
Download
ORIGINAL ARTICLE
Mechanism of Rhizoma Coptidis in epilepsy with network pharmacology
Dan Huang#, Yan Lv#, Chuansen Lu, Bo Zhang, Zongjun Fu, Yingliu Huang*
Department of Neurology, Hainan General Hospital, Hainan Affiliated Hospital of Hainan Medical University, Haikou, China
Abstract
Network pharmacology is a bioinformatics-based research strategy aimed at identifying drug actions and facilitating drug discovery. In this study, network pharmacology was used for exploring the anti-epileptic multi-target mechanism of Rhizoma Coptidis. The possible protein targets of Rhizoma Coptidis were predicted by constructing the pathway and network of drug targets. Then, the interaction of the main active components of Rhizoma Coptidis and predicted candidate targets were verified using molecular docking technology. Finally, nine active compounds were selected from Rhizoma Coptidis. A total of 68 targets associated with Rhizoma Coptidis treating epilepsy. The key targets were AKT1, IL6, VEGFA, and TP53. According to GO functional enrichment analysis, 289 items of biological process, 33 items of cellular component, and 55 items of molecular function were obtained. A total of 89 signaling pathways were identified through KEGG pathway enrichment analysis (P < 0.05), and HIF-1, TNF, and T-cell receptor signaling pathways were mainly related to epilepsy. Molecular docking showed quercetin and (R)-canadine combined well with the key targets. The active ingredient in Rhizoma Coptidis can regulate various signaling pathways, and have therapeutic effects on epilepsy.
Key words: Rhizoma Coptidis, epilepsy, molecular docking, network pharmacology
*Corresponding author: Yingliu Huang. Department of Neurology, Hainan General Hospital, Hainan Affiliated Hospital of Hainan Medical University, Haikou, 570311, China. Email address: [email protected]
#These authors contribute equally to this study.
Received 16 August 2021; Accepted 21 October 2021; Available online 1 May 2022
Copyright: Huang D, et al.
License: This open access article is licensed under Creative Commons Attribution 4.0 International (CC BY 4.0). http://creativecommons.org/licenses/by/4.0/
Introduction
Epilepsy is a long-term central nervous system disease that has a huge impact on the physiological, psychological, cognitive functions of patients.1–3 Its abominability and incidence rate pose a serious threat to the physical and mental health of patients and to the social development.4,5 Its typical manifestation is recurrent seizures, which damage the patients’ psychological, mental, and cognitive functions.6 Globally, more than 70 million people are affected by epilepsy.
Currently, treatment for epilepsy is mainly oral antiepileptic drugs. Most antiepileptic drugs target neurons and achieve an antiepileptic effect by inhibiting glutamate energy transfer or promoting Y-aminobutyric acid energy transfer. The first-line treatment of epilepsy is oral antiepileptic. Its antiepileptic mechanism mostly focuses on inhibiting neural excitability and neuroelectrical activity. However, on the premise of taking antiepileptic drugs in sufficient amount and treatment, 30% of patients still have uncontrollable seizures.8
Discovering novel therapeutic targets and developing therapeutic drugs with short onset latency and few side effects is the primary task of prevention and treatment of epilepsy. Traditional Chinese medicines (TCMs) play an important role in the prevention and treatment of diseases owing to their improved safety and few side effects and thus have attracted attention worldwide.9 TCMs have the following characteristics: multiple ingredients, multiple targets, and multiple ways to be effective.10 As a Chinese herbal medicine, Rhizoma Coptidis is a common Chinese medicine that shows multiple biological activities, including anti-carcinogenic, neuroprotective, and anti-cancer et al.11 Pharmacological research results show that Rhizoma Coptidis medicinal and health care values such as being anti-inflammatory, anti-viral, bacteriostatic, and hypoglycemic. The chemical constituents of Rhizoma Coptidis include alkaloids, lignans, coumarins, flavonoids, terpenoids, steroids, organic acids, volatile oil, and polysaccharides.12 It has a good synergistic effect on epilepsy treatment.13 Owing to the rapid progress of bioinformatics, systems biology, and poly--pharmacology, network-based drug discovery has become a promising approach for developing effective drugs. In 2007, Hopkins et al. proposed the concept of “network pharmacology,” which is a method for analyzing the interventions using drugs and potential treatment targets of diseases. Network pharmacology highlights a paradigm of the “network target, multi-component” strategy.14
In this study, we applied network pharmacology to clarify the mechanism of Rhizoma Coptidis in the treatment of epilepsy. We systematically analyzed the active ingredients, potential targets, pathways, and networks affected by Rhizoma Coptidis for the treatment of epilepsy. We also performed docking studies to predict the interactions that allow important compounds to bind to predicted targets. Our results may help clarify how Rhizoma Coptidis can be effective against epilepsy and facilitate the development of novel drugs.
Materials and Methods
Prediction of target genes associated with epilepsy
Epilepsy-related genes were searched from GeneCards (https://www.genecards.org/) and the National Center for Biotechnology Information database (https://www.ncbi.nlm.nih.gov/), and used epilepsy as the key word. After “Homo sapiens” was used as a filter term, 712 genes were screened by NCBI, and 2088 genes were screened by GeneCards. A total of 2228 genes were obtained after the deletion of repetitive genes.
Collection of components and targets of Rhizoma Coptidis
According to the natural product databases for Chinese herbal medicines: TCMSP (https://tcmsp-e.com/),15 the compounds and targets of Rhizoma Coptidis were collected. Absorption, distribution, metabolism, and excretion related models, including oral bioavailability (OB)16 and drug-likeness (DL),17 were used as the main variables that affect the absorption of drugs by the gastrointestinal tract. Therefore, we screened bioactive components under the standards of OB ≥ 30% and DL ≥ 0.1818, and obtained the related targets of each component. By using a Venn map, we intersected genes associated with epilepsy and the common targets of Rhizoma Coptidis. The intersected genes were probably the target genes of Rhizoma Coptidis in epilepsy treatment.
Protein–protein interaction data
The STRING database (https://string-db.org/cgi/input.pl)19 contains known and predicted protein–protein interactions (PPIs). A large number of PPIs were collected, including 9,643,763 proteins and 1,380,838,440 interactions and data obtained from experimental detection and bioinformatics prediction. The intersected genes were imported into the STRING database. The species was defined as “Homo sapiens.” PPI data were obtained, and the results were saved in TSV format. The information of node1 and node2, and combined score in the file was retained. The interaction network was drawn, and the network was analyzed using Cytoscape.
Network construction
The active components of Rhizoma Coptidis and intersected genes were imported into Cytoscape version 3.6.0 to construct the compound-target network of Rhizoma Coptidis.
Gene ontology and pathway analysis
We used the Database for Annotation, Visualization, and Integrated Discovery20 (DAVID, https://david.ncifcrf.gov/, v6.8) for gene ontology (GO) enrichment analysis, and pathway enrichment analysis was performed using the Kyoto Encyclopedia of Genes and Genomes21 (KEGG, http://www.kegg.jp/). Biological information annotation database (David, https://david.ncifcrf.gov/, version 6.8) provides a systematic and comprehensive annotation information on the biological functions of large-scale genes or proteins and can determine most significantly enriched biological annotation. The intersected genes were imported into the DAVID database. The select identifier was set as official gene symbol. The list type was set to gene list, and the species was limited to “Homo sapiens.” Go analysis and KEGG pathway analysis were performed based on the intersected genes.
Molecular docking
The active components were selected as the ligand, and the targets with the top four-degree values was selected as the receptors for molecular docking. (1) Ligand preparation: ligand molecular structure was downloaded from the TCMSP database and saved in mol2 format. (2) Preparation of receptors: the molecular structures of the receptors were downloaded from the RCSB PDB database, and the molecules with high reliability were selected and stored in PDB format. (3) Molecular docking: Autodock was used in dehydrating, hydrogenating, and computing charge, the results were saved in pdbqt format for molecular docking, and Discovery Studio 2016 was used for visualization. The selected target proteins was AKT1 (2uzs),22 VEGFA (6d3o: https://www.pdbus.org/structure/), IL6 (1alu),23 and TP53(3dcy: https://www.pdbus.org/structure/3DCY).
Results
Active compounds of Rhizoma Coptidis
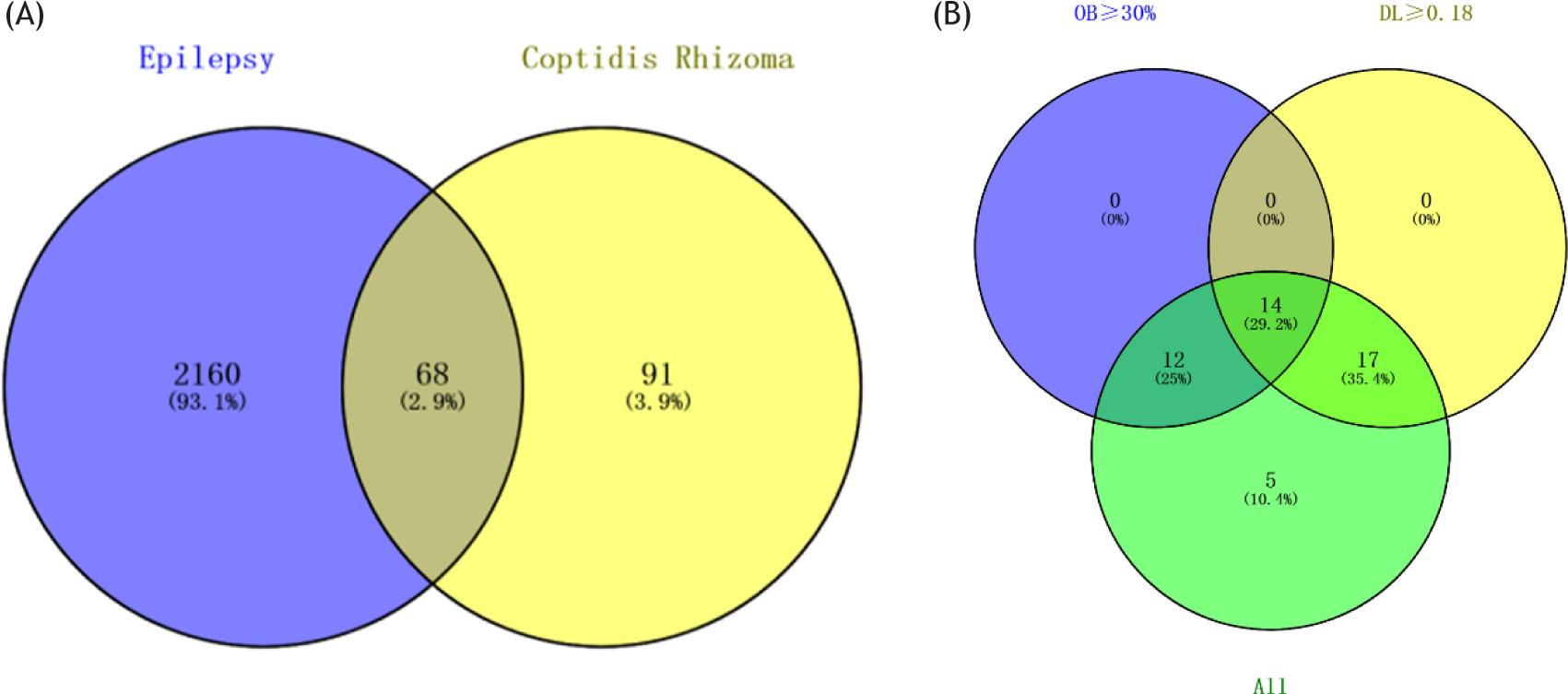
TCMSP database (http://ibts.hkbu.edu.hk/LSPtcmsp.php) is a unique pharmacological platform of the TCM system, which can calculate ADME-related characteristics of natural compounds.15 A total of 49 components of Rhizoma Coptidis were identified from TCMSP. The components were screened with the following criteria: OB ≥ 30% and DL ≥ 0.18. In this study, 14 bioactive components of Rhizoma Coptidis were screened, five of which had no targets. Finally, nine compounds were collected from the TCMSP database (Figure 1B; Table 1).
Figure 1 Rhizoma Coptidis screening of bioactive compounds and targets. (A) Mapping of Rhizoma Coptidis–related targets and epilepsy-related targets, 68 common targets were showed. (B) Venn diagram: 48 components (green section), and 14 bioactive components screened by two ADME-related models (blue section stands for the components of OB≥30%, yellow section stands for DL≥ 0.18).
Table 1 Basic information of active compounds in Rhizoma Coptidis.
| No | Mol ID | Molecule name | MW | OB (%) | DL |
|---|---|---|---|---|---|
| 1 | MOL001454 | berberine | 336.39 | 36.86 | 0.78 |
| 2 | MOL002894 | berberrubine | 322.36 | 35.74 | 0.73 |
| 3 | MOL002903 | (R)-Canadine | 339.42 | 55.37 | 0.77 |
| 4 | MOL002904 | Berlambine | 351.38 | 36.68 | 0.82 |
| 5 | MOL002907 | Corchoroside A_qt | 404.55 | 104.95 | 0.78 |
| 6 | MOL000622 | Magnograndiolide | 266.37 | 63.71 | 0.19 |
| 7 | MOL000785 | palmatine | 352.44 | 64.6 | 0.65 |
| 8 | MOL000098 | quercetin | 302.25 | 46.43 | 0.28 |
| 9 | MOL002668 | Worenine | 334.37 | 45.83 | 0.87 |
Table 2 Information of potential anti-epilepsy targets from Rhizoma Coptidis.
| Serial number | Name | Degree | Serial number | Name | Degree |
|---|---|---|---|---|---|
| 1 | CHRM3 | 1 | 35 | CYP3A4 | 22 |
| 2 | SCN5A | 2 | 36 | F2 | 25 |
| 3 | GABRA1 | 3 | 37 | NFE2L2 | 26 |
| 4 | NR3C2 | 4 | 38 | HIF1A | 26 |
| 5 | INSR | 5 | 39 | AHR | 27 |
| 6 | GRIA2 | 5 | 40 | CRP | 27 |
| 7 | KCNH2 | 5 | 41 | VCAM1 | 27 |
| 8 | NR1I3 | 7 | 42 | MMP3 | 28 |
| 9 | SLC6A3 | 7 | 43 | IL2 | 29 |
| 10 | TOP2A | 7 | 44 | IFNG | 29 |
| 11 | MAOB | 8 | 45 | BCL2L1 | 31 |
| 12 | POR | 8 | 46 | AR | 31 |
| 13 | DRD1 | 9 | 47 | ERBB2 | 32 |
| 14 | BAX | 9 | 48 | SPP1 | 33 |
| 15 | HTR3A | 10 | 49 | HMOX1 | 33 |
| 16 | GSTM1 | 10 | 50 | CCL2 | 33 |
| 17 | E2F1 | 11 | 51 | IL10 | 33 |
| 18 | NR1I2 | 11 | 52 | SERPINE1 | 33 |
| 19 | SLC6A4 | 11 | 53 | MMP2 | 35 |
| 20 | BCL2 | 11 | 54 | IL1B | 36 |
| 21 | CYP1A2 | 13 | 55 | MMP9 | 38 |
| 22 | OPRM1 | 13 | 56 | ESR1 | 39 |
| 23 | ACHE | 15 | 57 | MYC | 39 |
| 24 | CTSD | 15 | 58 | PTGS2 | 39 |
| 25 | RAF1 | 15 | 59 | MAPK1 | 39 |
| 26 | PLAT | 15 | 60 | EGFR | 40 |
| 27 | GSTP1 | 17 | 61 | FOS | 40 |
| 28 | CYP1A1 | 18 | 62 | CXCL8 | 41 |
| 29 | IL1A | 18 | 63 | TNF | 42 |
| 30 | GJA1 | 19 | 64 | CASP3 | 43 |
| 31 | HSPB1 | 20 | 65 | VEGFA | 45 |
| 32 | CD40LG | 20 | 66 | IL6 | 45 |
| 33 | IGFBP3 | 21 | 67 | TP53 | 47 |
| 34 | ABCG2 | 22 | 68 | AKT1 | 50 |
Epilepsy–Rhizoma Coptidis PPI network
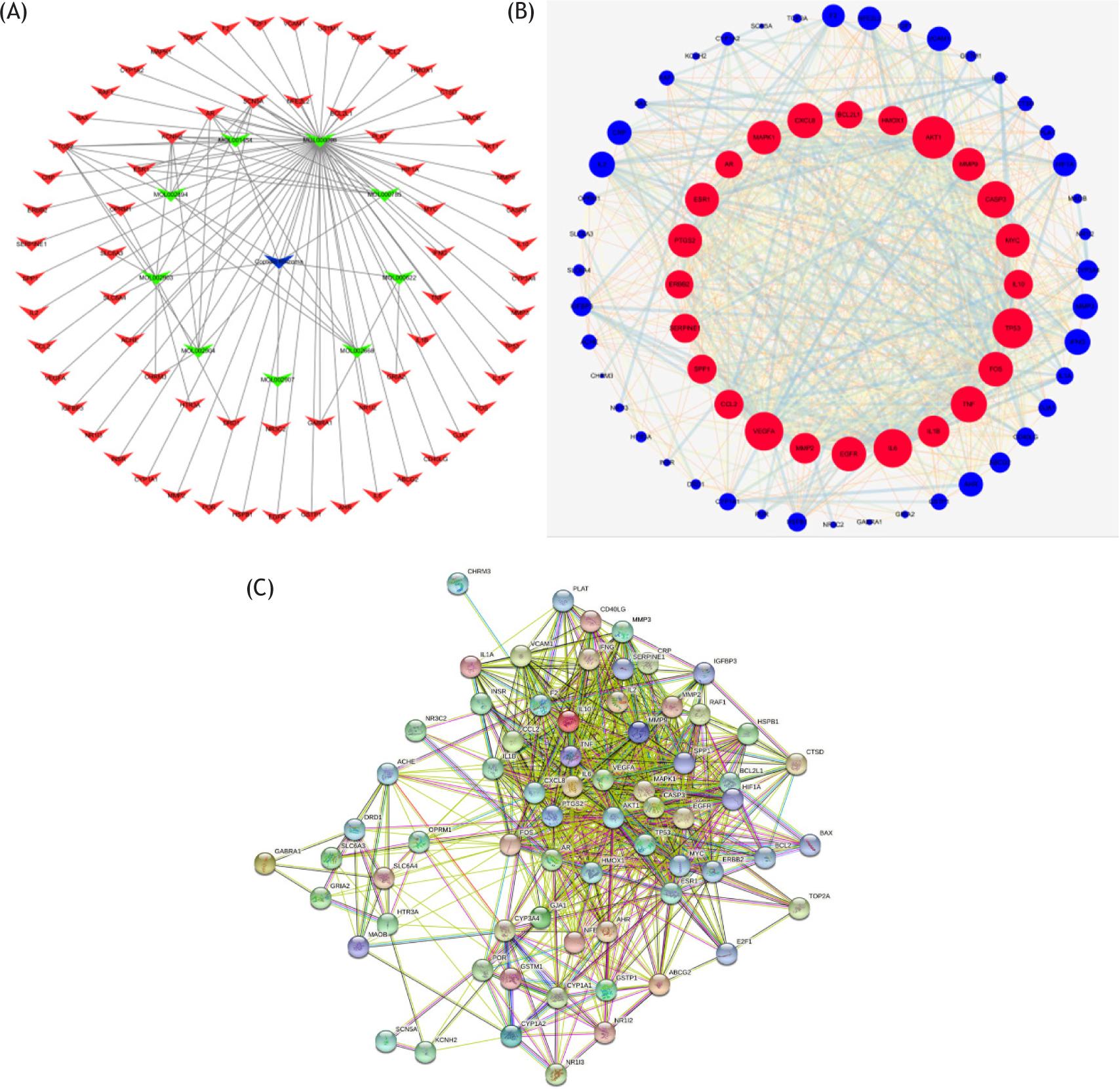
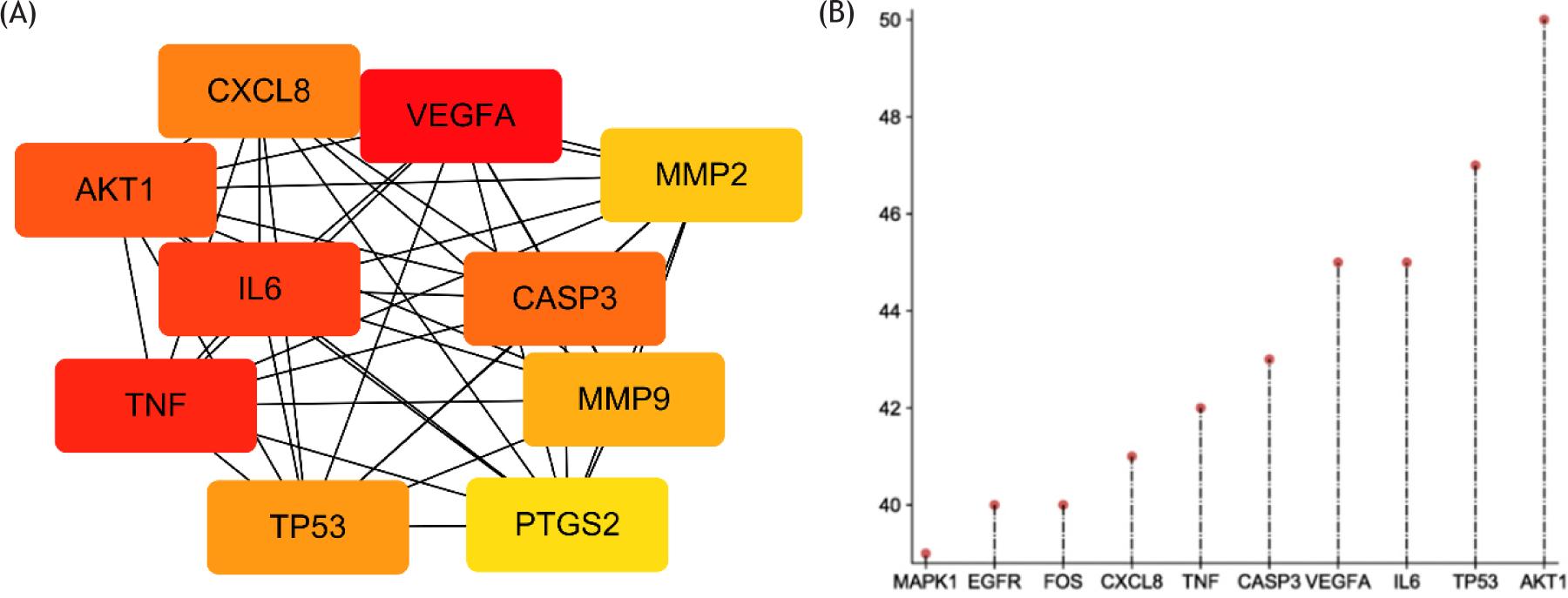
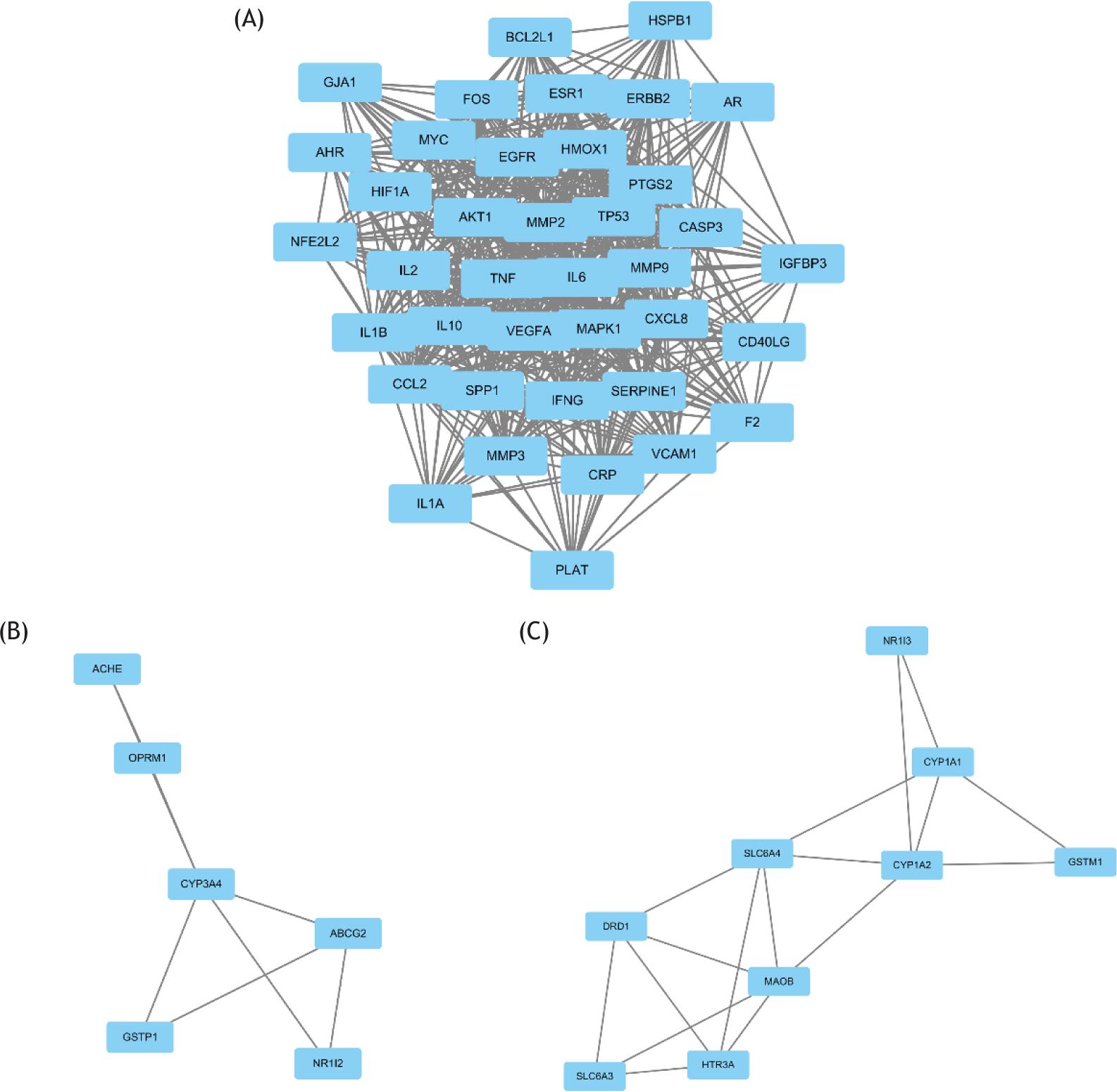
By using epilepsy as a keyword in NCBI and GeneCards database, 2228 genes related to depression were retrieved, and 159 target genes of epilepsy were collected from the TCMSP database. After the target genes of epilepsy and epilepsy were intersected, 68 common genes were found (Figure 1A). These genes can be the target genes of Rhizoma Coptidis in the treatment of epilepsy. Then, we built a visualized epilepsy–Rhizoma Coptidis–-component-target network with Cytoscape 3.6. The nodes of different colors and shapes represented the potential active components and targets. The edges represented the correlation between the active components and the targets (Figure 2A) and fully reflected the multi-component and multi-target characteristics of Rhizoma Coptidis. A total of 68 nodes and 784 edges were obtained, as shown in Figure 2B, and the average node degree was 23.1. The local clustering coefficient was 0.707. The size of the node represented the degree value of the targets. The degree value increased with node size. The thickness of the edge indicated the combination score. The combination score value increased with coarseness. The original protein interaction diagram of the STRING database was shown in Figure 2C. The top 10 target proteins were shown in Figure 3A, and the degree of protein was shown in Figure 3B. The three subnetworks were shown in Figure 4.
Figure 2 Common-target network. (A) Construction of Rhizoma Coptidis component-target visual network. Blue node stands for Rhizoma Coptidis, green nodes stand for bioactive components, and red nodes stand for predicted targets. (B) Interaction network of common target proteins. The red nodes represent the big hub nodes. The node size was proportional to the target degree in the network. (C) Protein–protein interaction mapping by string database.
Figure 3 Top 10 hub targets. (A) The top 10 targets screened out. (B) Degree level of each hub-target.
Figure 4 Subnetwork graph of protein–protein interaction. (A) Score = 29.312. (B) Score = 3.2. (C) Score = 4.25.
GO analysis of genes targeted by Rhizoma Coptidis
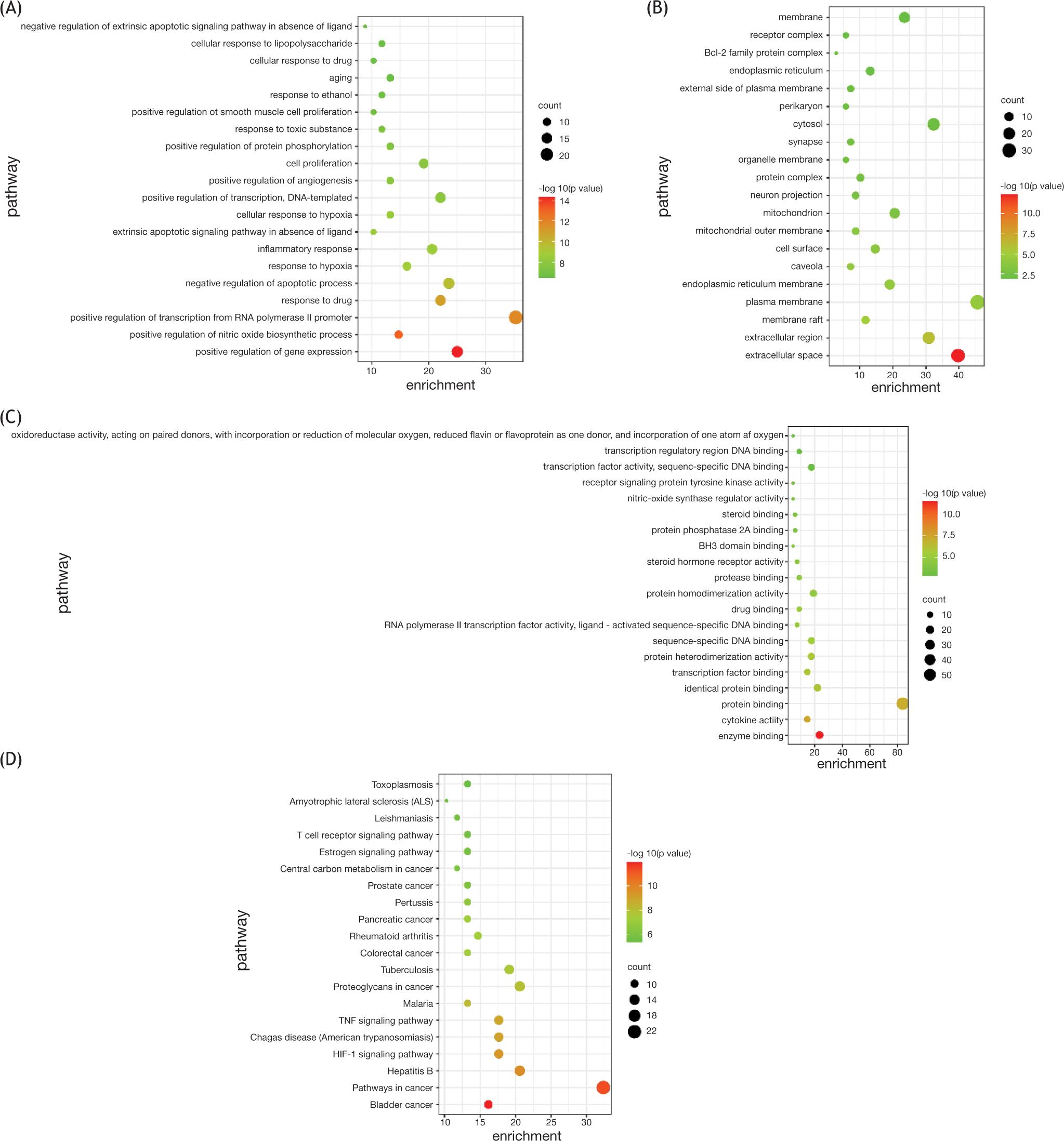
DAVID database was applied to the analysis of the related targets of Rhizoma Coptidis in the treatment of epilepsy. In biological process analysis, the top-ranked targets were positive regulation of gene expression (P = 0.00000000000000434, 17 targets/262), positive regulation of nitric oxide biosynthetic process (P = 0.0000000000000743, 10 targets/43), positive regulation of transcription from RNA polymerase II promoter (P = 0.0000000000015, 24 targets/981), response to drug (P = 0.0000000000125, 15 targets/304) and negative regulation of apoptotic process (P = 0.000000000236, 16 targets/455; Figure 5A). In cellular component analysis (Figure 5B), extracellular space (P = 0.000000000000429, 27 targets/1347), extracellular region (P = 0.00000075, 21 targets/1610), membrane raft (P = 0.0000104, 8 targets/206), plasma membrane (P = 0.0000474, 31 targets/4121), extracellular region, and extracellular space were at the top. In molecular function analysis (Figure 5C), enzyme binding (16 targets/P = 0.00000000000266, 333), cytokine activity (10 targets/P = 0.0000000302, 176), protein binding (57 targets/P = 0.0000000617, 8785), identical protein binding (15 targets/P = 0.00000111, 749) were at the top.
Figure 5 Enrichment of GO and KEGG pathway of Rhizoma Coptidis in the treatment of epilepsy. (A) Enriched gene ontology terms for biological process (BP). (B) Enriched gene ontology terms for cellular component. (C) Enriched gene ontology terms for molecular function (MF). (D) Enriched KEGG pathways of potential anti-epilepsy targets from main active ingredients of Rhizoma Coptidis.
KEGG analysis of proteins targeted by Rhizoma Coptidis
DAVID database was used in analyzing the KEGG pathway of the 68 targets of Rhizoma Coptidis for the treatment of epilepsy. According to the P value, the top 20 pathways were selected, which are shown in Figure 5D. The colors of the nodes in the figure (from green to red) reflected the P value from large to small. The nodes from small to large reflected the number of related genes in an increasing trend. The top-ranked pathways were bladder cancer (P = 0.00000000000106, 11 targets/41), pathways in cancer (P = 0.00000000000471, 22 targets/393), hepatitis B (P = 0.000000000209, 14 targets/145), HIF-1 signaling pathway (P = 0.000000000426, 12 targets/96), and Chagas disease (P = 0.00000000102, 12 targets/104). These results indicated that Rhizoma Coptidis role in the treatment of epilepsy by participating in the regulation of a variety of pathways.
Molecular docking
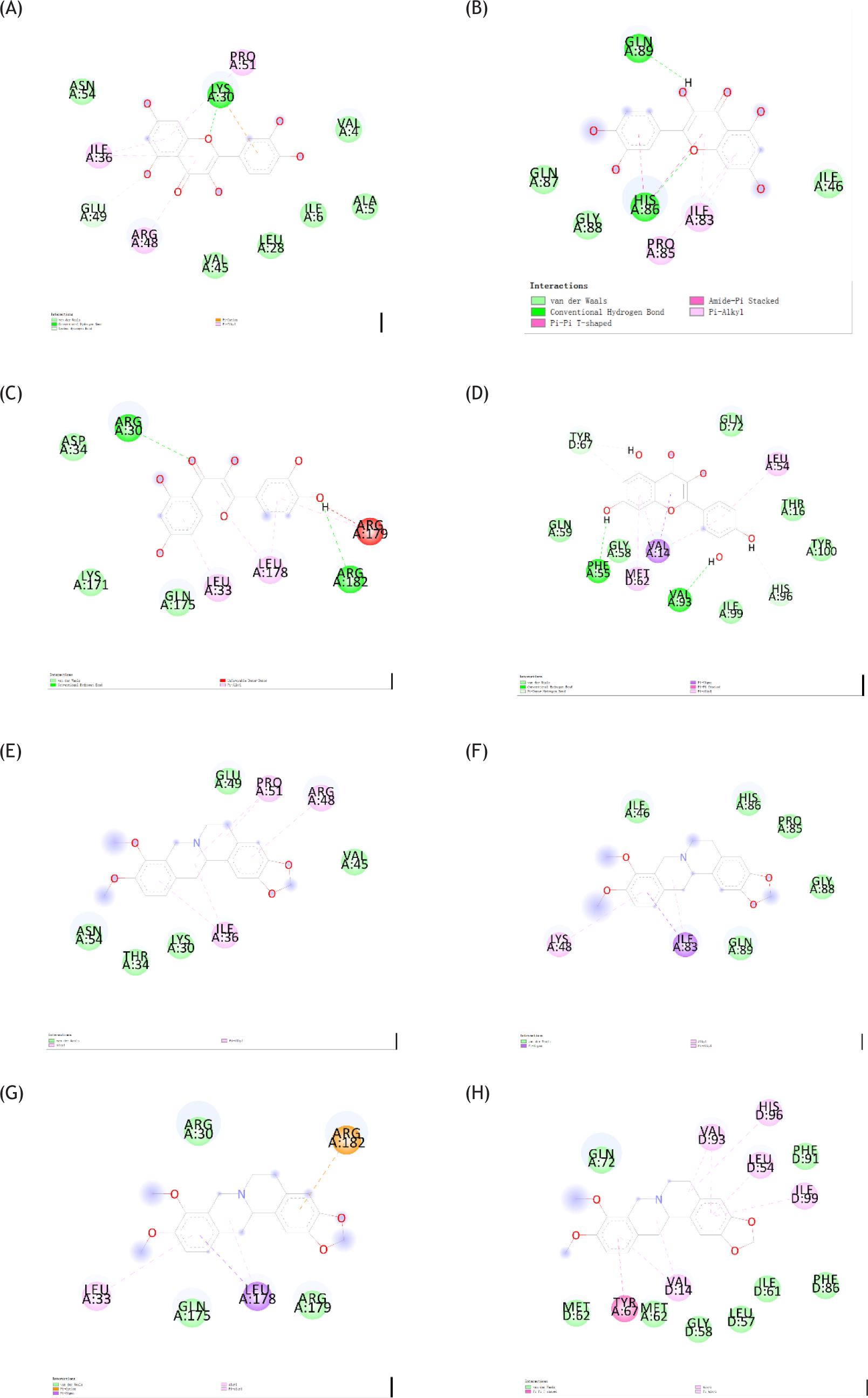
Molecular docking analysis provided a visual explanation of the interaction between ligand and its potential protein targets associated with epilepsy (Figure 6–8). We found that hydrogen bonding and Pi-alkyl were the main forms of interaction. The binding energy and RMSD between ligand and receptor were shown in Tables 3 and 4.
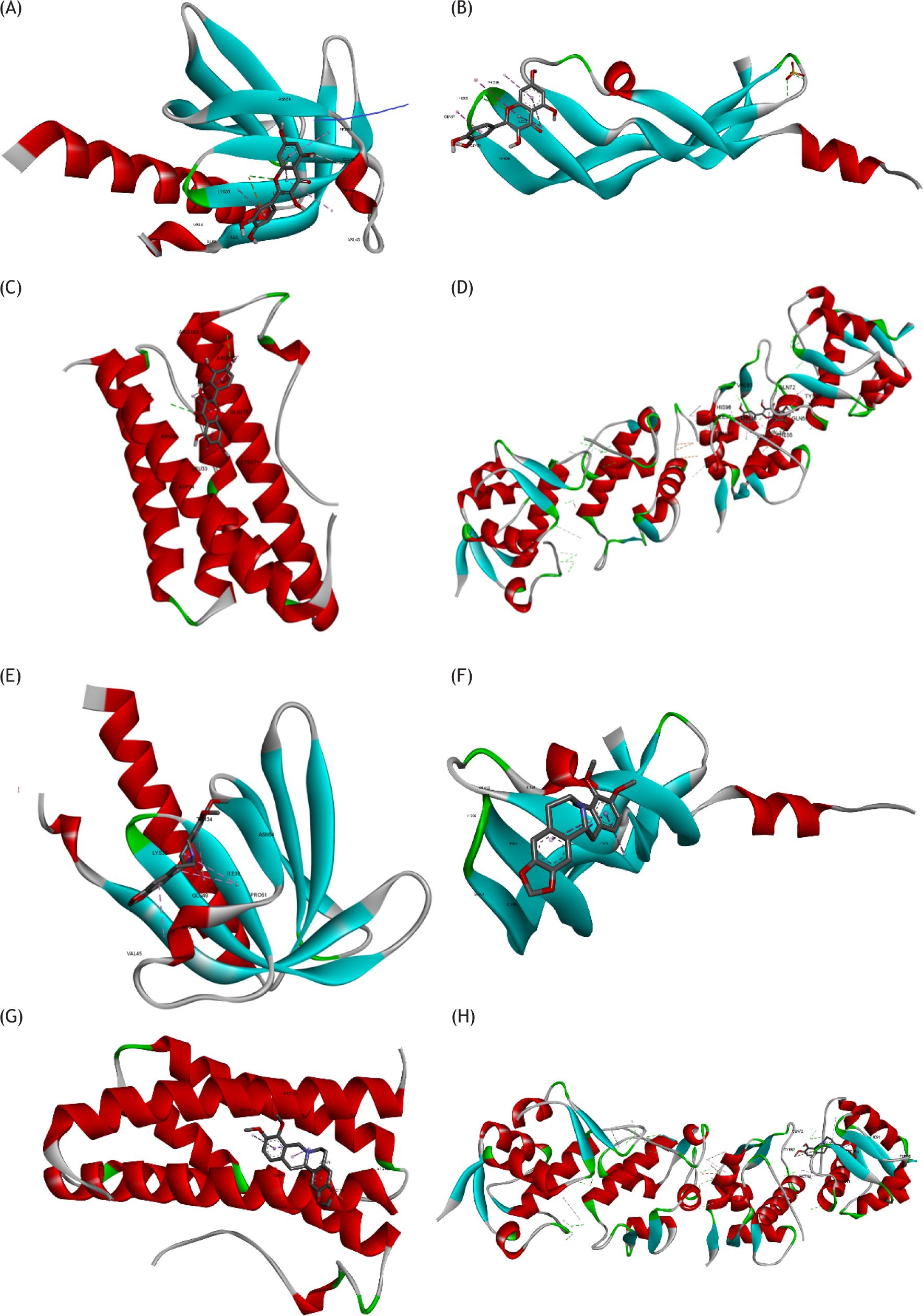
Figure 6 Structural model of active ingredients with hub targets. These four targets were AKT1, VEGFA, IL6, and TP53. The top two compounds were quercetin and (R)-canadine. The binding energy ranges from −7.8 to −4.4 kcal/mol. (A) Structural model of AKT1 with quercetin. (B) Structural model of VEGFA with quercetin. (C) Structural model of IL6 with quercetin. (D) Structural model of TP53 with quercetin. (E) Structural model of AKT1 with (R)-canadine. (F) Structural model of VEGFA with (R)-canadine. (G) Structural model of IL6 with (R)-canadine. (H) Structural model of TP53 with (R)-canadine.
Figure 7 Binding mode of proteins and different ligands. (A) Binding mode of AKT1 with quercetin. (B) Binding mode 1 of VEGFA with quercetin. (C) Binding mode of IL6 with quercetin. (D) Binding mode of TP53 with quercetin. (E) Binding mode of AKT1 with (R)-canadine. (F) Binding mode of VEGFA with (R)-canadine. (G) Binding mode of IL6 with (R)-canadine. (H) Binding mode of TP53 with (R)-canadine.
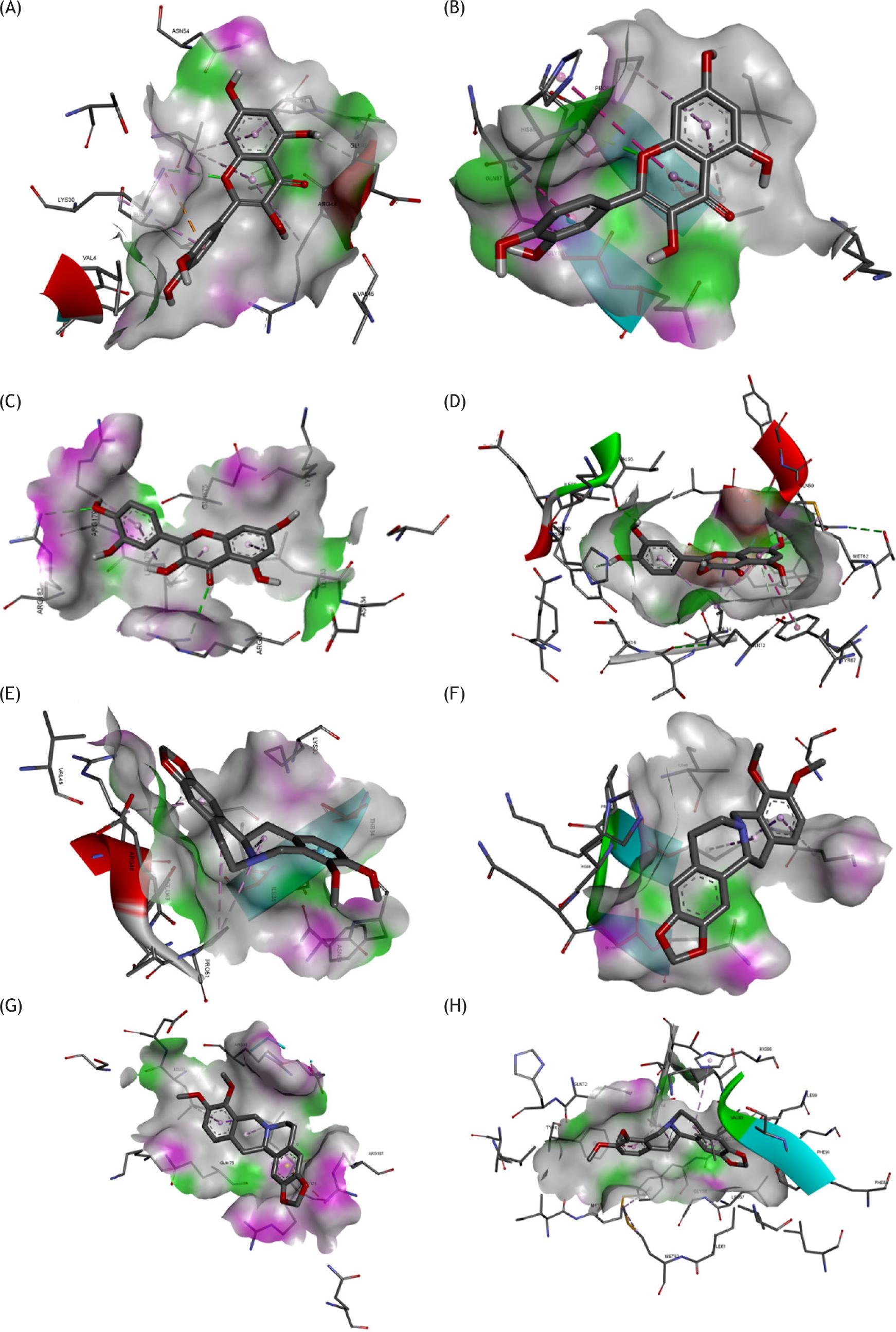
Figure 8 Binding site of active ingredients with hub targets. (A) Binding site of AKT1 with quercetin. (B) Binding site of VEGFA with quercetin. (C) Binding site of IL6 with quercetin. (D) Binding site of TP53 with quercetin. (E) Binding site of AKT1 with (R)-canadine. (F) Binding site of VEGFA with (R)-canadine. (G) Binding site of IL6 with (R)-canadine. (H) Binding site of TP53 with (R)-canadine.
Table 3 Molecular docking scores of two components of Rhizoma Coptidis and target protein (kcal/mol).
| Receptor | IL6 | VEGFA | AKT1 | TP53 |
|---|---|---|---|---|
| Ligand | ||||
| Quercetin | −4.7 | −4.4 | −5.6 | −7.8 |
| (R)-Canadine | −4.7 | −4.6 | −5.3 | −7.3 |
Table 4 RMSD of two components of Rhizoma Coptidis and target protein.
| Receptor | IL6 | VEGFA | AKT1 | TP53 |
|---|---|---|---|---|
| Ligand | ||||
| Quercetin | 1.193 | 1.558 | 1.496 | 1.205 |
| (R)-Canadine | 1.217 | 1.665 | 1.673 | 3.005 |
Discussion
As a neurological disorder, epilepsy is characterized by unpredictable and repeated occurrence of seizures.24 The global prevalence of epilepsy is about 7 cases per 1000 people.25 Behavioral and cognitive disorders are related to these seizures, which significantly impact patients’ quality of life. Patients frequently show impaired emotional learning and spatial memory when affected by epileptic seizures. Different comorbidities, such as sleep deprivation, anxiety, depression, and disorders, may have a bearing on these changes.26 Relevant relationship between epilepsy and systemic inflammation and consistent links between seizures, inflammatory cytokines (IL-6 and TNF-α), and iron regulation and metabolism, as acute phase and antioxidant markers, have been reported.27
Pharmacotherapy, primarily involving the use of anti-seizure drugs (ASDs), is an essential part of controlling seizures. However, nearly 30% of patients develop drug--resistant epilepsy, clinically defined as the persistence of seizure, following trials of two ASDs.28 The main research task is to develop new and effective drugs for epilepsy. Rhizoma Coptidis has good synergetic effect on the treatment of epilepsy.13 However, the mechanism of Rhizoma Coptidis in the treatment of epilepsy is unclear, thus its development and utilization are limited. This study used network pharmacology method to explore the anti-epilepsy mechanism of Rhizoma Coptidis, aiming to lay a foundation for the further application of Rhizoma Coptidis.
PPI results showed interactions among 68 targets, indicating that the targets were interrelated and played a role in the treatment of epilepsy through multi-channel and multi-faceted coordination. The top 10 target genes were MAPK1, EGFR, FOS, CXCL8, TNF, CASP3, VEGFA, IL6, TP53, and AKT1, which were of great significance in epilepsy treatment using Rhizoma Coptidis.
The vascular endothelial growth factor (VEGF) stimulates angiogenesis and regulates vascular permeability. It plays a certain role in neuronal regeneration and cell survival. Studies have focused on the effects of the VEGF family on the electrophysiological activities of neurons. VEGF regulates the activation of NMDA receptors in neurons through Src family protein kinase signaling pathway, mainly upregulating NR2B phosphorylation.29 In recent years, basic research has focused on the molecular mechanism and regulation of epilepsy cell apoptosis. The phosphatidylinositol 3-kinase (PI3K)/Akt signal transduction pathway is involved in cell proliferation, differentiation, and apoptosis, which is related to many physiological functions of the human body. Akt is a protective protein in the central part of this signal transduction pathway. The signal transduction pathway mediated by Akt is related to cell survival and growth.30,31 In the state of persistent epilepsy, the expression of AKT1 in rat hippocampus shows a dynamic process of increasing, decreasing and rising again, indicating that AKT1 may promote cell survival, neuron protection and antagonize apoptosis. TP53-induced glycolysis and apoptosis regulator (TIGAR), activates the pentose phosphate pathway (PPP).32 TIGAR protects neurons against oxidative stress and apoptosis in various disorders. For example, Zhou et al. showed that TIGAR is involved in cerebral ischemia by lowering ROS levels and apoptosis.33 Chen et al. showed that TP53-induced TIGAR is a promising therapeutic target for epilepsy because of its anti-epileptic, antioxidant, and anti-apoptotic effects.34 The role and the involvement of IL-6 in epilepsy have not been firmly established, but some studies implicate IL-6 signaling in epilepsy.35 Increased plasma levels of IL-6 have been reported in epilepsy.36,37 Following status epilepticus in rats,38 kindling of basolateral amygdala increased IL-6 mRNA in the rat hippocampus,39 IL-6 knockout mice showed decreased incidence of seizures induced by Theiler’s murine encephalomyelitis virus.40
As a sub-class of favonoids, quercetin is abundantly found in various vegetables and fruits.41 It shows inhibition of chronic inflammation, atherosclerosis and cancer in human.42 For example, quercetin administration can decrease histological signs of acute inflammation in animals in a dose-dependent manner by inhibiting the release of chemokine as the lipid peroxidation end-product malondialdehyde and increasing antioxidant enzyme activity.43 Quercetin also exhibits a neuro-protective function in several central nervous system disorders.44,45 Recent studies have shown that quercetin inhibited KA-induced epilepsy by microglia cell inactivation and the production of NF-κB, TNF-α, and IL-1β.
(R)-Canadine has a variety of biological activities.46 It has proved to be a new generation of dopamine (DA) antagonist in biochemistry and neuropharmacology. In addition, (R)-canadine can also increase the spontaneous discharge of DA neurons. These results not only support the conclusion that (R)-canadine is Da antagonist but also prove that one of its blocking sites is Da auto receptor (D2 receptor). Wu et al.47 used nystatin perforated patch clamp method and voltage clamp whole cell recording method to study the effect of (R)-canadine on single pyramidal neurons in hippocampal CA1 area induced by Da. The holding potential (VH) was at −20 mV. Da-induced effects including transient outward current, slow inward current, and their combination. The outward current was −83.5 ± 8.0 mV, which was close to the K+ equilibrium potential. It is sensitive to tetraethylamine (TEA), a K+ channel blocker, suggesting that K+ participates in the outward current. (R)-Canadine reversibly inhibited the three effects induced by Da.
(R)-canadine can inhibit the increase of Da-induced outward current dose–response curve, suggesting that (R)-canadine is a noncompetitive inhibition. These results confirm the hypothesis that (R)-canadine can be used as a new dopamine antagonist. In addition, (R)-canadine can inhibit Da-induced K+ current in cells, indicating that (R)-canadine may have other pharmacological effects on the central nervous system. Wu et al.48 continued to use whole cell patch clamp technique and found that (R)-canadine not only inhibited Da-induced outward K+ current but also inhibited Ach-, caffeine-, or strychnine-induced outward K+ current. Wu et al.49 found that (R)-canadine can block the K-ATP channel of DA neurons in acutely isolated rat compact part of substantia nigra (SNC). In the current clamp mode, functional K-ATP channels were opened by continuous perfusion of rotenone, an inhibitor of mitochondrial respiratory chain complex I, with perforated patch clamp technique. (R)-canadine can reversibly block the opening of K-ATP channel in a dose--dependent manner. Compared with (R)-canadine analogues, (R)-canadine showed stronger blocking effect. In addition, (R)-canadine exposure alone increased the occurrence of action potential. (R)-canadine plays an important role in dopamine D2 receptor antagonist (sulpiride). In the presence of (R)-canadine, the membrane hyperpolarization induced by rotenone was restored, which indicated that (R)-canadine could excite DA neurons in SNC by blocking K-ATP channel. It was found that the expression of K-ATP in SNC DA neurons was significantly increased. The blocking of THB channel is a new mechanism of (R)-canadine, which may contribute to its neuro-protective effect in the treatment of Parkinson’s disease. Xu Changqing et al.50 first used 3.3 μmol/L β-Amyloid peptide-induced apoptosis of cultured hippocampal neurons in vitro to establish the model. The appearance of apoptotic nucleus or apoptotic body, the shrinkage and budding of neuronal cell body, and the state of apoptotic cells were observed with light, scanning electron, and transmission electron microscopes, and the number of apoptotic cells was counted through flow cytometry. Used 3 μmol/L Aβ, the apoptosis model of hippocampal neurons was established successfully. The effect of (R)-canadine on Aβ apoptosis of cultured hippocampal neurons in vitro and the effect of free Ca2+ in neurons were studied. The results showed that 1, 10, and 100 μ (R)-canadine could significantly reduce Aβ with the increase of drug concentration, and the effect of reducing the number of apoptotic cells was more obvious. Compared with the model group, (R)-canadine could significantly increase the survival number of neurons in a dose-dependent manner. Li Shuyu’s results showed that quercetin of 50mg/kg could increase the expression of XIAP in hippocampal neurons after status epilepticus in rats,51 and quercetin has the most corresponding therapeutic targets, so it may be the most important compound of Rhizoma Coptidis in the treatment of epilepsy.
TCM has the characteristics of multi-component and multi-target. The effect of TCM refers to the comprehensive result that the effective molecular combination of TCM widely regulates the biological process of the body, and some active compounds in TCM act on the body through different pathways, so as to play a therapeutic effect. Therefore, through network pharmacology methods, we can get the “drug-component-target-pathway” network based on the theory of TCM, which can help comprehensively understand the mechanism of TCM in the treatment of diseases and provide data for clinical treatment. In this study, network pharmacology methods were used, and according to the results, the possible mechanism of Rhizoma Coptidis for epilepsy was proposed. Therefore, further clinical verification is needed, so as to more systematically and scientifically elaborate the mechanism of Rhizoma Coptidis in the treatment of epilepsy.52
Conclusion
Our research may inspire and guide further work, establish the molecular target of Rhizoma Coptidis in the treatment of epilepsy, and apply the network pharmacology method to develop new drug. This study showed that TNF signaling pathway and AKT1, TP53, IL6, and other molecules are of great significance in the treatment of epilepsy, which must be thoroughly tested in vivo and in vitro.
Acknowledgments
Funding: Hainan Province Key R&D Program (Grant No. ZDYF2020165) and Natural Science Foundation of Hainan Province(820MS134).
Funding
No.
Conflict of Interest
The authors declare no conflict of interest.
Data Availability Statement
All of the data were based on public data and no additional data were available.
REFERENCES
1. Wennberg R, Hiploylee C, Tai P, et al. Is concussion a risk factor for epilepsy? Can J Neurol Sci. 2018;45:275–282. 10.1017/cjn.2017.300
2. Uslu FI, Gokcal E, Gelisin O. Epilepsy-related knowledge and attitudes toward people with epilepsy among hospital staff in Van City, Turkey. Epilepsy Behav. 2020;103. 10.1016/j.yebeh.2019.04.014
3. Casotto V, Ranzato F, Girardi P, et al. Increasing epilepsy-related mortality: A multiple causes of death study in Northern Italy. Seizure-Eur J Epilep. 2022;94:1–6. 10.1016/j.seizure.2021.11.003
4. Ru YD,Li JB, Chen HY, et al. Epilepsy Detection Based on Variational Mode Decomposition and Improved Sample Entropy. Comput Intel Neurosc. 2022. 10.1155/2022/6180441
5. Lu L, Jia Y, Kirunda JB, et al. Effects of noise and synaptic weight on propagation of subthreshold excitatory postsynaptic current signal in a feed-forward neural network. Nonlin Dynam. 2019;95:1673–1686. 10.1007/s11071-018-4652-9
6. Triplet EM, Nickels K , Wong-Kisiel L, et al. A tale of two cohorts: Differing outcomes in infantile-onset focal epilepsy. Epilepsia. 2022. 10.1111/epi.17181
7. Xia L, Liu L, Wang Q, Relationship Between the Pyroptosis Pathway and Epilepsy: A Bioinformatic Analysis. Front Neurol. 2022; 10.3389/fneur.2021.782739
8. Patel DC, Tewari BP, Chaunsali L, et al. Neuron-glia interactions in the pathophysiology of epilepsy. Nat Rev Neurosci. 2019;20:282–297. 10.1038/s41583-019-0126-4
9. Deng P, Liang H, Xie KM, et al. Study on the molecular mechanism of Guizhi Jia Shaoyao decoction for the treatment of knee osteoarthritis by utilizing network pharmacology and molecular docking technology. Allergol Immunopath. 2021;49(6):16–30. 10.15586/aei.v49i6.484
10. Dong YK, Tao B, Xue X; et al. Molecular mechanism of Epicedium treatment for depression based on network pharmacology and molecular dockingtechnology. BMC Complement Med. 2021;21(1):1–13. 10.1186/s12906-021-03389-w
11. Roh KB, Ryu D, Cho E, et al. Coptis chinensis Franch Directly Inhibits Proteolytic Activation of Kallikrein 5 and Cathelicidin Associated with Rosacea in Epidermal Keratinocytes. Molecules. 2020;25(23):1–17. 10.3390/molecules25235556
12. Park SM, Min B-G, Jung JY, et al. Combination of Pelargonium sidoides and Coptis chinensis root inhibits nuclear factor kappa B-mediated inflammatory response in vitro and in vivo. BMC Complement Alternat Med. 2018;18. 10.1186/s12906-018-2088-x
13. Wang L, Zhuang B, Wang S, et al. Study of coptidis rhizoma on the synergism to epilepsy and the regulation to brain delivery of antiepileptic drugs mediated by P-glycoprotein. Chin J Mod Appl Pharm. 2015;32:660–663.
14. Ma C, Xu T, Sun X, et al. Network pharmacology and bioinformatics approach reveals the therapeutic mechanism of action of baicalein in hepatocellular carcinoma. Evid Based Complement Alternat Med. 2019;2019. 10.1155/2019/7518374
15. Ru J, Li P, Wang J, et al. TCMSP: a database of systems pharmacology for drug discovery from herbal medicines. J Cheminform. 2014;6. 10.1186/1758-2946-6-13
16. Xu X, Zhang W, Huang C, et al. A novel chemometric method for the prediction of human oral bioavailability. Int J Molec Sci. 2012;13:6964–6982. 10.3390/ijms13066964
17. Walters WP, Murcko MA. Prediction of “drug-likeness.” Adv Drug Deliv Rev. 2002;54:255–271. 10.1016/S0169-409X(02)00003-0
18. Feng W, Ao H, Yue S, et al. Systems pharmacology reveals the unique mechanism features of Shenzhu Capsule for treatment of ulcerative colitis in comparison with synthetic drugs. Sci Rep. 2018;8. 10.1038/s41598-018-34509-1
19. von Mering C, Jensen LJ, Snel B, et al. STRING: known and predicted protein-protein associations, integrated and transferred across organisms. Nucl Acids Res. 2005;33:D433–D437. 10.1093/nar/gki005
20. Huang DW, Sherman BT, Lempicki RA. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat Protoc. 2009;4:44–57. 10.1038/nprot.2008.211
21. Chen L, Zhang Y-H, Wang S, et al. Prediction and analysis of essential genes using the enrichments of gene ontology and KEGG pathways. PLoS One. 2017;12. 10.1371/journal.pone.0184129
22. Carpten JD, Faber AL, Horn C, et al. A transforming mutation in the pleckstrin homology domain of AKT1 in cancer. Nature. 2007;448:439–U1. 10.1038/nature05933
23. Somers W, Stahl M, Seehra JS. 1.9 A crystal structure of interleukin 6: implications for a novel mode of receptor dimerization and signaling. EMBO J. 1997;16:989–997. 10.1093/emboj/16.5.989
24. Ek RO, Sirinyildiz F. Effects of melatonin at different doses on experimental epilepsy model induced by pentylenetetrazole. Meandros Med Dent J. 2021;22:177–186. 10.4274/meandros.galenos.2021.49358
25. Robinson SJ. Childhood epilepsy and autism spectrum disorders: psychiatric problems, phenotypic expression, and anticonvulsants. Neuropsychol Rev. 2012;22:271–279. 10.1007/s11065-012-9212-3
26. Mosinska P, Socala K, Nieoczym D, et al. Anticonvulsant activity of melatonin, but not melatonin receptor agonists Neu-P11 and Neu-P67, in mice. Behav Brain Res. 2016;307:199–207. 10.1016/j.bbr.2016.03.036
27. Tombini M, Squitti R, Cacciapaglia F, et al. Inflammation and iron metabolism in adult patients with epilepsy: does a link exist? Epilepsy Res. 2013;107:244–252. 10.1016/j.eplepsyres.2013.09.010
28. Achar A, Ghosh C. Multiple hurdle mechanism and blood-brain barrier in epilepsy: glucocorticoid receptor-heat shock proteins on drug regulation. Neur Regener Res. 2021;16:2427–2428. 10.4103/1673-5374.313046
29. Howell KR, Hoda MN, Pillai A. VEGF activates NR2B phosphorylation through Dab1 pathway. Neurosci Lett. 2013;552:30–34. 10.1016/j.neulet.2013.07.032
30. Terasaki K, Gen Y, Iwai N, et al. SOX2 enhances cell survival and induces resistance to apoptosis under serum starvation conditions through the AKT/GSK-3 beta signaling pathway in esophageal squamous cell carcinoma. Oncol Lett. 2021;21(4):1–7. 10.3892/ol.2021.12530
31. Camaya I, Donnelly S, O’Brien B. Targeting the PI3K/Akt signaling pathway in pancreaticβ-cells to enhance their survival and function: An emerging therapeutic strategy for type 1 diabetes. 2022. J Diabetes. 10.1111/1753-0407.13252
32. Chen CY, Mei Q, Wang LL, et al. TIGAR suppresses seizures induced by kainic acid through inhibiting oxidative stress and neuronal apoptosis. Biochem Bioph Res Co. 2019.10.1016/j.bbrc.2019.05.156
33. Zhou J-H, Zhang T-T, Song D-D, et al. TIGAR contributes to ischemic tolerance induced by cerebral preconditioning through scavenging of reactive oxygen species and inhibition of apoptosis. Sci Rep. 2016;6. 10.1038/srep27096
34. Chen C, Mei Q, Wang L, et al. TIGAR suppresses seizures induced by kainic acid through inhibiting oxidative stress and neuronal apoptosis. Biochem Biophys Res Commun. 2019;515:436–441. 10.1016/j.bbrc.2019.05.156
35. Washington J, III, Kumar U, Medel-Matus J-S, et al. Cytokine-dependent bidirectional connection between impaired social behavior and susceptibility to seizures associated with maternal immune activation in mice. Epilepsy Behav. 2015;50:40–45. 10.1016/j.yebeh.2015.05.040
36. Chmielewska N, Maciejak P, Osuch B, et al. Pro-inflammatory cytokines, but not brain-and extracellular matrix-derived proteins, are increased in the plasma following electrically induced kindling of seizures. Pharmacol Rep. 2021;73(2):506–515. 10.1007/s43440-020-00208-w
37. Ocak O, Zorlu, Y, Kirkali G, et al. The role of plasma cytokine levels in the differential diagnosis of epileptic and psychogenic non-epileptic seizures. Ann Clin Anal Med. 2021; 12(S2):223–227.
38. Putra M, Gage M, Sharma S, et al. Diapocynin, an NADPH oxidase inhibitor, counteracts diisopropylfluorophosphate--induced long-term neurotoxicity in the rat model. Ann Ny Acad Sci. 2020;1479(1):75–93.
39. Ye F, Chen X-Q, Bao G-S, et al. Effect of topiramate on interleukin 6 expression in the hippocampus of amygdala-kindled epileptic rats. Exp Therap Med. 2014;7:223–227. 10.3892/etm.2013.1396
40. Cusick MF, Libbey JE, Doty DJ, et al. The role of peripheral interleukin-6 in the development of acute seizures following virus encephalitis. J Neurovirol. 2017;23(5):696–703. 10.1007/s13365-017-0554-8
41. Ozyilmaz E, Arpaci PU, Sayin S, et al. Inclusion complexes of water-soluble calix[n]arenes with quercetin: preparation, characterization, water solubility, and antioxidant features. Monatsh Chem. 2022;153(2):201–209. 10.1007/s00706-021-02885-6
42. Chen C, Zhong QX, Chen ZX. Improved aqueous solubility, bioaccessibility and cellular uptake of quercetin following pH-driven encapsulation in whey protein isolate. Int J Food Sci Tech. 2022. 10.1111/ijfs.15579
43. Dong Y-S, Wang J-L, Feng D-Y, et al. Protective effect of quercetin against oxidative stress and brain edema in an experimental rat model of subarachnoid hemorrhage. Int J Med Sci. 2014;11:282–290. 10.7150/ijms.7634
44. Wu Y, Wei HP, Li P, et al. Quercetin Administration Following Hypoxia-Induced Neonatal Brain Damage Attenuates Later-Life Seizure Susceptibility and Anxiety-Related Behavior: Modulating Inflammatory Response. Front Pediatr. 2022.10.3389/fped.2022.791815
45. Enogieru AB, Haylett W, Hiss DC, et al. Rutin as a Potent Antioxidant: Implications for Neurodegenerative Disorders. Oxid Med Cell Longev. 2018. 10.1155/2018/6241017
46. Lu Z, Sun W, Duan X, et al. Chemical constituents from Corydalis yanhusuo. Zhongguo Zhong Yao Za Zhi. 2012;37:235–237.
47. Wu J, Jin GZ. Tetrahydroberberine inhibits acetylcholine--induced K+ current in acutely dissociated rat hippocampal CA1 pyramidal neurons. Neurosci Lett. 1997;222:115–118. 10.1016/S0304-3940(97)13356-0
48. Wu J, Jin GZ. Tetrahydroberberine blocks membrane K+ chan-nels underlying its inhibition of intracellular message-me-diated outward currents in acutely dissociated CA1 neurons from rat hippocampus. Brain Res. 1997;775:214–218. 10.1016/S0006-8993(97)00960-8
49. Wu C, Yang K, Liu Q, et al. Tetrahydroberberine blocks ATP-sensitive potassium channels in dopamine neurons acutely-dissociated from rat substantia nigra pars com-pacta. Neuropharmacology. 2010;59:567–572. 10.1016/j.neuropharm.2010.08.018
50. Xu CQ. Research on the mechanism of neuronal apoptosis in Alzheimer’s disease and the effects of tetrohydroberberine on the apoptosis. Sheng Li Ke Xue Jin Zhan [Progr Physiol]. 1999;30:224–226.
51. Shuyu LI, Bo X, Fangfang BI, et al. The expression of XIAP mRNA and protein in the rat hippocampus and quercetin treatment following status epilepticus. Chin J Nerv Ment Dis. 2008;34:542–545.
52. Wu D, Zheng Z, Fan S, et al. Ameliorating effect of quercetin on epilepsy by inhibition of inflammation in glial cells. Exp Therap Med. 2020;20:854–859. 10.3892/etm.2020.8742