CA, USA) and 1% antibiotics (Beyotime, Shanghai, China) at 37°C with 5% CO2.
DSS-treated NCM460 cells were used as an in vitro model for colitis. DSS 2% (MP Biomedicals, Southern California, USA) was added to the culture medium and cells were cultured for 24 h till the cell density reached about 70%. Additionally, DEX (0.1-, 1- and 10-μM) was used to pretreat NCM460 cells for 1 h.
Cell Counting Kit-8 (CCK-8) assay
In short, NCM460 cells grown in 96-well plates (5.5 × 103 cells/well) were pretreated with DEX for 1 h and 5% CO2 at 37°C. Treated or untreated cells in each well were cultivated with 10-μL CCK8 reagent for 2 h at 37°C. The absorbance was estimated at 450 nm by a microplate reader (Thermo Fisher Multiskan FC, Waltham, MA, USA).
TUNEL assay
After treated with DEX for 1 h and induced by DSS for 24 h, treated NCM460 cells were observed using a fluorescence microscope. This was done after 4',6-diamidino-2-phenylindole (DAPI) staining using a terminal deoxynucleotidyl transferase (TdT)-mediated deoxyuridine triphosphate (dUTP) in situ nick-end labeling (TUNEL) detection kit (Roche, Shanghai, China).
Enzyme linked immunosorbent assay (ELISA)
NCM460 cells were plated into 96-well plates (5.5 × 103 cells/well). In one experiment, NCM460 cells were pretreated with DEX (0.1-, 1- and 10-μM) for 1 h and then induced by DSS for another 24 h. In another experiment 1-nM U46619 (RhoA/ROCK agonist) and 10-μM DEX were used to pretreat NCM460 cells for 1 h and then these cells were induced by DSS for another 24 h. Finally, the cell culture media were harvested and tumor necrosis factor-α (TNF-α), interleukin-1β (IL-1β) and IL-6 levels were respectively tested using TNF-α ELISA kit (PT518), IL-1β ELISA kit (PI305) and IL-6 ELISA kit (PI330) (all from Beyotime).
Western blot analysis
With the adoption of radioimmunoprecipitation assay (RIPA) lysis buffer (Cell Signaling Technologies, Beverly, USA), treated NCM460 cells were subjected to protein isolation at 4°C for 20 min, followed by centrifugation (10,000 × g) for 10 min at 4°C. The bicinchoninic acid (BCA) assay measured protein concentration in supernatants. Subsequently, 30-µg total protein from each group was detached by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to polyvinylidene fluoride (PVDF) membranes. After impeded with 5% non-fat milk at room temperature for 1 h, the membranes were incubated at 4°C overnight with the following specific antibodies against cyclooxygenase-2 (COX-2), inducible nitric oxide synthase (iNOS), ZO-1, occludin, claudin-1, RhoA, ROCK1, ROCK2 and glyceraldehyde 3-phosphate dehydrogenase (GAPDH). Next day, the membranes were washed thrice and incubated with horseradish peroxidase (HRP)-conjugated goat-anti-rabbit secondary antibody at room temperature for 1 h. Subsequently, the membranes were visualised using a hemiluminescence kit (Millipore, Billerica, MA, USA) and Image-Pro Plus software (v6; Media Cybernetics Inc., Rockville, MD, USA) was used to analyze grey values.
Determination of transepithelial electrical resistance (TEER)
TEER assay was used to assess the permeability of NCM460 monolayers.17 NCM460 cells inoculated in 24-well transwell filters were allowed to grow until they formed a tight layer in an incubator at 37°C, 5% CO2 and saturated humidity. After indicated treatment, the TEER value was estimated by an epithelial voltohmmeter (World Precision Instruments, Sarasota, FL, USA).
Statistical analysis
Each experiment was independently performed in triplicate, and all data were displayed as mean ± standard deviation (SD). Multiple comparisons were evaluated by one-way analysis of variance followed by Tukey’s test. Differences between two groups were determined by t-test; P < 0.05 implied statistically significant.
Results
DEX increases viability and inhibits apoptosis of DSS-induced NCM460 cells
It was observed that the viability of NCM460 cells was not changed after administrated with 0.1-, 1- and 10-μM DEX (Figure 1B). The viability was inhibited and apoptosis was promoted in NCM460 cells after DSS induction; this all was reversed by DEX (Figures 1C and D).
DEX suppresses inflammatory response in DSS-induced NCM460 cells
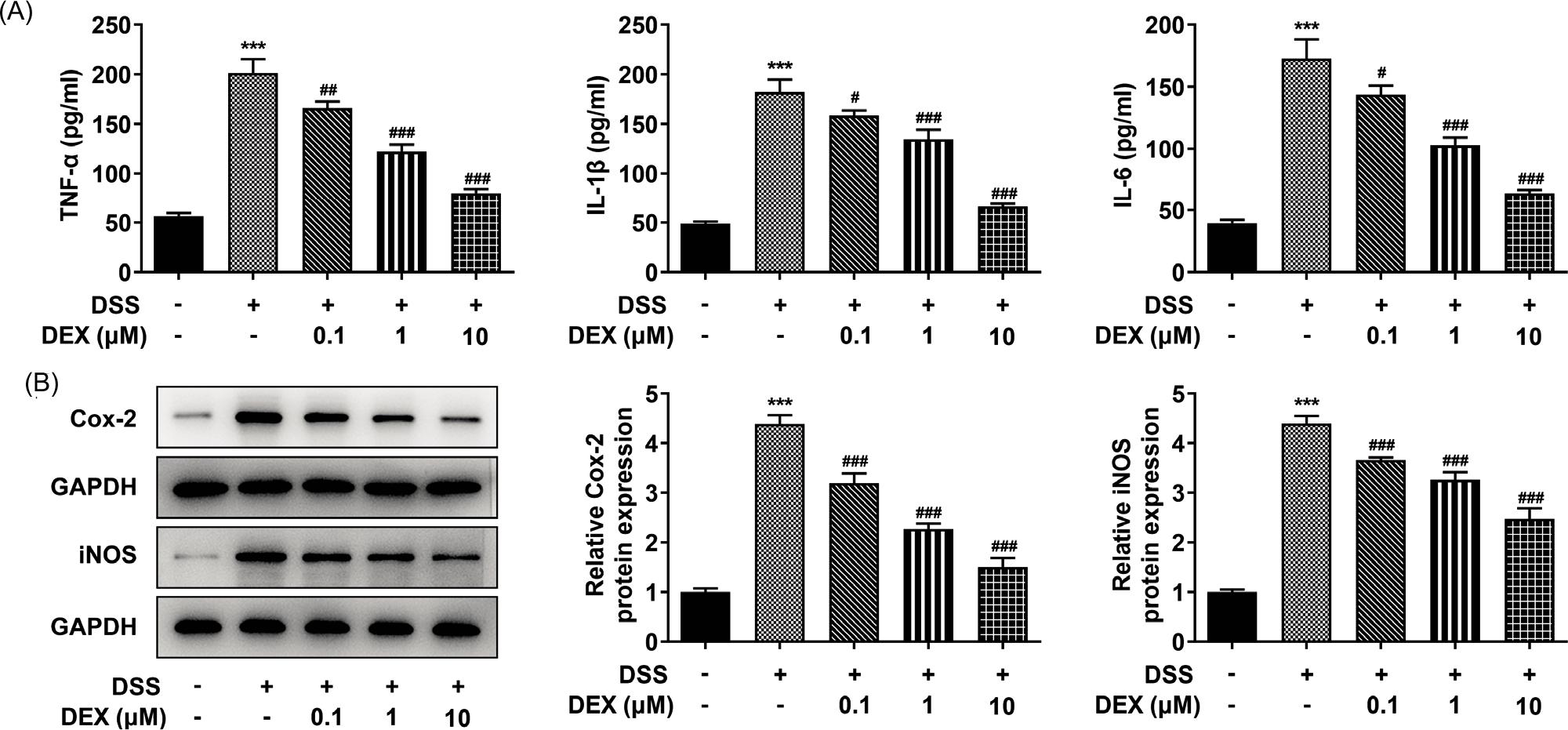
The levels of TNF-α, IL-1β and IL-6, which were deemed as inflammatory factors, and the expressions of inflammation-related factors (COX-2 and iNOS) were significantly increased in DSS-induced NCM460 cells (Figures 2A and B). However, DEX hampered inflammation dependent on increasing concentrations.
Figure 2 DEX suppresses inflammatory response in DSS-induced NCM460 cells. (A) Levels of inflammatory factors in DSS-induced NCM460 cells treated with DEX were determined by their commercial kits. (B) Western blot analysis determined the influence of DEX on the expression of inflammation-related factors in DSS-treated NCM460 cells. ***P < 0.001 vs. control group; #P < 0.05, ##P < 0.01 and ###P < 0.001 vs. DSS group.
DEX inhibits barrier damage in DSS-induced NCM460 cells
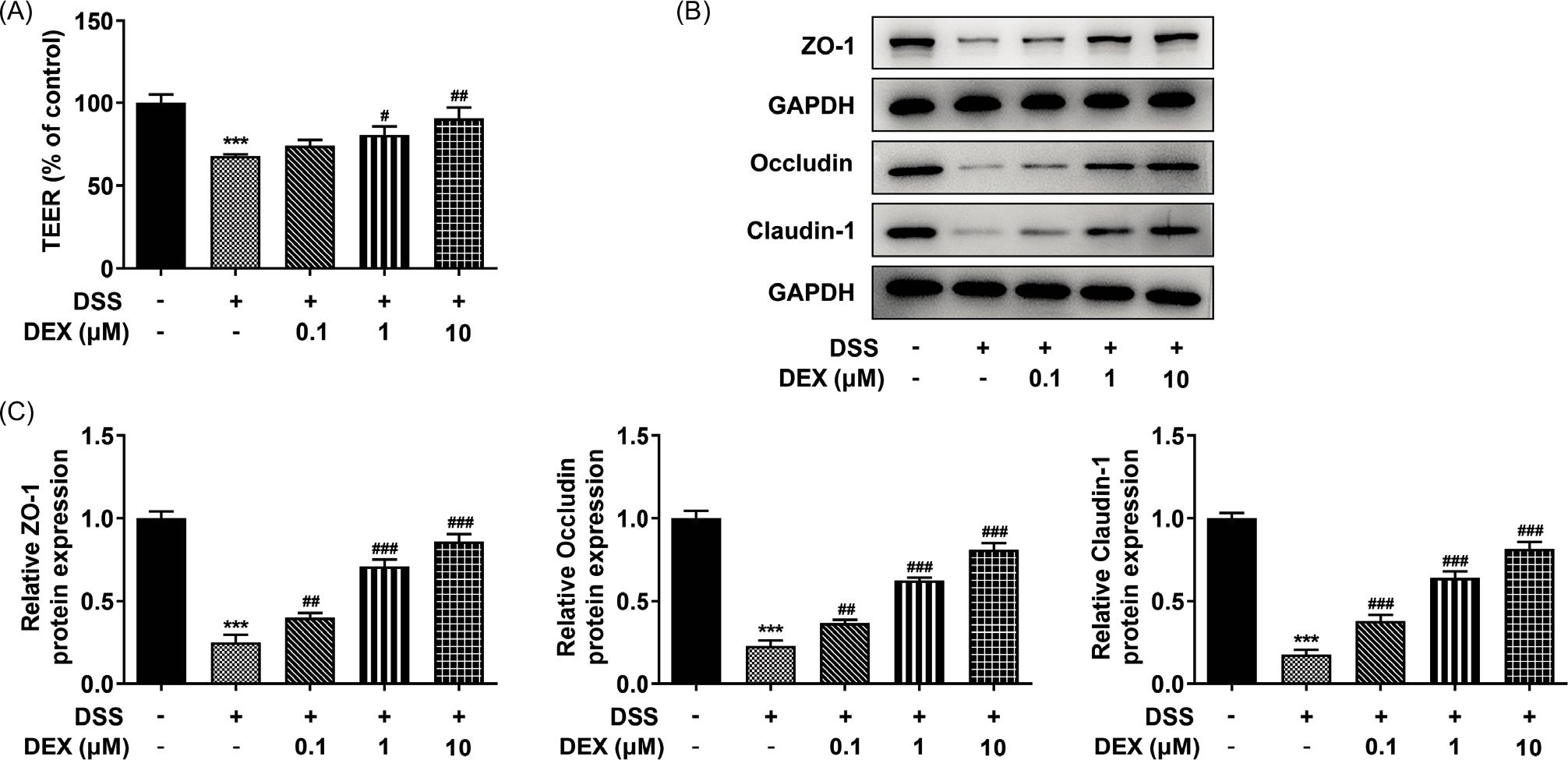
As exhibited in Figure 3A, DSS reduced the level of TEER while DEX improved the reduced TEER. Besides, ZO-1, occludin and claudin-1 protein levels were downregulated in NCM460 cells exposed to DSS. This DSS-induced alteration of tight junction protein expression was reversed by DEX (Figures 3B and C).
Figure 3 DEX inhibits barrier damage in DSS-induced NCM460 cells. (A) The permeability of DSS-induced NCM460 monolayers treated with DEX was detected by TEER measurement. (B) and (C) Expression of tight junction proteins in DSS-induced NCM460 cells treated with DEX was detected by Western blot analysis. ***P < 0.001 vs. control group. #P < 0.05, ##P < 0.01 and ###P < 0.001 vs. DSS group.
DEX inhibits the RhoA/ROCK signaling pathway in DSS-induced NCM460 cells
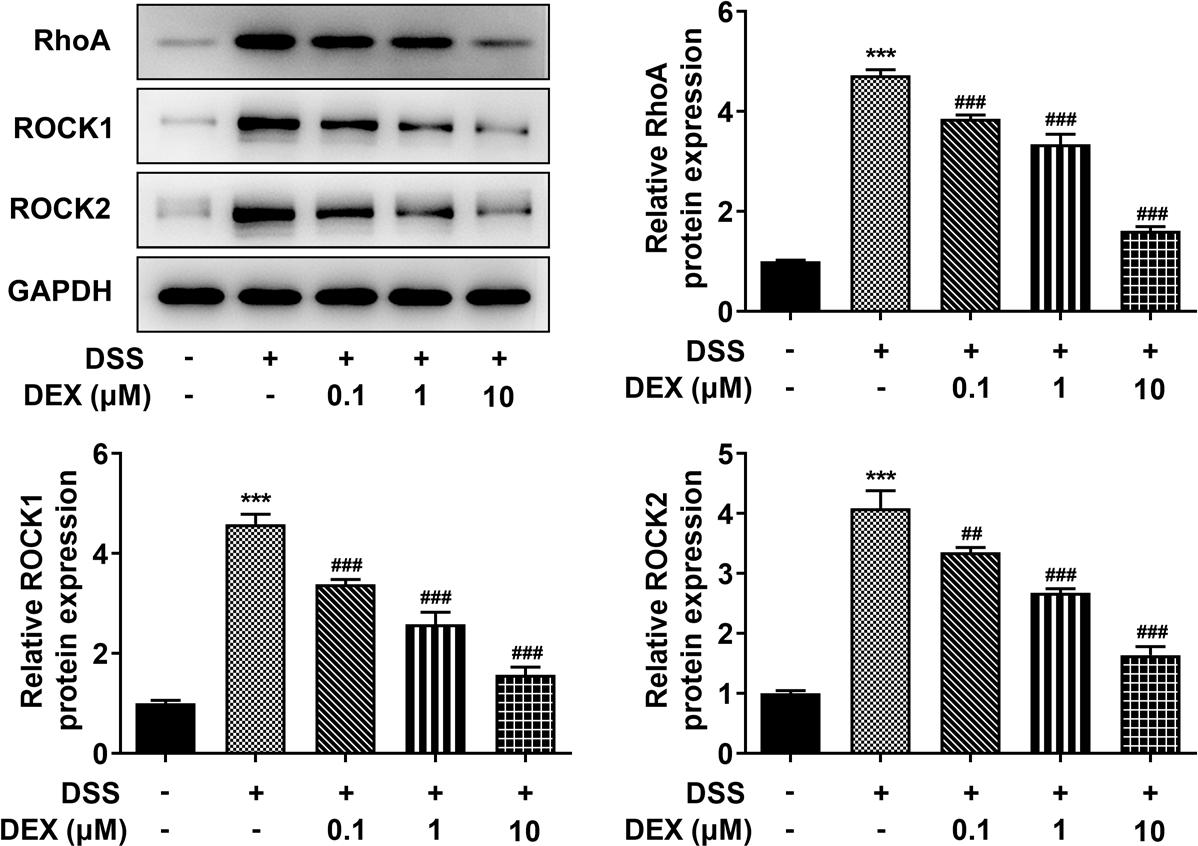
In order to probe the association of DEX with RhoA/ROCK pathway activation stimulated by DSS, Western blot analysis was used to test the protein levels of RhoA, ROCK1 and ROCK2. It turned out that expressions of RhoA, ROCK1 and ROCK2 were elevated in the DSS group. However, these effects were significantly inhibited by DEX (Figure 4).
Figure 4 DEX inhibits the RhoA/Rock signaling pathway in DSS-induced NCM460 cells. Expression of RhoA/ROCK signaling pathway-related proteins in DSS-induced NCM460 cells treated with DEX was determined by Western blot analysis. ***P < 0.001 vs. control group. ##P < 0.01 and ###P < 0.001 vs. DSS group.
U46619 weakens the inhibiting effect of DEX on inflammation in DSS-induced NCM460 cells
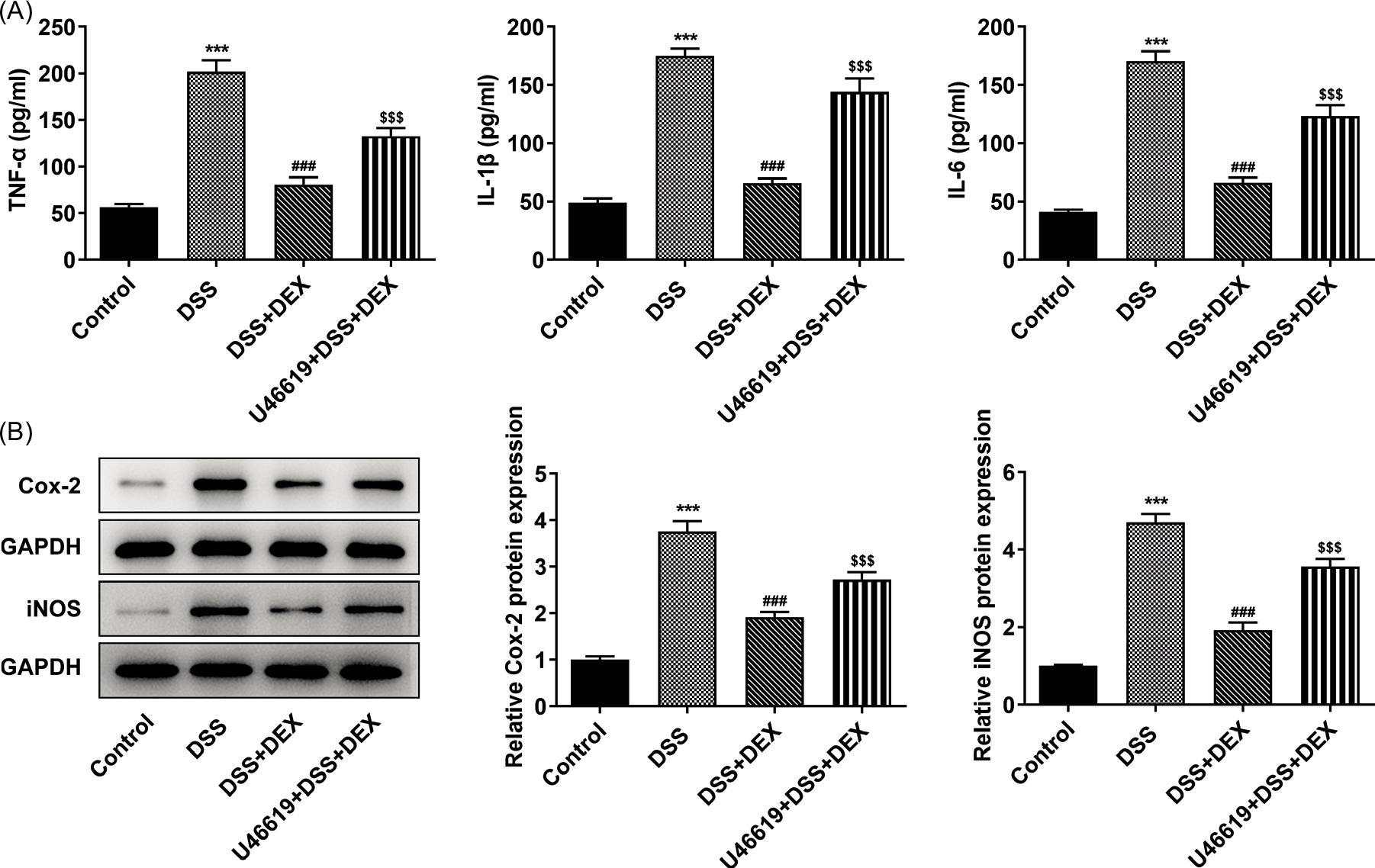
Regarding the protective effects of different concentrations of DEX observed in previous studies, 10-μM DEX was selected for subsequent experiment. U46619 weakened the protective effects of DEX on inflammation, as reflected by the upregulated levels of inflammatory factors (TNF-α, IL-1β and IL-6; Figure 5A) and increased expressions of inflammation-related proteins (COX-2 and iNOS; Figure 5B).
Figure 5 U46619 weakens the inhibiting effect of DEX on inflammation in DSS-induced NCM460 cells. (A) Levels of inflammatory factors in DSS-induced NCM460 cells treated with DEX and U46619 were determined by their commercial kits. (B) Western blot analysis tested the impact of DEX and U46619 on the expression of inflammation-related factors in DSS-treated NCM460 cells. Expression of inflammation-related proteins in DSS-induced NCM460 cells treated with DEX and U46619 was detected by Western blot analysis. ***P < 0.001 vs. control group; ###P < 0.001 vs. DSS group; $$$P < 0.001 vs. DSS+DEX group.
U46619 weakens the inhibiting effect of DEX on barrier damage in DSS-induced NCM460 cells
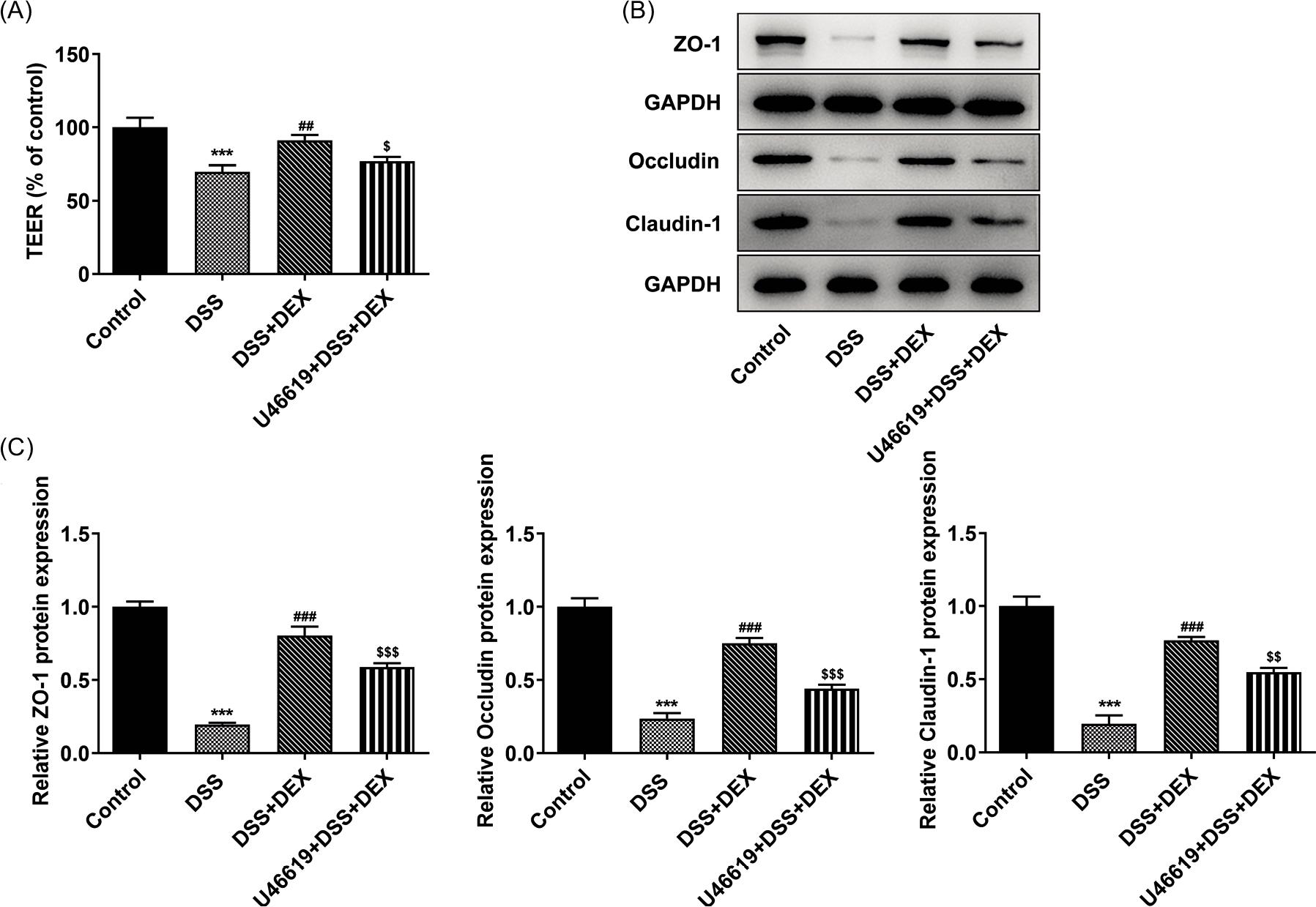
U46619 decreased the level of TEER in DSS-induced NCM460 cells treated with DEX (Figure 6A). Correspondingly, expressions of ZO-1, occluding and claudin-1 were down-regulated after U46619 treatment in DSS-induced NCM460 cells (Figure 6B).
Figure 6 U46619 weakens the inhibiting effect of DEX on barrier damage in DSS-induced NCM460 cells. (A) The permeability of DSS-induced NCM460 monolayers treated with DEX and U46619 was detected by TEER measurement. (B) and (C) Expression of tight junction proteins in DSS-induced NCM460 cells treated with DEX and U46619 was detected by Western blot analysis. ***P < 0.001 vs. control group. ##P < 0.01 and ###P < 0.001 vs. DSS group. $P < 0.05, $$P < 0.01 and $$$P < 0.001 vs. DSS+DEX group.
Discussion
Presently, treatments for ulcerative colitis continue to develop, but available therapies are limited to symptomatic relief and non-specific immunomodulators.18 Therefore, the treatment of ulcerative colitis remains an area of significant medical requirement.
It is determined that high levels of pro-inflammatory mediators such as cytokines (TNF-α IL-1β and IL-6) is a dominant feature of ulcerative colitis which leads to chronic inflammatory ulcers and in turn leukocyte infiltration of colonic mucosal injury.19 TNF-α is deemed as a driving factor for the destruction of epithelial barrier, inducing apoptosis of epithelial cells. IL-6 is related to the recruitment and activation of neutrophils, exacerbating the inflammatory state of inflammatory enteritis. Excessive production of IL-1β and IL-6 may lead to increased intestinal inflammation and intestinal motility disorders.18
COX-2 is an important target gene product of hypoxia regulatory factors and is engaged in the regulation of apoptosis, angiogenesis, inflammation and tumorigenesis.20,21 In vivo prostaglandin E2 (PGE2), nitric oxide (NO) and IL-17 demonstrated strong pro-inflammatory response. iNOS is a protein-independent enzyme primarily expressed in inflammatory cells and is an important factor in inducing NO production.22,23 COX-2 and iNOS are key upstream enzymes for the synthesis of PGE2 and NO, and their abundance expression can directly limit the production of downstream PGE2 and NO.24 In this study, TNF-α, IL-1β and IL-6 levels and COX-2 and iNOS expressions were enhanced in DSS-exposed NCM460 cells which were lessened after DEX treatment.
Intestinal mucosal barrier function plays an important role in the development of ulcerative colitis.25,26 The intestinal mucosal barrier is the first barrier against harsh environment and is primarily formed by tight junctions (TJs) of epithelial cells. Tight junctions consist of transmembrane proteins (occludins and claudins) and helper protein (ZO) to prevent the spread of pathogens and harmful antigens within the epithelium.27 Zonula occludens, occludins and claudins can seal gap between adjacent intestinal epithelial cells and retain antigens and microorganisms in the intestinal lumen which occupies a significant position for maintaining intestinal permeability, tissue differentiation and internal environment stability.28 Therefore, it is important to regulate tight junctions to maintain integrity of the ulcerative colitis epithelial barrier. Here, ZO-1, occludin and claudin-1 expressions were declined apparently, accompanied by the decreased TEER level. Nonetheless, DEX could reverse the above results.
ROCK, chief effector in the downstream of RhoA, has been demonstrated to regulate biological pathways, affecting smooth muscle tension levels and various physiological characteristics related to actin cytoskeletal changes.29,30 Recent reports have documented that the RhoA/ROCK signaling pathway is implicated in the activation of immune system and brings about the release of pro-inflammatory factors.11 Brusatol-enriched Brucea javanica oil (BE-BJO) inhibited NF-κB and RhoA/ROCK signaling pathway by suppressing colonic inflammation and enhancing intestinal epithelial barrier function to ameliorate DSS ulcerative colitis.31 Oxymatrine could relieve DSS-induced acute intestinal inflammation by blocking RhoA/ROCK signaling pathway to protect the integrity of epithelial barrier, alleviate inflammation and oxidative stress, and maintain balance between Th17 and Treg cells.32 Our experimental results established that the RhoA/ROCK signaling pathway was activated in NCM460 cells after DSS induction, which was suppressed by DEX. In addition, U46619, an agonist of RhoA/ROCK signaling pathway, could weaken the effect of DEX on DSS-induced NCM460 cells to promote inflammation and reduce TEER level.
Conclusion
Our results indicated that DEX could protect against inflammation and barrier damage in DSS-induced NCM460 cells by inhibiting the RhoA/ROCK signaling pathway. These findings confirmed a therapeutic potential of DEX in ulcerative colitis. Are there other signaling pathways involved in DEX effect on DSS-treated NCM460 cells is required to be explored in the future research.
Competing interests
The authors declared that they have no competing interests.
Availability of data and material
The experimental data will be available on request.
REFERENCES
1. Lebeer S, Vanderleyden J, De Keersmaecker SC. Host interactions of probiotic bacterial surface molecules: Comparison with commensals and pathogens. Nature Rev Microbiol. 2010;8:171–84. 10.1038/nrmicro2297
2. Adams SM and Bornemann PH. Ulcerative colitis. Am Family Physician. 2013;87:699–705.
3. Eisenstein M. Ulcerative colitis: Towards remission. Nature. 2018;563:S33. 10.1038/d41586-018-07276-2
4. Hou JK, El-Serag H, Thirumurthi S. Distribution and manifestations of inflammatory bowel disease in Asians, Hispanics, and African Americans: A systematic review. AmJ Gastroenterol. 2009;104:2100–9. 10.1038/ajg.2009.190
5. Weerink MAS, Struys M, Hannivoort LN, Barends CRM, Absalom AR, Colin P. Clinical pharmacokinetics and pharmacodynamics of dexmedetomidine. Clin Pharmacokinet. 2017;56:893–913. 10.1007/s40262-017-0507-7
6. Yang J, Wu Y, Xu Y, Jia J, Xi W, Deng H, et al. Dexmedetomidine resists intestinal ischemia-reperfusion injury by inhibiting TLR4/MyD88/NF-κB signaling. J Surg Res. 2021;260:350–8. 10.1016/j.jss.2020.11.041
7. Qin C, Jiang Y, Chen X, Bian Y, Wang Y, Xie K, et al. Dexmedetomidine protects against burn-induced intestinal barrier injury via the MLCK/p-MLC signalling pathway. Burns J Int Soc Burn Inj. 2021;47:1576–85. 10.1016/j.burns.2021.01.017
8. Chen Y, Miao L, Yao Y, Wu W, Wu X, Gong C, et al. Dexmedetomidine ameliorates CLP-induced rat intestinal injury via inhibition of inflammation. Mediat Inflamm. 2015;2015:918361. 10.1155/2015/918361
9. Chen CJ, Xiao P, Chen Y, Fang R. Selenium deficiency affects uterine smooth muscle contraction through regulation of the RhoA/ROCK signalling pathway in mice. Biol Trace Elem Res. 2019;192:277–86. 10.1007/s12011-019-01677-8
10. Knipe RS, Probst CK, Lagares D, Franklin A, Spinney JJ, Brazee PL, et al. The Rho kinase isoforms ROCK1 and ROCK2 each contribute to the development of experimental pulmonary fibrosis. Am J Resp Cell Mol Biol. 2018;58:471–81. 10.1165/rcmb.2017-0075OC
11. Yang W, Zhou G, Yu T, Chen L, Yu L, Guo Y, et al. Critical role of ROCK2 activity in facilitating mucosal CD4(+) T cell activation in inflammatory bowel disease. J Autoimmun. 2018;89:125–38. 10.1016/j.jaut.2017.12.009
12. Zou Y, Ma L, Zhao Y, Zhang S, Zhou C, Cai Y. Inhibition of Rho kinase protects against colitis in mice by attenuating intestinal epithelial barrier dysfunction via MLC and the NF-κB pathway. Int J Mol Med. 2018;41:430–8. 10.3892/ijmm.2017.3197
13. Kang W, Shang L, Wang T, Liu H, Ge S. Rho-kinase inhibitor Y-27632 downregulates LPS-induced IL-6 and IL-8 production via blocking p38 MAPK and NF-κB pathways in human gingival fibroblasts. J Periodontol. 2018;89:883–93. 10.1002/JPER.17-0571
14. Wang S, Lin Y, Yuan X, Li F, Guo L, Wu B. REV-ERBα integrates colon clock with experimental colitis through regulation of NF-κB/NLRP3 axis. Nature Communicat. 2018;9:4246. 10.1038/s41467-018-06568-5
15. Hu T, Lin Q, Guo T, Yang T, Zhou W, Deng X, et al. Polysaccharide isolated from Phellinus linteus mycelia exerts anti-inflammatory effects via MAPK and PPAR signaling pathways. Carbohyd Polym. 2018;200:487–97. 10.1016/j.carbpol.2018.08.021
16. Sun T, Gong Q, Wu Y, Shen Z, Zhang Y, Ge S, et al. Dexmedetomidine alleviates cardiomyocyte apoptosis and cardiac dysfunction may be associated with inhibition of RhoA/ROCK pathway in mice with myocardial infarction. Naunyn-Schmiedeberg’s Arch Pharmacol. 2021;394:1569–77. 10.1007/s00210-021-02082-6
17. Wang Y, Chen H, Chen Q, Jiao FZ, Zhang WB, Gong ZJ. The protective mechanism of CAY10683 on intestinal mucosal barrier in acute liver failure through LPS/TLR4/MyD88 pathway. Mediat Inflamm. 2018;2018:7859601. 10.1155/2018/7859601
18. Zizzo MG, Caldara G, Bellanca A, Nuzzo D, Di Carlo M, Serio R. Preventive effects of guanosine on intestinal inflammation in 2,4-dinitrobenzene sulfonic acid (DNBS)-induced colitis in rats. Inflammopharmacol. 2019;27:349–59. 10.1007/s10787-018-0506-9
19. Bevivino G, Monteleone G. Advances in understanding the role of cytokines in inflammatory bowel disease. Expert Rev Gastroenterol Hepatol. 2018;12:907–15. 10.1080/17474124.2018.1503053
20. El Miedany Y, Youssef S, Ahmed I, El Gaafary M. The gastrointestinal safety and effect on disease activity of etoricoxib, a selective cox-2 inhibitor in inflammatory bowel diseases. Am J Gastroenterol. 2006;101:311–7. 10.1111/j.1572-0241.2006.00384.x
21. El-Gowelli HM, Saad EI, Abdel-Galil AG, Ibrahim ER. Co-administration of α-lipoic acid and cyclosporine aggravates colon ulceration of acetic acid-induced ulcerative colitis via facilitation of NO/COX-2/miR-210 cascade. Toxicol Appl Pharmacol. 2015;288:300–12. 10.1016/j.taap.2015.08.002
22. Yu L, Yan J, Sun Z. D-limonene exhibits anti-inflammatory and antioxidant properties in an ulcerative colitis rat model via regulation of iNOS, COX-2, PGE2 and ERK signaling pathways. Mol Med Rep. 2017;15:2339–46. 10.3892/mmr.2017.6241
23. Guo Y, Wu X, Wu Q, Lu Y, Shi J, Chen X. Dihydrotanshinone I, a natural product, ameliorates DSS-induced experimental ulcerative colitis in mice. Toxicol Appl Pharmacol. 2018;344:35–45. 10.1016/j.taap.2018.02.018
24. Heng-Guang Z. Effects of tetrandrine on COX-2/PGE_2,iNOS/NO expression in LPS-stimulated RAW264. 7 cells.J China Pharm Univ. 2011;42:141–4.
25. Li C, Cai YY, Yan ZX. Brain-derived neurotrophic factor preserves intestinal mucosal barrier function and alters gut microbiota in mice. Kaohsiung J Med Sci. 2018;34:134–41. 10.1016/j.kjms.2017.11.002
26. Chen S, Zuo S, Zhu J, Yue T, Bu D, Wang X, et al. Decreased expression of cystathionine β-synthase exacerbates intestinal barrier injury in ulcerative colitis. J Crohn’s Colitis. 2019;13:1067–80. 10.1093/ecco-jcc/jjz027
27. Gai L, Chu L, Xia R, Chen Q, Sun X. Barbaloin attenuates mucosal damage in experimental models of rat colitis by regulating inflammation and the AMPK signaling pathway. Med Sci Monitor Int Med J Exp Clin Res. 2019;25:10045–56. 10.12659/MSM.918935
28. Wang J, Zhang C, Guo C, Li X. Chitosan ameliorates DSS-induced ulcerative colitis mice by enhancing intestinal barrier function and improving microflora. Int J Mol Sci. 2019;20:5751. 10.3390/ijms20225751
29. Al-Shboul O. The role of the RhoA/ROCK pathway in gender-dependent differences in gastric smooth muscle contraction. J Physiol Sci (JPS). 2016;66:85–92. 10.1007/s12576-015-0400-9
30. Del Re DP, Miyamoto S, Brown JH. RhoA/Rho kinase up-regulate Bax to activate a mitochondrial death pathway and induce cardiomyocyte apoptosis. J Biol Chem. 2007;282:8069–78. 10.1074/jbc.M604298200
31. Zheng X, Mai L, Wang T, Xu Y, Su Z, Chen J, et al. Brusatol-enriched brucea javanica oil ameliorated dextran sulfate sodium-induced colitis in mice: Involvement of NF-κB and RhoA/ROCK signaling pathways. BioMed Res Int. 2021;2021:5561221. 10.1155/2021/5561221
32. Wang Y, Shou Z, Fan H, Xu M, Chen Q, Tang Q, et al. Protective effects of oxymatrine against DSS-induced acute intestinal inflammation in mice via blocking the RhoA/ROCK signaling pathway. Biosci Rep. 2019;39:BSR20182297. 10.1042/BSR20182297