
Download
ORIGINAL ARTICLE
Bruceine D ameliorates the balance of Th1/Th2 in a mouse model of ovalbumin-induced allergic asthma via inhibiting the NOTCH pathway
Ying Niea#, Bangkun Yangb#, Junfeng Hua, Lingling Zhanga, Zhimin Mac*
aDepartment of Pediatrics, Wuhan No.1 Hospital, Wuhan City, China
bDepartment of Neurosurgery, Zhongnan Hospital of Wuhan University, Wuhan City, China
cDepartment of Respiratory Medicine, The Second Hospital of Hebei Medical University, Shijiazhuang City, China
Abstract
Allergic asthma is a heterogeneous inflammatory disorder triggered by inhaled allergens, leading to airflow obstruction, bronchial inflammation, and airway hyperresponsiveness (AHR). T helper (Th) 2 cell-mediated immune response and airway inflammation are the key features of allergic asthma. Bruceine D (BD) is a bioactive compound extracted from the seeds of Brucea javanica. The present study aimed to investigate the effects of increased doses of BD on AHR, secretion of Th1-/Th2-associated cytokines, and inflammatory cell infiltration in ovalbumin (OVA)-induced allergic asthma mice. The results showed that BD reduced OVA-induced inflammatory cell infiltration and bronchial hyperresponsiveness into the peribronchial tissues and perivascular areas. Mice treated with BD also showed significantly decreased expressions of Th2-associated cytokines (i.e., interleukin (IL)-4, IL-5, and IL-13) and elevated production of Th1-associated cytokines (i.e., interferon gamma and IL-2) following OVA stimulation. BD treatment dose-dependently inhibited OVA-induced accumulation of inflammatory cells in asthmatic mice. Further analysis revealed that OVA exposure upregulated pulmonary expressions of NOTCH signaling receptors, a group of transmembrane proteins that communicate signals upon binding to transmembrane ligands expressed on adjacent cells, while BD treatment significantly abolished OVA-induced activation of the NOTCH pathway. In conclusion, BD protected mice against OVA-induced allergic asthma by reducing AHR and restoring the Th1/Th2 balance through the NOTCH signaling pathway. Our findings highlighted the potential of BD as a therapeutic agent for allergic asthma.
Key words: Bruceine D, NOTCH, ovalbumin-induced allergic asthma, T helper 1 cells, T helper 2 cells
*Corresponding author: Zhimin Ma, Department of Respiratory Medicine, The Second Hospital of Hebei Medical University, No. 215 Heping West Road, Shijiazhuang City, Hebei Province, China. Tel: +86-0311-66007372. Email address: [email protected]
#These authors contributed equally to the work.
Received 6 September 2021; Accepted 19 October 2021; Available online 4 November 2021
Copyright: Nie Y, et al.
License: This open access article is licensed under Creative Commons Attribution 4.0 International (CC BY 4.0). http://creativecommons.org/licenses/by/4.0/
Introduction
Allergic asthma is a chronic, heterogeneous inflammatory syndrome triggered by inhaled allergens, leading to airflow obstruction, bronchial inflammation, airway hyperresponsiveness (AHR), and possibly allergic shock.1 The prevalence of allergic asthma has been increasing from 5.0 to 7.3% over the past two decades, while the prevalence of nonatopic asthma remained stable around 3.5%.2 The highest incidence of allergic asthma is observed in early childhood.3 Bronchodilators, such as β2-adrenoceptor agonists and inhaled corticosteroids, as well as anti-inflammatory drugs are the most widely used treatment strategies for allergic asthma, but varied treatment responses have been observed in different individuals.4 Hence, the development of new therapeutic strategies with high effectiveness and improved reliability is of great importance for patients with allergic asthma.
T helper cells play crucial roles in the pathophysiological process of allergic asthma.5,6 T helper 2 cell (Th2)-mediated immune response and airway inflammation are the key characteristics of this chronic disorder.7 Upregulation of Th2-associated cytokines, such as interleukin (IL)-4, IL-5, and IL-13, in asthmatic patients induces the infiltration and activation of eosinophils in the lung.8 In contrast, the cytokines released by T helper 1 cells (Th1), such as interferon gamma (IFN-γ), have been shown to inhibit the development of allergic asthma.9 Thus, therapeutic agents that can restore the Th1/Th2 balance may be of interest for the treatment of allergic asthma.
Bruceine D (BD) is a natural, bioactive compound extracted from the seeds of Brucea javanica.10 Previous literature has reported that BD acts as a tumor suppressor in different types of human cancers, including lung carcinoma,11 gastric cancer,12 and breast cancer.13 It also plays a neuroprotective role in Parkinson’s disease by inhibiting oxidative stress and the production of inflammatory factors in the brain.14 BD is proved to be an inhibitor of NOTCH signaling receptors, a group of transmembrane proteins that communicate signals upon binding to transmembrane ligands expressed on adjacent cells.15 A recent study reported that inhibition of the NOTCH pathway reduced airway inflammation in asthmatic mice.16 However, whether Bruceine D can alleviate allergic asthma and whether the NOTCH pathway is involved in this process remain unknown.
In the present study, we used ovalbumin (OVA) to induce allergic asthma in mice and investigated the effects of Bruceine D on AHR, secretion of Th1/Th2-associated cytokines, and infiltration of inflammatory cell in mice following OVA challenge. The effect of Bruceine D on the NOTCH pathway in asthmatic mice was also explored.
Materials and methods
Animal model of allergic asthma
Twenty-five male BALB/c mice (8 weeks old, 18–20 g) were purchased from Shanghai Laboratory Animal Center (Shanghai, China) and housed in a specific pathogen-free environment with a temperature of 22–24°C, 12-h alternating light–dark cycle, and 50–60% humidity. All animals had ad libitum access to water and food. All experiments were approved by the Animal Ethics Committee of Wuhan Myhalic Biotechnology Co., Ltd. (Approval no. HLK-20210202-01) and performed following the Guide for the Care and Use of Laboratory Animals.17
After 1 week of acclimation, mice were randomly assigned to five groups (n = 5 per group): Sham, OVA, OVA + 10 mg/kg BD, OVA + 20 mg/kg BD, and OVA + 40 mg/kg BD. A model of OVA-induced allergic asthma was established as previously reported.18 Briefly, all mice, except those in the Sham group, were intraperitoneally injected with 200 μg of OVA (Sigma, St. Louis, USA) and 1 mg of aluminum hydroxide (Sigma) dissolved in 0.2 mL of normal saline on Day 1, 8, and 15. From Day 22 to 25, mice were placed in an incompletely closed container and continuously exposed to 2.5% OVA (30 min per day) using an ultrasonic nebulizer (Omron Co., Tokyo, Japan).
Drug treatment
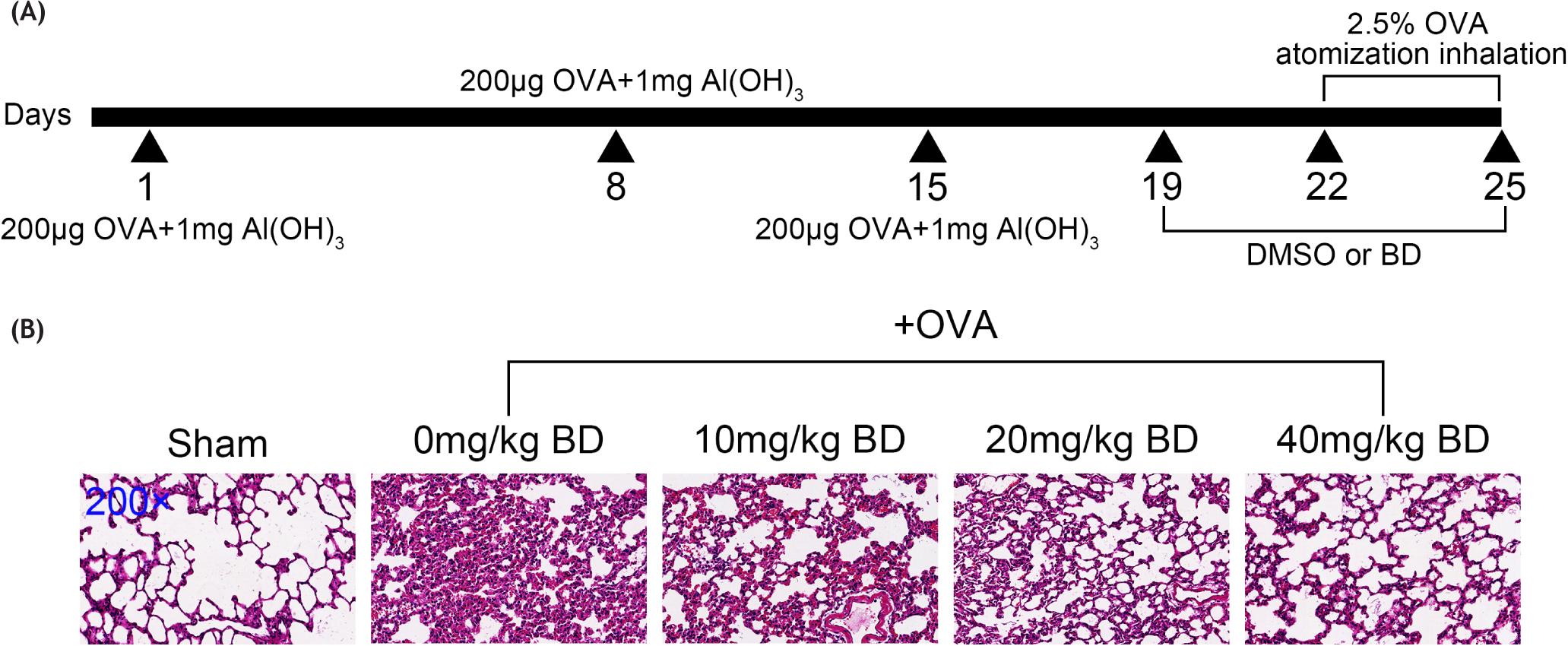
BD (≥97% purity; CAS Number: 21499-66-1) was purchased from Tauto Biotech Co., Ltd. (Shanghai, China) and dissolved in DMSO (Sigma) to different concentrations. From Day 19 to Day 25 of OVA stimulation, model mice subjected to BD treatment were intraperitoneally injected once per day with different concentrations of BD (10, 20, and 40 mg/kg) 1 h before OVA challenge, while the OVA group were treated with an equal amount of DMSO. The Sham group remained untreated. Schematic diagram of the experimental procedure is shown in Figure 1A.
Figure 1 Pulmonary morphological changes in OVA-challenged mice treated with or without BD. (A) Schematic diagram of the experimental procedure. (B) An animal model of OVA-induced allergic asthma was established in BALB/c mice. The OVA, OVA + 10 mg/kg BD, OVA + 20 mg/kg BD, and OVA + 40 mg/kg BD groups (n = 5 per group) were treated with BD at various doses or DMSO from Day 19 to Day 25 of OVA stimulation. The Sham group (n = 5) received no OVA exposure. The lung tissues were harvested at 24 h after the last OVA inhalation, sectioned, and stained with H&E. The images were observed at ×200 magnification.
Histopathological analysis of lung tissues
Immediately after the AHR assessment, all animals were euthanized by an intraperitoneal injection of Pentobarbital Sodium (1%, 80 mg/kg). The lung tissues were harvested and stored at -80°C. A proportion of tissue samples was fixed in 4% paraformaldehyde overnight and then cut into 5-μm sections. The sections were then dewaxed, dehydrated in ethanol, and stained with hematoxylin and eosin (H&E; Sigma). Microscopic images were captured at ×200 magnification using a light microscope (Olympus, Tokyo Japan).
Assessment of airway hyperresponsiveness
At 24 h after the last OVA inhalation, mice were tested for AHR using the Buxco Noninvasive Airway Mechanics System (Buxco Electronics Inc., Wilmington, USA) as previously mentioned.16 Animals were anesthetized by an intraperitoneal injection of 7% hydrate chloral (0.5 mL/100 g) and then exposed to aerosolized normal saline or increasing concentrations (10–50 mg/mL) of methacholine (Sigma). The airway resistance in the cannula was assessed.
Enzyme-linked immunosorbent assay
The lung tissues were homogenized using RIPA buffer with protease inhibitors (Bio-Rad). The pulmonary concentrations of IFN-γ, IL-2, IL-4, IL-5, and IL-13 were measured using enzyme-linked immunosorbent assay (ELISA) kits (BioSource International, Camarillo, USA). The optical density was measured at 450 nm using a microplate reader (Bio-TEK, Winooski, USA).
Inflammatory cell counting
Blood samples were obtained from the brachial plexus of each mouse after the AHR assessment, and centrifuged at 1300 × g for 10 min at 4°C. The serum supernatant was then collected. Inflammatory cells, including white blood cells, monocytes, basophils, neutrophils, lymphocytes, and eosinophils were counted using a Hemavet 950 Hematology System (Drew Scientific, Inc., Miami Lakes, USA)
Western blot
The homogenized lung tissue samples were prepared using RIPA buffer with protease inhibitors (Bio-Rad). The bicinchoninic acid assay (Pierce, Rockford, USA) was used to determine protein concentration. An equal amount of protein samples (50 µg) were separated by a 10% SDS-PAGE gel and transferred onto the PVDF membrane (MilliporeSigma, Burlington, USA). After blocking with 5% nonfat dry milk, the PVDF membranes were incubated with designated primary antibodies overnight at 4°C: NOTCH1 (1:1000; cat no. 52627, Abcam, Cambridge, USA), NOTCH2 (1:1000; cat no. 518049, Santa Cruz Biotechnology, Dallas, USA), NOTCH3 (1:1000; cat no. 2889, Cell Signaling Technology, Danvers, USA), DLL4 (1:1000; cat no. 270534, Abcam), and β-actin (1:3000; cat no. 8226, Abcam). After washed with Tris-buffered saline tween, the membranes were incubated with a secondary antibody (1:5000; cat no. 6721, Abcam) at room temperature for 1 h. The bands were visualized by Alphalmager 2000 Imaging System (Alpha Innotech, San Leandro, USA) and quantified using the Image J software.
Statistical analysis
Data were analyzed by the SPSS software (v. 24.0) and presented as mean ± standard deviation from three independent experiments. One-way ANOVA followed by Bonferroni’s post-hoc test was performed to determine statistical significance among groups. A p-value of <0.05 was considered statistically significant.
Results
Pulmonary morphological alterations in mice with allergic asthma
To investigate the effects of BD on asthma and the potential pathways involved in this process, we established an OVA-induced allergic asthma mouse model and treated mice with or without BD at various concentrations. Compared with the Sham group, OVA-challenged mice showed severe inflammatory cell infiltration into the peribronchial connective tissues and perivascular areas. The BD treatment, however, reduced infiltration of inflammatory cells, and ameliorated fibrosis around the airways and vessels in OVA-stimulated mice. The higher the dose of BD given to mice, the less the morphological alterations observed in the lungs (Figure 1B). The histopathological analysis indicated that BD treatment repaired OVA-induced pulmonary tissue injury in mice.
Effect of BD on OVA-induced AHR in mice
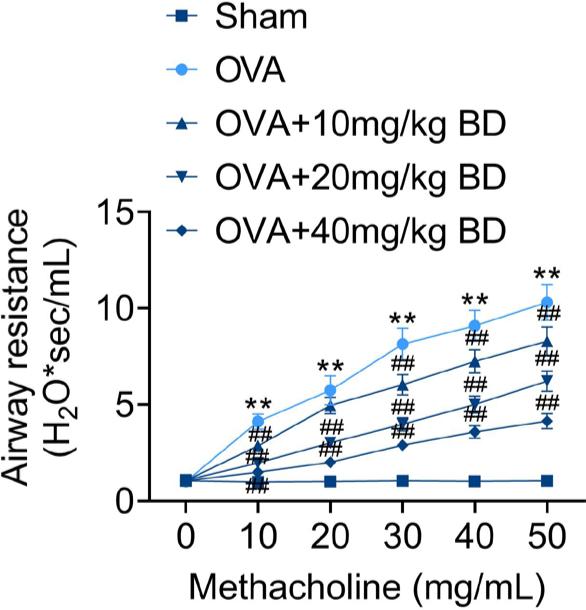
Since increased bronchial hyperresponsiveness is the main manifestation of asthma, we further investigated the effect of BD on airway resistance. Compared with the Sham group, OVA-challenged mice showed significantly increased airway resistance when the concentration of methacholine exceeded 10 mg/mL. BD significantly decreased OVA-induced airway resistance in mice at all concentrations, and the most robust effect was observed at the highest concentration (40 mg/kg) (Figure 2). These results suggested that BD alleviated OVA-induced AHR in asthmatic mice.
Figure 2 Effect of BD on AHR in mice with OVA-induced allergic asthma. The AHR assessment was performed 24 h after the final OVA challenge. Mice were administered with increasing concentrations of methacholine (0–50 mg/mL) under anesthesia. The airway resistance of each mouse was assessed. **p < 0.01 compared with the Sham group; ##p < 0.01 compared with the OVA group.
Effect of BD on Th1-/Th2-associated cytokines in mice with OVA-induced asthma
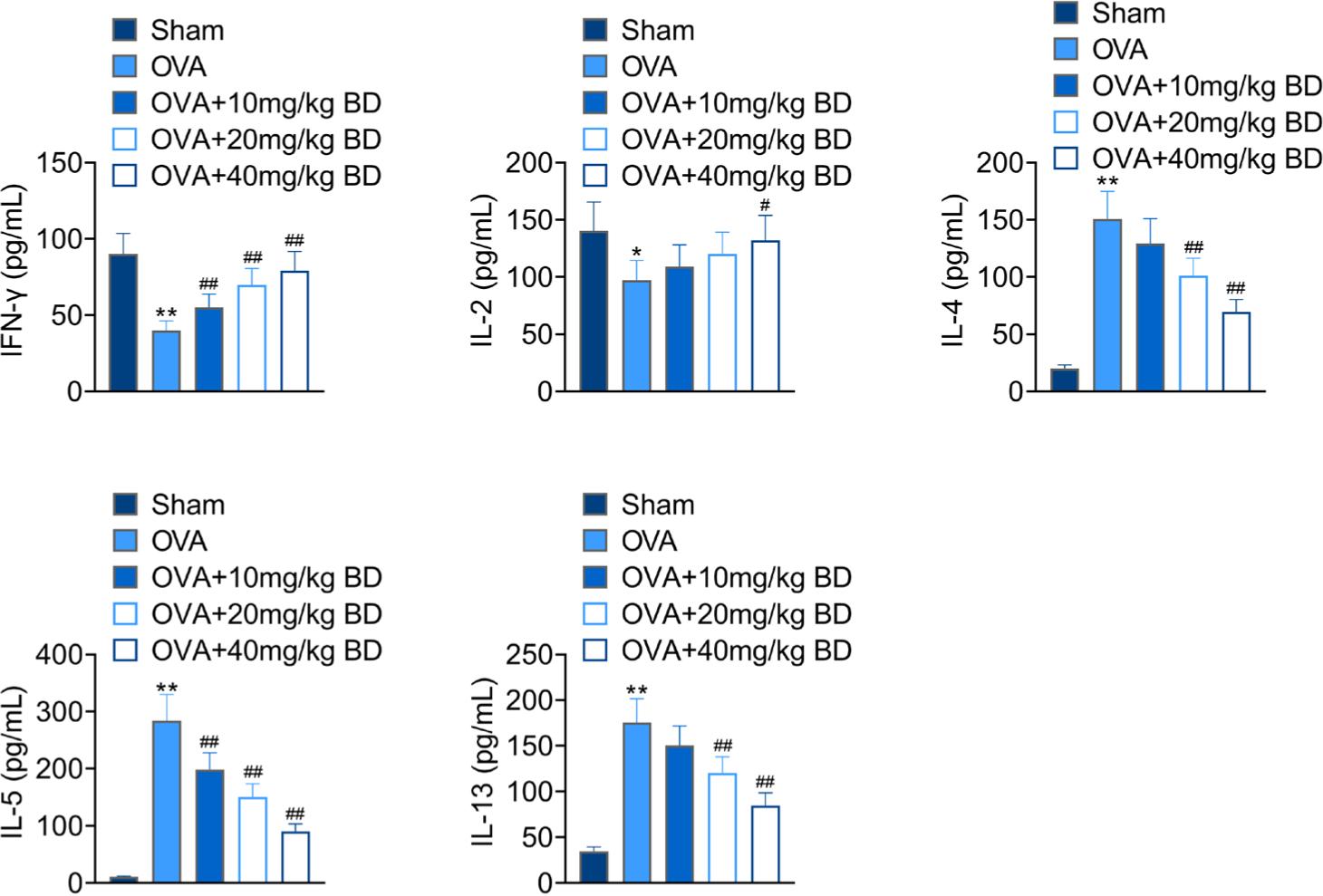
To explore whether BD affected the Th1/Th2 balance in mice with allergic asthma, we measured the levels of Th1-/Th2-associated cytokines in lung tissues. The Th1 cytokines (i.e., IFN-γ and IL-2) were significantly downregulated, while the levels of inflammatory Th2 cytokines (i.e., IL-4, IL-5, and IL-13) were markedly elevated in OVA-challenged mice compared with the Sham group (Figure 3), indicating that OVA stimulation resulted in the imbalance of Th1/Th2 in mice. BD at the highest concentration (40 mg/kg) effectively upregulated IFN-γ and IL-2, and reduced the production of Th2-associated cytokines in OVA-stimulated mice, implying that BD treatment restored the Th1/Th2 balance in asthma.
Figure 3 Effect of BD on the production of Th1/Th2-associated cytokines in OVA-exposed mice. The pulmonary concentrations of Th1/Th2-associated cytokines in all groups of mice were measured by ELISA, including IFN-γ, IL-2, IL-4, IL-5, and IL-13. *p < 0.05, **p < 0.01 compared with the Sham group; #p < 0.05, ##p < 0.01 compared with the OVA group.
Effect of BD on the number of inflammatory cells in the blood of OVA-treated mice
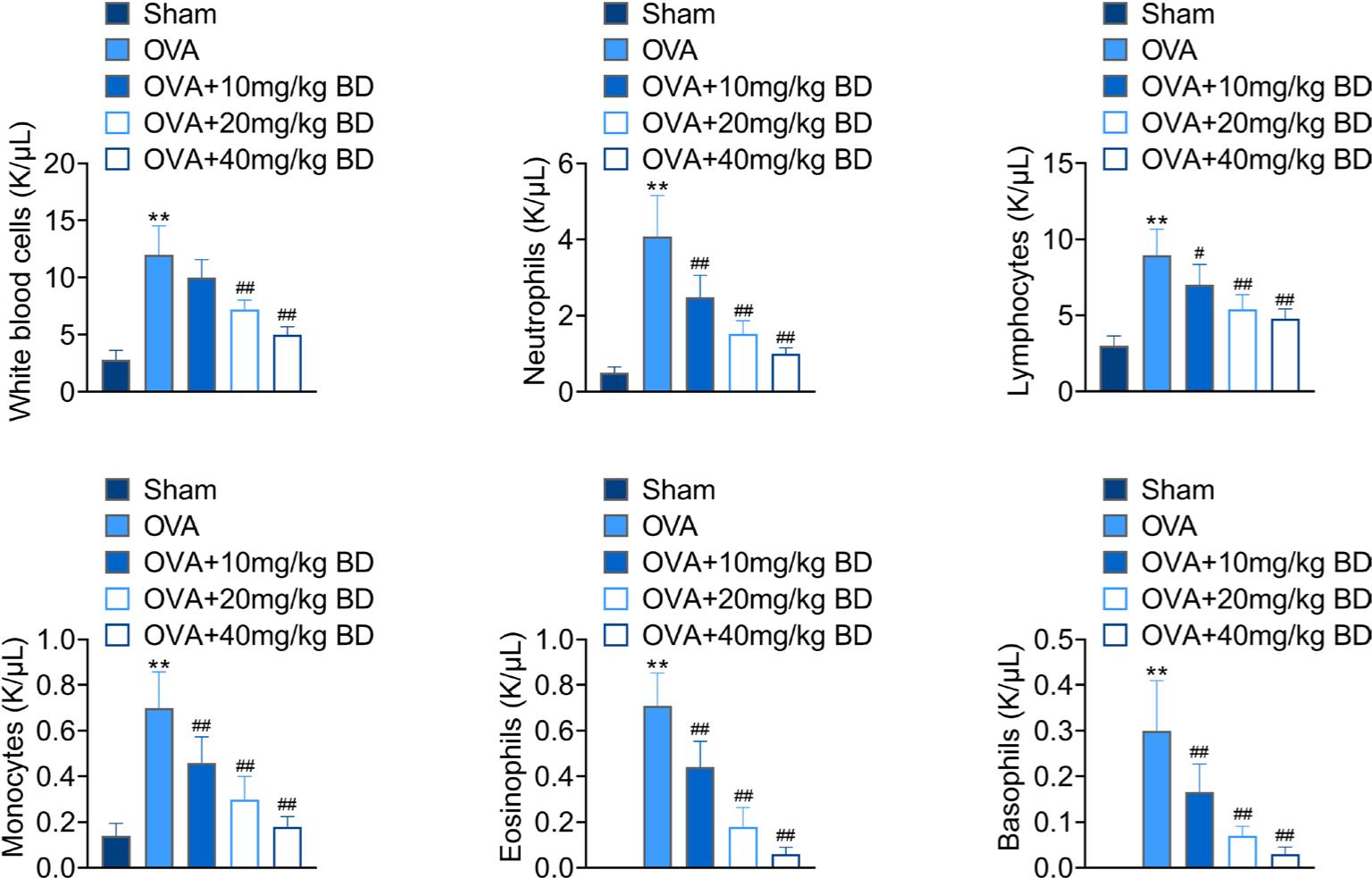
Next, we counted the number of inflammatory cells in the blood of mice to determine the regulatory effect of BD on inflammatory cell population. OVA stimulation significantly increased the number of white blood cells, monocytes, basophils, neutrophils, lymphocytes, and eosinophils in the blood of mice compared with the Sham group. BD treatment, however, almost completely eliminated the effect of OVA challenge on the induction of inflammatory cells (Figure 4). These data suggested that BD inhibited the accumulation of inflammatory cells in mice following OVA stimulation.
Figure 4 Effect of BD on the number of inflammatory cells in OVA-challenged mice. The serum samples were collected from all mice after the AHR assessment. The numbers of inflammatory cells, including white blood cells, neutrophils, lymphocytes, monocytes, eosinophils, and basophils in the blood were counted by a hemocytometer. **p < 0.01 compared with the Sham group; #p < 0.05, ##p < 0.01 compared with the OVA group.
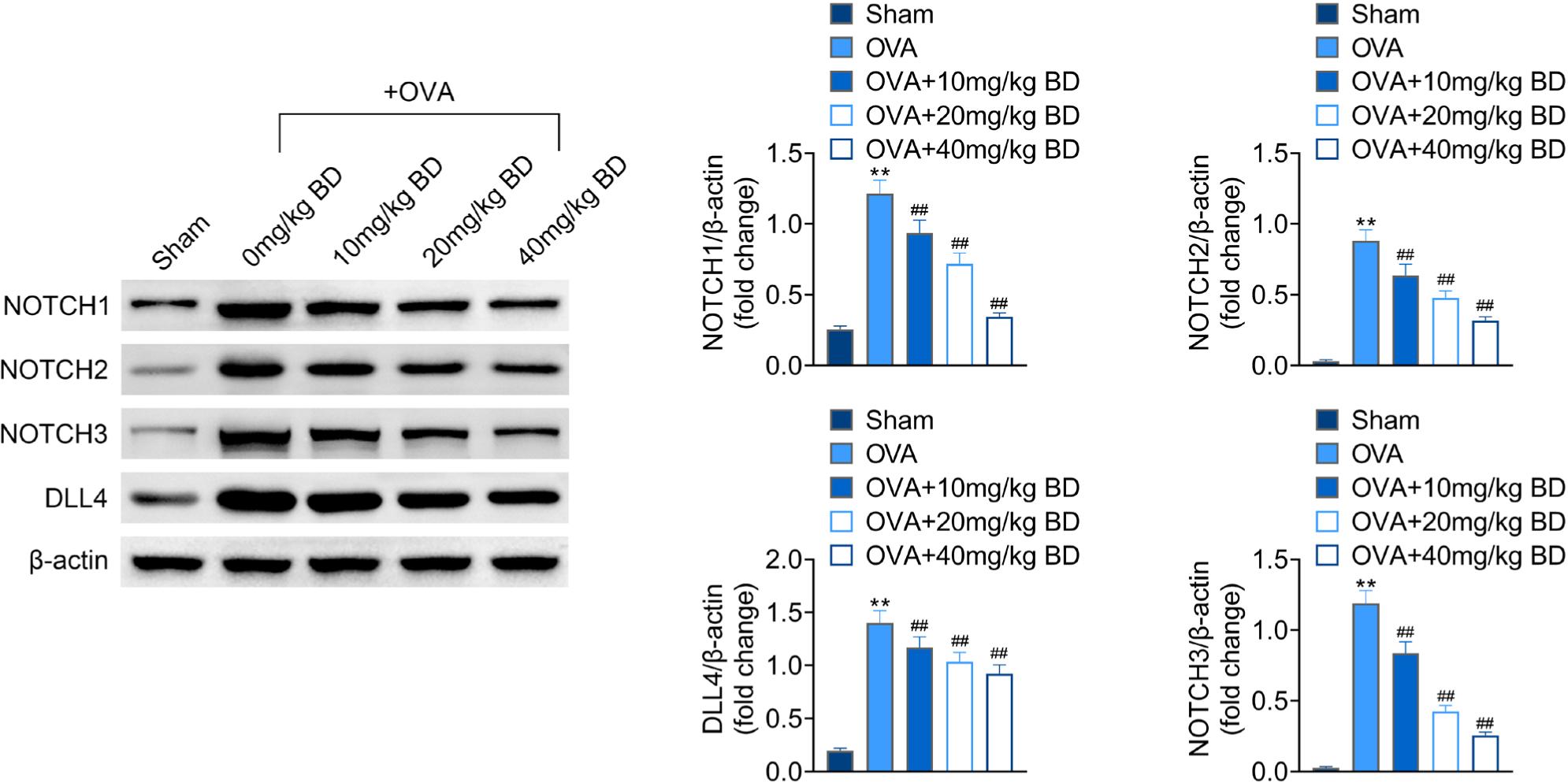
Effect of BD on OVA-induced activation of the NOTCH pathway
Lastly, we found that OVA challenge induced the activation of the NOTCH pathway in mice, as evidenced by significantly upregulated pulmonary expressions of NOTCH1, NOTCH2, NOTCH3, and DLL4, a transmembrane protein that acts as a ligand for Notch receptors. Treatment of BD at all doses efficiently decreased the levels of all NOTCH-signaling-related proteins in a concentration-dependent manner (Figure 5), indicating that BD restrained the activation of the NOTCH signaling in OVA-challenged mice.
Figure 5 Effect of BD on OVA-induced activation of the NOTCH pathway. The expressions of NOTCH1, NOTCH2, NOTCH3, and DLL4 in mouse lung tissues were examined by Western blot. Beta-actin was used as an internal control. **p < 0.01 compared with the Sham group; ##p < 0.01 compared with the OVA group.
Discussion
Despite major advances in the treatment of allergic asthma over the past decades, it remains a serious health problem that affects at least 300 million people worldwide.19 The potential use of pharmaceutical compounds to reduce bronchial inflammation and inhibit AHR in asthma has been explored. In the present study, we showed that Bruceine D restored the Th1/Th2 balance and inhibited inflammatory cell infiltration in OVA-induced allergic asthma mice via suppressing the NOTCH pathway.
In allergic asthma, airway structural changes, such as the loss of epithelial integrity, subepithelial fibrosis, increased smooth muscle mass, and neovascularization, have been shown to cause airway narrowing, AHR, and infiltration of inflammatory cells.20 In this study, histological analysis showed that BD ameliorated OVA-induced inflammatory cell infiltration into the perivascular and peribronchial tissues and reduced fibrosis around these areas. Bronchial hyperresponsiveness, referring to the predisposition of the airway to narrow excessively in response to stimuli, has been considered a major hallmark of allergic asthma.21 Previous evidence has suggested that the severity of AHR is associated with an increased level of treatment required for asthma control and an increased risk of life-threatening exacerbation.22 Here, we found that BD treatment significantly and dose-dependently decreased OVA-induced AHR in mice.
The imbalance of Th1 and Th2 cells is a key contributor to the pathogenesis of allergic asthma.23 The inappropriate immune responses to allergens trigger the differentiation of naïve T CD4+ cells toward Th2 cells and consequently leads to the excessive secretion of Th2-associated cytokines, particularly IL-4, IL-5, and IL-13, which induce AHR, mucus hypersecretion, and goblet cell hyperplasia.24 The Th1-associated cytokines (i.e., IFN-γ and IL-2), however, have been reported to inhibit the expression of IL-4 and reduce the aggregation of immune cells.25 In this study, mice treated with BD showed significantly downregulated Th2-associated cytokines and elevated production of Th1-associated cytokines following OVA stimulation, suggesting that BD intervention restored the Th1/Th2 balance in asthmatic mice.
Inflammatory cells, including white blood cells, basophils, neutrophils, monocytes, lymphocytes, and eosinophils, are essential components of allergic inflammation. For example, neutrophils produce excessive amounts of inflammatory cytokines, thus contributing to airway remodeling and exaggerating asthma.26 Lymphocytes are involved in the regulation of airway immune response in asthma and secrete proinflammatory factors.27 Eosinophils release amyloid granules at the site of inflammation to induce airway obstruction and tissue edema, and stimulate the secretion of proinflammatory cytokines, which further promote bronchial inflammation.28 Here, our results demonstrated that BD treatment dose-dependently inhibited OVA-induced accumulation of inflammatory cells in asthmatic mice. Further investigations of inflammatory cell count in the lung tissues are needed to validate the current findings.
The activation of the NOTCH pathway plays an indispensable role in allergic asthma. Inhibition of NOTCH signaling in CD4+ T cells protected mice against allergic airway inflammation and the development of AHR.29 It has also been reported that T-cell-specific NOTCH deficiency prevents house-dust-mite-driven eosinophilic bronchial inflammation and reduces the production of Th2 cytokines and AHR.30 Our data showed that OVA exposure increased pulmonary expressions of all NOTCH signaling receptors, while BD treatment significantly abolished OVA-induced activation of the NOTCH pathway.
Conclusion
Taken together, BD protected mice against OVA-induced allergic asthma by reducing AHR and restoring the balance between Th1- and Th2-associated cytokines through deactivating the NOTCH pathway. Our findings suggest that BD may be used as a therapeutic agent for allergic asthma. Future investigations are needed to determine the optimal dosage and delivery method of BD.
Acknowledgements
Not applicable.
Funding
This work was supported by Wuhan Municipal Health Commission Project (Grant No. WX19C15).
Conflicts of Interest
The authors state that there are no conflicts of interest to disclose.
Ethics approval
Ethical approval was obtained from the Animal Ethics Committee of Wuhan Myhalic Biotechnology Co., Ltd. (Approval no. HLK-20210202-01).
Availability of data and materials
All data generated or analyzed during this study are included in this article.
Authors’ contributions
Ying Nie and Bangkun Yang designed the study, supervised data collection; Junfeng Hu and Lingling Zhang analyzed and interpreted the data; Zhimin Ma prepared the manuscript for publication and reviewed the draft of the manuscript. All authors have read and approved the manuscript.
REFERENCES
1. Quirt J, Hildebrand KJ, Mazza J, Noya F, Kim H. Asthma. Allergy Asthma Clin Immunol. 2018;14(Suppl 2):50. 10.1186/s13223-018-0279-0
2. Backman H, Räisänen P, Hedman L, Stridsman C, Andersson M, Lindberg A, et al. Increased prevalence of allergic asthma from 1996 to 2006 and further to 2016-results from three population surveys. Clin Exp Allergy. 2017;47(11):1426–1435. 10.1111/cea.12963
3. Pakkasela J, Ilmarinen P, Honkamäki J, Tuomisto LE, Andersén H, Piirilä P, et al. Age-specific incidence of allergic and non-allergic asthma. BMC Pulm Med. 2020;20(1):9. 10.1186/s12890-019-1040-2
4. Ramadan AA, Gaffin JM, Israel E, Phipatanakul W. Asthma and corticosteroid responses in childhood and adult asthma. Clin Chest Med. 2019;40(1):163–177. 10.1016/j.ccm.2018.10.010
5. Kim YM, Kim YS, Jeon SG, Kim YK. Immunopathogenesis of allergic asthma: more than the th2 hypothesis. Allergy Asthma Immunol Res. 2013;5(4):189–196. 10.4168/aair.2013.5.4.189
6. Azman S, Sekar M, Bonam SR, Gan SH, Wahidin S, Lum PT, et al. Traditional medicinal plants conferring protection against ovalbumin-induced asthma in experimental animals: a review. J Asthma Allergy. 2021;14:641–662. 10.2147/JAA.S296391
7. León B, Ballesteros-Tato A. Modulating Th2 cell immunity for the treatment of asthma. Front Immunol. 2021;12:637948. 10.3389/fimmu.2021.637948
8. Boonpiyathad T, Sözener ZC, Satitsuksanoa P, Akdis CA. Immunologic mechanisms in asthma. Semin Immunol. 2019;46:101333. 10.1016/j.smim.2019.101333
9. Koch S, Finotto S. Role of interferon-λ in allergic asthma. J Innate Immun. 2015;7(3):224–230. 10.1159/000369459
10. Subeki, Matsuura H, Takahashi K, Nabeta K, Yamasaki M, Maede Y, et al. Screening of Indonesian medicinal plant extracts for antibabesial activity and isolation of new quassinoids from Brucea javanica. J Nat Products. 2007;70(10):1654–1657. 10.1021/np070236h
11. Fan J, Ren D, Wang J, Liu X, Zhang H, Wu M, et al. Bruceine D induces lung cancer cell apoptosis and autophagy via the ROS/MAPK signaling pathway in vitro and in vivo. Cell Death Dis. 2020;11(2):126. 10.1038/s41419-020-2317-3
12. Li L, Dong Z, Shi P, Tan L, Xu J, Huang P, et al. Bruceine D inhibits cell proliferation through downregulating LINC01667/MicroRNA-138-5p/Cyclin E1 axis in gastric cancer. Front Pharmacol. 2020;11:584960. 10.3389/fphar.2020.584960
13. Luo C, Wang Y, Wei C, Chen Y, Ji Z. The anti-migration and anti-invasion effects of Bruceine D in human triple-negative breast cancer MDA-MB-231 cells. Exp Ther Med. 2020;19(1):273–279. 10.3892/etm.2019.8187
14. Yang Y, Kong F, Ding Q, Cai Y, Hao Y, Tang B. Bruceine D elevates Nrf2 activation to restrain Parkinson’s disease in mice through suppressing oxidative stress and inflammatory response. Biochem Biophys Res Commun. 2020;526(4):1013–1020. 10.1016/j.bbrc.2020.03.097
15. Cheng Z, Yuan X, Qu Y, Li X, Wu G, Li C, et al. Bruceine D inhibits hepatocellular carcinoma growth by targeting β-catenin/jagged1 pathways. Cancer Lett. 2017;403:195–205. 10.1016/j.canlet.2017.06.014
16. Hua S, Liu F, Wang M. Emodin alleviates the airway inflammation of cough variant asthma in mice by regulating the notch pathway. Med Sci Monit. 2019;25:5621–5629. 10.12659/MSM.915080
17. National Research Council (US) Committee for the Update of the Guide for the Care and Use of Laboratory Animals. Guide for the Care and Use of Laboratory Animals. 8th ed. Washington (DC): National Academies Press (US); 2011. PMid: 21595115. https://pubmed.ncbi.nlm.nih.gov/21595115/
18. Tang Y, Huang W, Song Q, Zheng X, He R, Liu J. Paeonol ameliorates ovalbumin-induced asthma through the inhibition of TLR4/NF-κB and MAPK signaling. Evid Based Complement Alternat Med. 2018;2018:3063145. 10.1155/2018/3063145
19. D’Amato G, Vitale C, Molino A, Stanziola A, Sanduzzi A, Vatrella A, et al. Asthma-related deaths. Multidiscip Respir Med. 2016;11:37. 10.1186/s40248-016-0073-0
20. Bergeron C, Tulic MK, Hamid Q. Airway remodelling in asthma: from benchside to clinical practice. Can Respir J. 2010;17(4):e85–93. 10.1155/2010/318029
21. Brannan JD, Lougheed MD. Airway hyperresponsiveness in asthma: mechanisms, clinical significance, and treatment. Front Physiol. 2012;3:460. 10.3389/fphys.2012.00460
22. Chapman DG, Irvin CG. Mechanisms of airway hyper-responsiveness in asthma: the past, present and yet to come. Clin Exp Allergy. 2015;45(4):706–719. 10.1111/cea.12506
23. Kudo M, Ishigatsubo Y, Aoki I. Pathology of asthma. Front Microbiol. 2013;4:263. 10.3389/fmicb.2013.00263
24. Caminati M, Pham DL, Bagnasco D, Canonica GW. Type 2 immunity in asthma. World Allergy Organ J. 2018;11(1):13. 10.1186/s40413-018-0192-5
25. Raphael I, Nalawade S, Eagar TN, Forsthuber TG. T cell subsets and their signature cytokines in autoimmune and inflammatory diseases. Cytokine. 2015;74(1):5–17. 10.1016/j.cyto.2014.09.011
26. Ray A, Kolls JK. Neutrophilic inflammation in asthma and association with disease severity. Trends Immunol. 2017;38(12):942–954. 10.1016/j.it.2017.07.003
27. Larché M, Robinson DS, Kay AB. The role of T lymphocytes in the pathogenesis of asthma. J Allergy Clin Immunol. 2003;111(3):450–463; quiz 64. 10.1067/mai.2003.169
28. Patel TR, Sur S. IgE and eosinophils as therapeutic targets in asthma. Curr Opin Allergy Clin Immunol. 2017;17(1):42–49. 10.1097/ACI.0000000000000336
29. Okamoto M, Matsuda H, Joetham A, Lucas JJ, Domenico J, Yasutomo K, et al. Jagged1 on dendritic cells and Notch on CD4+ T cells initiate lung allergic responsiveness by inducing IL-4 production. J Immunol. 2009;183(5):2995–3003. 10.4049/jimmunol.0900692
30. Tindemans I, van Schoonhoven A, KleinJan A, de Bruijn MJ, Lukkes M, van Nimwegen M, et al. Notch signaling licenses allergic airway inflammation by promoting Th2 cell lymph node egress. J Clin Investig. 2020;130(7):3576–3591. 10.1172/JCI128310