
Download
ORIGINAL ARTICLE
Knockdown of GBP5 alleviates renal damage caused by psoriasis by regulating NF-κB/STAT3 pathway
Fang Ren†, Jin Li†, Xingyan Ni, Hongshan Yuan, Wenliang Yan*
Department of Dermatology, Jinling Hospital, Affiliated Hospital of Medical School, Nanjing University, Nanjing, Jiangsu, China
Abstract
Background: Kidney impairment resulting from psoriasis constitutes a serious complication, affecting the overall well-being of patients and necessitating a thorough comprehension for efficient management. Guanylate-binding protein 5 (GBP5) is known to play a role in inflammatory responses, but its function in psoriasis remains unclear and warrants investigation in.
Objective: To pinpoint GBP5 as innovative therapeutic target and decipher the underlying mechanisms in kidney impairment resulting from psoriasis.
Methods: Skin samples from psoriatic patients were used to detect GBP5 expression through Immunoblot and qPCR. Hacat cells were treated with TNF-α to construct the psoriasis skin cell model. Edu and CCK-8 assays were performed to confirm the effects on cell viability, ELISA was conducted to confirm the effects on inflammation. H&E staining and PASI scocing were conducted to confirm the effects on renal damage. Immunoblot confirmed the mechanism.
Results: GBP5 was highly expressed in psoriasis skin tissues. Ablation of GBP5 reduced tumor necrosis factor alpha (TNF-α)-stimulated growth as well as inflammation in human immortalized keratinocyte (HaCaT) cell. In the imiquimod (IMQ)-stimulated mouse model, GBP5 knockdown alleviated psoriasis symptoms and reduced renal damage. Mechanically, GBP5 depletion suppressed the activation of nuclear factor kappa-light-chain-enhancer of activated B cells–signal transducer and activator of transcription 3 (NF-κB/STAT3) axis.
Conclusion: Inhibiting GBP5 can mitigate the renal injury caused by psoriasis through NF-κB/STAT3 axix.
Key words: GBP5, Inflammation, NF-κB/STAT3 pathway, psoriasis, renal damage
*Corresponding author: Wenliang Yan, Department of Dermatology, Jinling Hospital, Affiliated Hospital of Medical School, Nanjing University, No. 305 Zhongshan East Road, Nanjing, Jiangsu 210002, China. Email address: [email protected]
Received 8 August 2024; Accepted 5 October 2024; Available online 1 November 2024
†These authors contributed equally to the work.
Copyright: Ren F, et al.
License: This open access article is licensed under Creative Commons Attribution 4.0 International (CC BY 4.0). http://creativecommons.org/licenses/by/4.0/
Introduction
Psoriasis is a type of chronic inflammatory disease featured by the autoimmunity and a strong genetic predisposition, affecting 2% of global population.1 The chronic relapsing nature of this illness poses a lifelong treatment challenge, leading to a substantial financial burden on patients.2 Psoriasis is typified by abnormal keratinization, epidermal hyperproliferation, and inflammation.3
Glomerular podocytes play a crucial role in psoriasis pathogenesis.4 These cells are responsible for creating filtration barrier in the glomeruli, which is responsible for filtering and eliminating waste products and surplus substances from the bloodstream.5 Patients with psoriasis often exhibit renal impairment, but the precise mechanism remains unclear.
Research indicates that psoriasis may trigger damage to podocytes and induce apoptosis by activating inflammation, which could be associated with increased levels of inflammation and atypical autoimmune reactions.6 The pathogenesis of psoriasis is not fully understood, but it is widely believed that inflammatory responses and abnormal activation of immune cells play key roles in its development and progression. Overproduction of inflammatory cytokines triggers a cascade of intracellular signaling pathways, which in turn activate transcription factors, resulting in an upsurge of cytokine secretion from immune cells.7 This exacerbates epidermal symptoms and ultimately worsens psoriasis.8
Previous research indicates that the nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) and signal transducer and activator of transcription (STAT) are involved in the gene expression of cytokines during inflammation.9,10 Tumor necrosis factor alpha (TNF-α) was employed to induce the development of a psoriasis cell model, triggering the NF-κB pathway and enhancing the levels of phosphorylated (p)-NF-κB) expression.11 Activated NF-κB translocates to the nucleus and affects the downstream processes.12
Guanylate-binding protein 5 (GBP5) is a member of interferon gamma (IFN-γ)-induced guanosine triphosphate (GTPase) family, involved in cellular inflammatory responses, including inflammasome activation and cytokine release.13 Research indicates a rise in GBP5 expression within the renal cortex tissue of mice with lupus nephritis, and hindering this can improve renal damage by reducing pro-inflammatory factors.14 GBP5 expression is significantly upregulated in rosacea-like skin inflammation, where it exacerbates inflammation by skewing macrophage polarization toward the M1 phenotype via NF-κB axis.15 Further investigation is required to understand the impact of GBP5 on NF-κB pathway in various diseases. Moreover, GBP5 expression tends to increase, enhancing the proliferation and migration of gastric cancer cells.16 However, the role and mechanism of GBP5 in psoriasis and psoriasis-induced renal damage (PRRD) remain unclear.
This study aims to investigate the role of GBP5 in psoriasis and its associated renal damage. We first indicated that GBP5 could affect psoriasis and renal impairment via NF-κB/STAT3 axis, potentially leading to the development of novel treatment strategies.
Materials and Methods
Human samples
Skin tissue specimens were obtained from a cohort of 20 psoriatic patients. Specimens from psoriatic patients were extracted from the affected skin regions whereas samples from healthy controls were procured from the unaffected skin surrounding the areas affected by psoriasis. The study was approved by the Ethics Committee of Jinling Hospital, Affiliated Hospital of Medical School, Nanjing University, China (2023DZKY-035-0l).
Cell culture and treatment
Human immortalized keratinocyte (HaCaT) cells were cultured in Dulbecco’s modified eagle medium (DMEM; C11995500BT; Beyotime, Shanghai, China) supplemented with 10% fetal bovine serum (FBS; C0221; Beyotime). Cells were treated with TNF-α (10 ng/mL; P5055; Beyotime) for 12 h to induce inflammatory response. To conduct knockdown experiments, HaCaT cells were transfected with either short hairpin RNA (shRNA) specifically targeting sh-GBP5 or a short hairpin non-targeting control (sh-NC) using lipofectamine 3000 (L3000015; Invitrogen, Carlsbad, CA, USA).
Animal model
Male BALB/c mice (8–11 weeks old) were used to develop an imiquimod (IMQ)-induced psoriasis-like model. Imiquimod (62.5 mg; HY-15562; MedChem Express) was applied topically to the shaved back of mice consecutively for 7 days. Mice were divided into the following four groups: control, IMQ, IMQ+sh-NC, and IMQ+sh-GBP5. GBP5 knockdown was achieved by intradermal injection of sh-GBP5 lentiviral particles (GenePharma, Shanghai, China). Mice were intravenously (i.v.) injected with a mixture of three shRNA (2×1011 vg/mice) viruses and scrambled shRNA (2×1011 vg/mice, i.v.) via tail vein injection using a 30-gauge needle. Further experiments were conducted using dorsal skin and renal tissues. Supplementary Figure S1 illustrated the schedule of injection administration to animal in terms of time and dosage.
Immunoblot analysis
Equal amounts of protein were separated by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE), transferred to polyvinylidene fluoride (PVDF) membranes (IPVH00010; Millipore, MA, USA), and then blocked with 5% non-fat milk. The membranes were incubated overnight with primary antibodies at 4°C, including anti-GBP5 (1:500; ab109006; Abcam, Cambridge, UK), anti-β-actin (1:500; ab8227; Abcam), anti-p-p65 (1:1000; ab86299; Abcam), anti-p65 (1:500; ab16502; Abcam), anti-p-IκBα (1:500; ab12135; Abcam), anti-IκBα (1:1000; ab32518; Abcam), anti-p-STAT3 (1:300; ab76315; Abcam), and anti-STAT3 (1:300; ab68153; Abcam). After washing, membranes were incubated with horseradish peroxidase (HRP) secondary antibodies (1:2000; ab6721; Abcam). Bands were visualized using an enhanced chemiluminescence (ECL) detection kit.
Quantitative Polymerase Chain Reaction (qPCR) Analysis
Total RNA was extracted using TRIzol reagent (15596026; Invitrogen) and reverse transcribed into cDNA using a reverse transcription kit (D7168M; Beyotime). qPCR was performed using SYBR Green Master Mix (D7260M; Beyotime). The mRNA levels of target genes were normalized to glyceraldehyde 3-phosphate dehydrogenase (GAPDH) and calculated using the 2-ΔΔCt method. Primers used for qPCR analysis were as follows: GBP5: forward: CAGTGCATCAGTGGCATCAT, reverse: GAGGTCTCCAGGTTGATGGA; IL-1β: forward: ATGATGGCTTATTACAGTGGCAA, reverse: GTCGGAGATTCGTAGCTGGA; ICAM-1: forward: ATGCCCAGACATCTGTGTCC, reverse: TGGAGTTCACCCCTTTTTCC; IL-6: forward: ACTCACCTCTTCAGAACGAATTG, reverse: CCATCTTTGGAAGGTTCAGGTTG; TNF-α: forward: CCTCTCTCTAATCAGCCCTCTG, reverse: GAGGACCTGGGAGTAGATGAG; and GAPDH: forward: GGAGCGAGATCCCTCCAAAAT, reverse: GGCTGTTGTCATACTTCTCATGG.
Cell Counting Kit-8 (CCK-8) assay
Cell proliferation was assessed with CCK-8 (C0038; Beyotime) assay, and the optical density at 450 nm was quantified using a microplate reader (Bio-Rad, Hercules, CA, USA).
EdU staining
Cell proliferation was also evaluated using the 5-ethynyl-2’-deoxyuridine (EdU) cell proliferation kit (C10310-3; Beyotime). HaCaT cells were incubated with EdU for 2 h, fixed with 4% paraformaldehyde (PFA), and the percentage of EdU-positive cells was determined (Zeiss Axio Observer, Cambridge, UK).
Enzyme-Linked-Immunosorbent Serologic Assay (ELISA)
Levels of IL-1β, TNF-α, and IL-6 were measured using ELISA kits (PI303, PI305, and PI330; Beyotime). In brief, the samples were pipetted into wells, followed by the introduction of primary antibodies conjugated with biotin. Avidin conjugated with HRP was added to the wells. Subsequently, an enzyme substrate was introduced to initiate color development. The optical density of each well was assessed using a microplate reader.
Histological analysis
Sections were stained with hematoxylin and eosin (H&E; C0105; Beyotime) for histopathological examination. Images were captured using a light microscope (Zeiss, Axio Observer), and pathological changes were evaluated by a blinded pathologist.
Psoriasis Area and Severity Index (PASI) scoring
The severity of psoriasis in IMQ-treated mice was evaluated using PASI. PASI scores were recorded on days 0, 2, 4, and 7.
Serum biochemistry
Serum creatinine (Scr) and blood Urea Nitrogen (BUN) levels were measured using commercial assay kits (C013-2-1 and C011-2-1; Beyotime). Animal blood samples were collected, and serum was separated by centrifugation.
Statistical analysis
Data were presented as mean±SD. Statistical comparisons between groups were made using one-way ANOVA, followed by Tukey’s post hoc test; P < 0.05 was considered statistically significant.
Results
GBP5 is highly expressed in psoriasis skin tissues
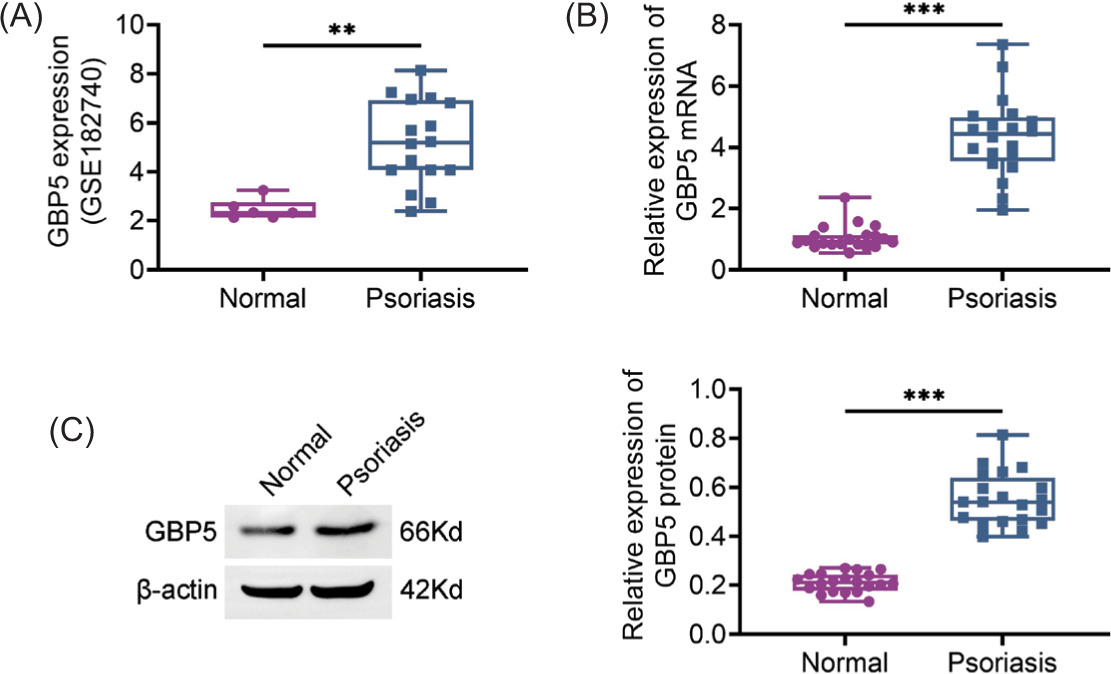
In order to investigate the role of GBP5 in psoriasis and its associated renal damage, initially we examined the expression levels of GBP5 in psoriasis skin tissues. Analysis of the GSE182740 dataset revealed a rise in the levels of GBP5 protein in the samples of patients with psoriasis (Figure 1A). Likewise, qPCR findings confirmed the increased expression of GBP5 mRNA in skin tissues affected by psoriasis (Figure 1B). Subsequent immunoblot analysis corroborated these results by illustrating heightened GBP5 protein levels in psoriasis samples in comparison to those from healthy skin tissues (Figure 1C). The robust expression of GBP5 in psoriatic skin tissues suggests its plausible role in the pathogenesis of the disease.
Figure 1 GBP5 is highly expressed in psoriasis skin tissues. (A) GBP5 expression levels in normal and psoriasis skin tissues were analyzed using the GSE182740 dataset. (B) Relative expression of GBP5 mRNA in normal and psoriasis skin tissues measured by qPCR. (C) Relative expression of GBP5 protein in normal and psoriasis skin tissues analyzed by Immunoblot analysis. Data are presented as mean ± SD. **P < 0.01, ***P < 0.001, compared to control.
Knockdown of GBP5 reduces TNF-α-stimulated growth as well as inflammation in HaCaT cells
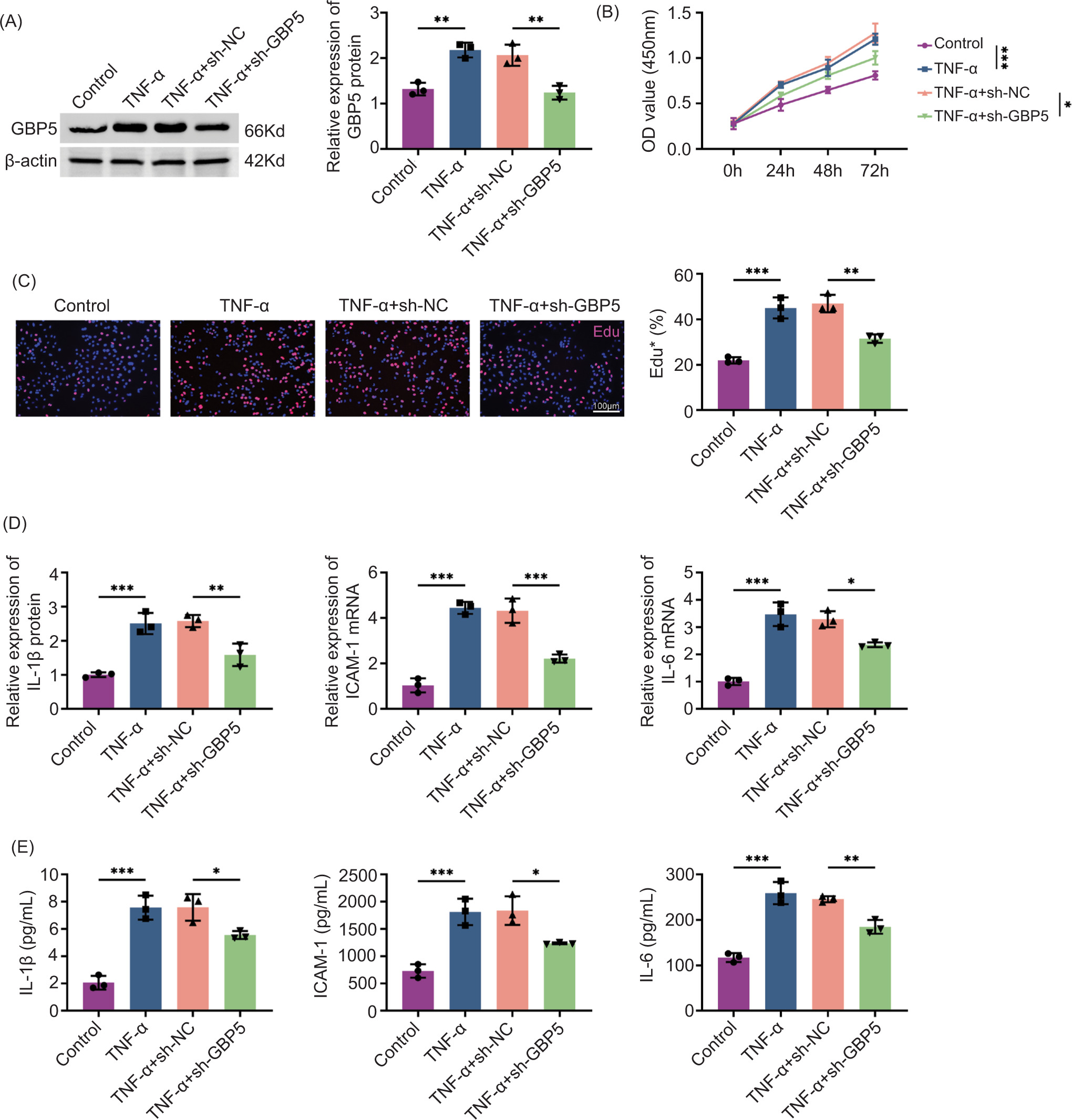
Given the elevated expression of GBP5 in psoriasis tissues, we next explored its functional role in keratinocyte proliferation and inflammation, which are key features of psoriasis. HaCaT cells were stimulated with TNF-α to induce inflammatory response, followed by GBP5 knockdown using sh-GBP5. Immunoblot analysis confirmed that GBP5 was effectively reduced in cells treated with sh-GBP5 (Figure 2A). Cell proliferation, measured by CCK-8 assay, was significantly reduced in the TNF-α+sh-GBP5 group (Figure 2B). Edu staining further confirmed that GBP5 knockdown reduced the percentage of proliferating cells in the TNF-α+sh-GBP5 group (Figure 2C). qPCR analysis revealed that the mRNA levels of inflammatory cytokines IL-1β, ICAM-1, and IL-6 were significantly decreased in the TNF-α+sh-GBP5 group (Figure 2D). In the TNF-α+sh-GBP5 group, a notable decrease was observed in the levels of IL-1β, ICAM-1, and IL-6 in cell culture supernatants, as depicted in Figure 2E. This suggests that inhibiting GBP5 alleviates both growth and inflammatory responses induced by TNF-α in HaCaT cells.
Figure 2 Knockdown of GBP5 reduces TNF-α-induced proliferation and inflammation in HaCaT cells. (A) Immunoblot analysis showing the relative expression of GBP5 protein in HaCaT cells treated with TNF-α with or without sh-GBP5 knockdown. (B) Cell proliferation measured by CCK-8 assay at different time points (0 h, 24 h, 48 h, 72 h) in control, TNF-α, TNF-α+sh-NC, and TNF-α+sh-GBP5 groups. (C) Edu staining showing the percentage of proliferating cells in control, TNF-α, TNF-α+sh-NC, and TNF-α+sh-GBP5 groups. Scale bar: 100 um. (D) Relative expression levels of IL-1β, ICAM-1, and IL-6 mRNA in HaCaT cells measured by qPCR in control, TNF-α, TNF-α+sh-NC, and TNF-α+sh-GBP5 groups. (E) Levels of IL-1β, ICAM-1, and IL-6 in cell culture supernatants measured by ELISA in control, TNF-α, TNF-α+sh-NC, and TNF-α+sh-GBP5 groups. Data are presented as mean±SD. **P < 0.01, ***P < 0.001, compared to control and TNF+sh-NC.
Knockdown of GBP5 alleviates psoriasis symptoms in IMQ-stimulated mice
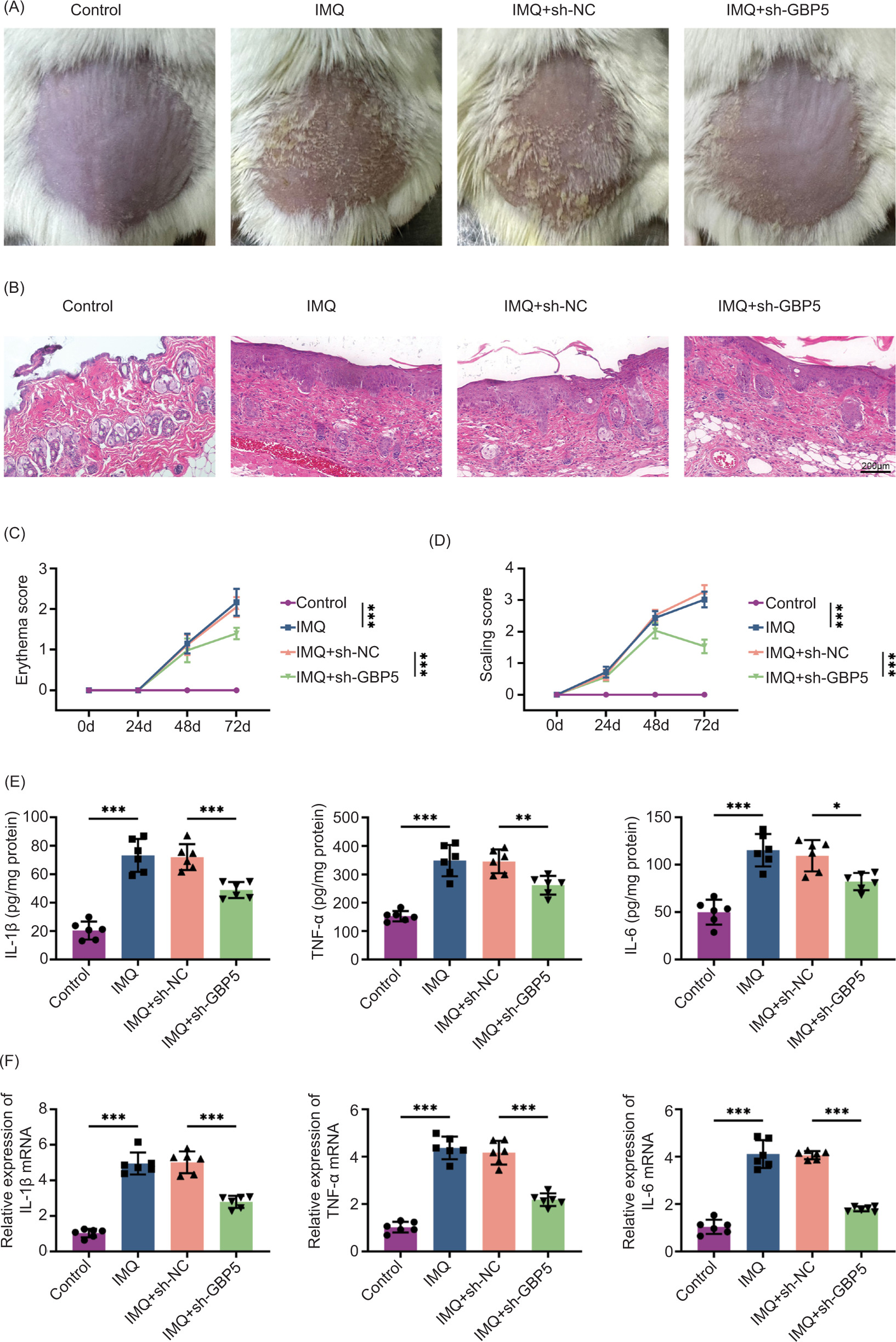
To further evaluate the in vivo effects of GBP5 knockdown, we used an IMQ-stimulated psoriasis-like mouse model. Mice administered with IMQ exhibited typical symptoms of psoriasis, such as redness and flaking, which were notably alleviated in the IMQ+sh-GBP5 treated group (Figure 3A). Examination of dorsal skin tissues through histological analysis revealed a decrease in the thickening of the epidermis and the infiltration of inflammatory cells in the IMQ+sh-GBP5 treated group (Figure 3B). Throughout the experiment, the scores for redness and flaking were significantly lower in the IMQ+sh-GBP5 treated group (Figures 3C and D). The ELISA results demonstrated a considerable reduction in the levels of inflammatory factors in the dorsal skin tissues of the IMQ+sh-GBP5 treated group (Figure 3E). Moreover, qPCR analysis validated a decrease in the mRNA expression levels of inflammatory factors in the dorsal skin tissues of the IMQ+sh-GBP5 treated group (Figure 3F). These results demonstrate that GBP5 knockdown alleviates psoriasis symptoms in IMQ-induced mice.
Figure 3 Knockdown of GBP5 alleviates psoriasis symptoms in IMQ-induced mice. (A) Photographs showing the skin condition of IMQ-induced mice treated with sh-GBP5, compared to control, IMQ, and IMQ+sh-NC groups. (B) Histological analysis of skin tissues from different groups stained with H&E. (C) Erythema scores of mice in control, IMQ, IMQ+sh-NC, and IMQ+sh-GBP5 groups over time. (D) Scaling scores of mice in control, IMQ, IMQ+sh-NC, and IMQ+sh-GBP5 groups over time. (E) Levels of IL-1β, TNF-α, and IL-6 in skin tissues measured by ELISA in control, IMQ, IMQ+sh-NC, and IMQ+sh-GBP5 groups. (F) Relative expression levels of IL-1β, TNF-α, and IL-6 mRNA in skin tissues measured by qPCR in control, IMQ, IMQ+sh-NC, and IMQ+sh-GBP5 groups. Data are presented as mean±SD. *P < 0.05, **P< 0.01, ***P< 0.001, compared to control and TNF+sh-NC.
Knockdown of GBP5 reduces renal damage in IMQ-induced mice
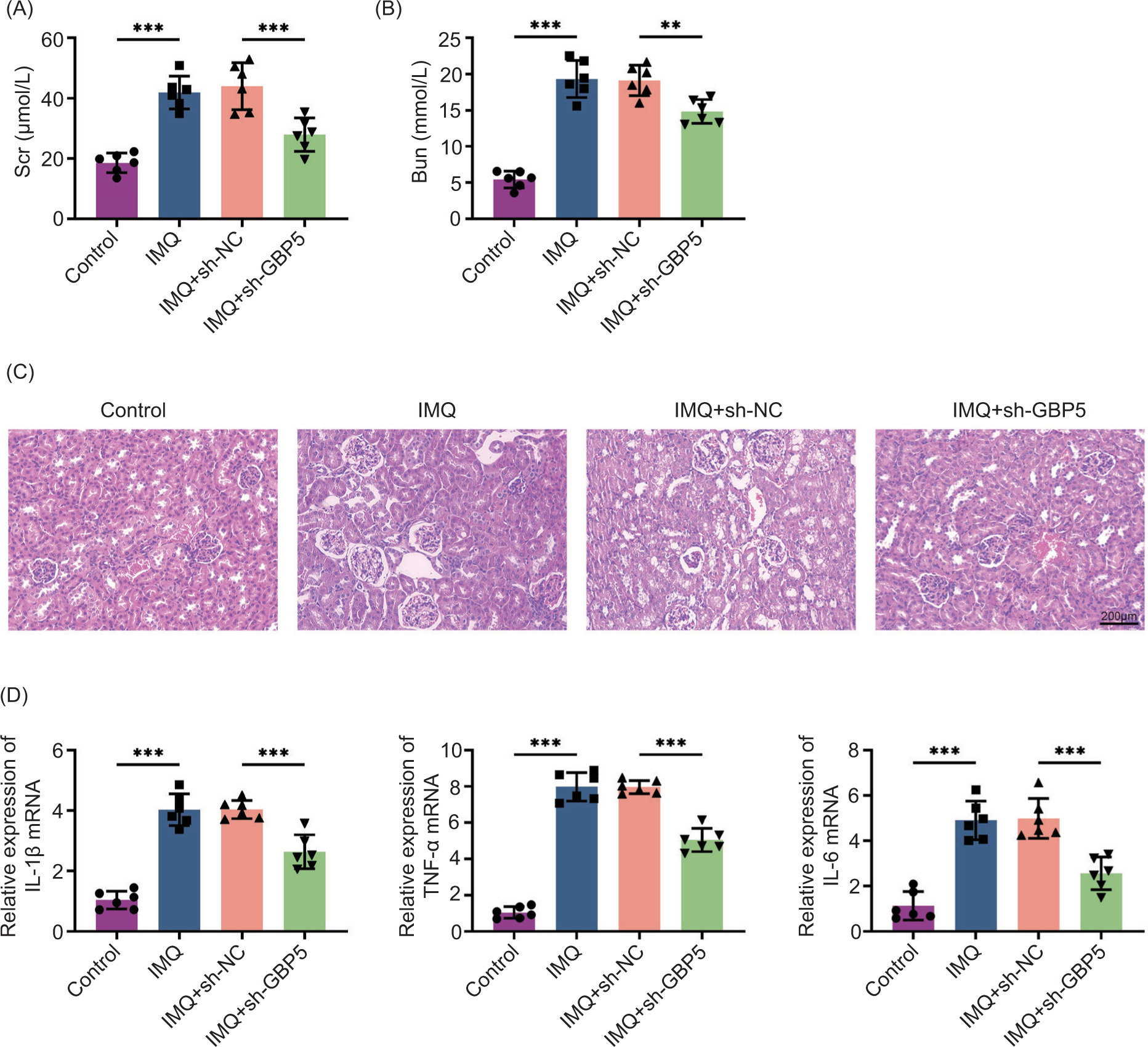
Considering that psoriasis can lead to systemic complications, including renal damage, we assessed the impact of GBP5 knockdown on renal damage in the IMQ-induced mouse model. Scr and Bun levels, which are indicators of renal function, were measured. Both Scr and Bun levels were significantly elevated in IMQ-treated mice but were reduced in the IMQ+sh-GBP5 group (Figures 4A,B). Histological examination of kidney tissues indicated a reduction in both glomerular and tubular damage in the group treated with IMQ and sh-GBP5 (Figure 4C). Of note, the analysis through qPCR analysis revealed a notable decrease in the mRNA expression levels of inflammatory factors within kidney tissues in the IMQ+sh-GBP5 group (Figure 4D). These results collectively suggest that the knockdown of GBP5 contributes to the mitigation of renal damage in IMQ-induced mice.
Figure 4 Knockdown of GBP5 reduces renal damage in IMQ-induced mice. (A) Serum creatinine (Scr) levels in control, IMQ, IMQ+sh-NC, and IMQ+sh-GBP5 groups. (B) Blood urea nitrogen (Bun) levels in control, IMQ, IMQ+sh-NC, and IMQ+sh-GBP5 groups. (C) Histological analysis of kidney tissues from different groups stained with H&E. Scale bar: 200 um. (D) Relative expression levels of IL-1β, TNF-α, and IL-6 mRNA in kidney tissues measured by qPCR in control, IMQ, IMQ+sh-NC, and IMQ+sh-GBP5 groups. Data are presented as mean±SD. **P < 0.01, ***P < 0.001, compared to control.
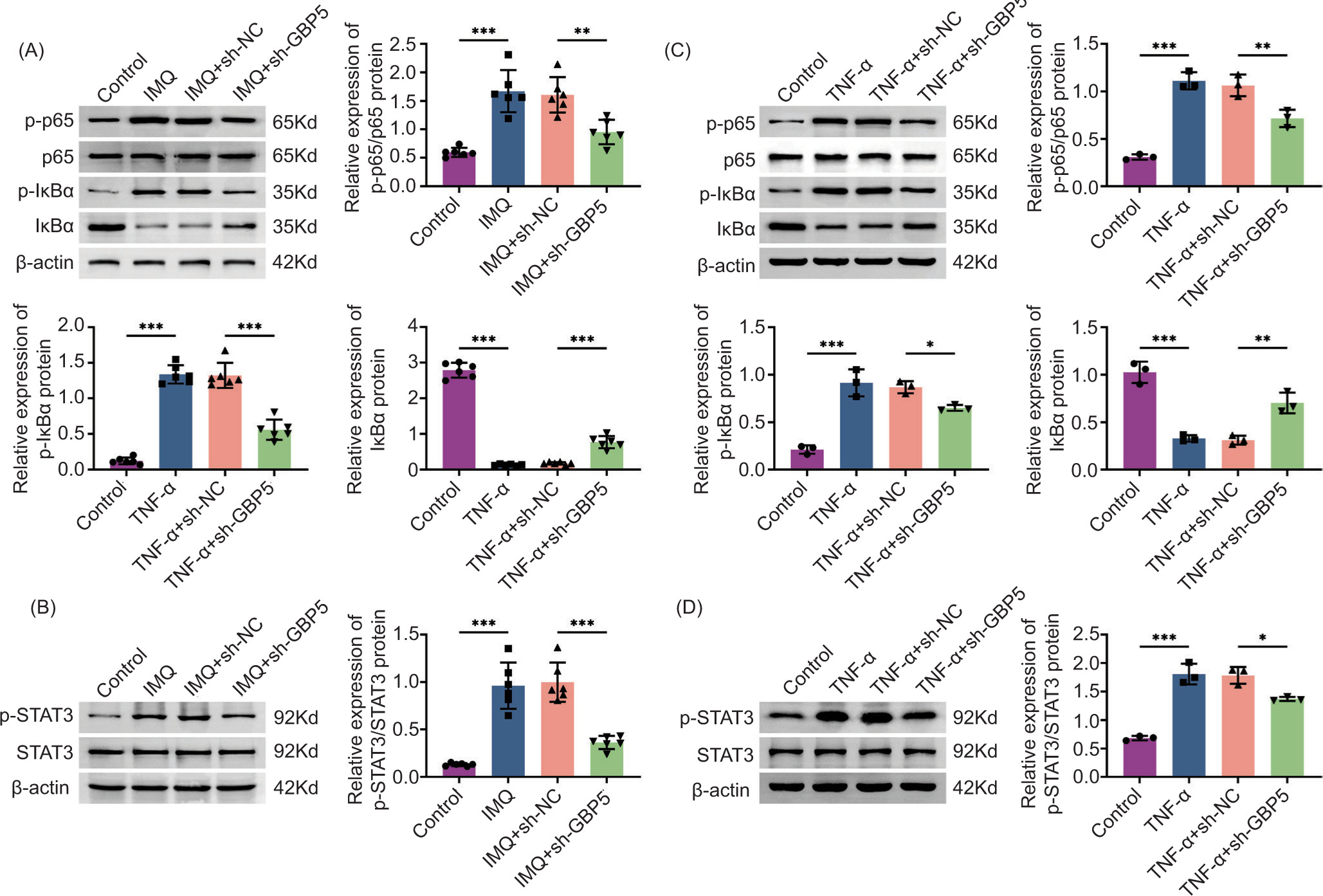
Knockdown of GBP5 regulates the NF-κB/STAT3 pathway in IMQ-induced mice and TNF-α-induced HaCaT cells
In order to elucidate the mechanism by which GBP5 influences inflammation and tissue damage, we investigated its mechanism. Immunoblot analysis showed that the phosphorylation levels of p65 and IκBα were elevated in the IMQ group but reduced in the IMQ+sh-GBP5 group (Figure 5A). Similarly, the phosphorylation level of STAT3 was higher in the IMQ group but decreased in the IMQ+sh-GBP5 group (Figure 5B). In the cell model of TNF-α-induced HaCaT cells, we observed a consistent increase in the levels of phosphorylated p65, IκBα, and STAT3 in the TNF-α group. However, these levels were decreased in the TNF-α+sh-GBP5 group (Figures 5C and D). These results suggest that GBP5 knockdown suppresses the activation of NF-κB/STAT3 axis in IMQ-induced mice, which correlates with reduced inflammation and renal damage.
Figure 5 Knockdown of GBP5 regulates the NF-κB/STAT3 pathway in IMQ-induced mice and TNF-α-induced HaCaT cells. (A) Immunoblot analysis showing the relative expression of p-p65/p65 and p-IκBα/IκBα proteins in kidney tissues in control, IMQ, IMQ+sh-NC, and IMQ+sh-GBP5 groups. The expression and phosphorylation levels of the indicated proteins were measured. (B) Immunoblot analysis showing the relative expression of p-STAT3/STAT3 proteins in kidney tissues in control, IMQ, IMQ+sh-NC, and IMQ+sh-GBP5 groups. The expression and phosphorylation levels of the indicated proteins were measured. (C) Immunoblot analysis showing the relative expression of p-p65/p65 and p-IκBα/IκBα proteins in HaCaT cells in control, TNF-α, TNF-α+sh-NC, and TNF-α+sh-GBP5 groups. The expression and phosphorylation levels of the indicated proteins were measured. (D) Immunoblot analysis showing the relative expression of p-STAT3/STAT3 proteins in HaCaT cells in control, TNF-α, TNF-α+sh-NC, and TNF-α+sh-GBP5 groups. The expression and phosphorylation levels of the indicated proteins were measured. Data are presented as mean±SD. **P < 0.01, ***P < 0.001, compared to control.
Discussion
Psoriasis is a type of chronic inflammatory skin disease featured by keratinocyte growth, altered differentiation, and extensive infiltration of immune cells. Psoriasis impacts roughly 2% of population globally, presenting a considerable challenge because of its persistent and recurring characteristics. Moreover, PRRD is a lesser-known but crucial issue that exacerbates the overall health implications of the disease for patients.17 Understanding the molecular mechanisms underlying both psoriasis and PRRD is essential for developing effective treatments. Our study focused on GBP5 and its regulatory role in NF-κB/STAT3 axis.
Proliferation and inflammation are central to the pathogenesis of psoriasis and PRRD.18 Psoriasis is characterized by the excessive growth of keratinocytes, leading to the formation of thick and scaly patches on the skin. At the same time, the inflammatory processes in the body contribute to the persistence and worsening of the condition. Inflammatory cytokines are significantly elevated in psoriasis, promoting keratinocyte proliferation and recruiting immune cells to the skin. Our study demonstrated that knocking down of GBP5 significantly reduced TNF-α-induced growth as well as inflammation in HaCaT cells. The findings are consistent with the prior research that suggested that inflammatory cytokines had a role in promoting both skin thickening and kidney impairment in psoriatic patients. Moreover, studies have demonstrated that the activated inflammatory pathways in psoriasis can have systemic repercussions, such as kidney damage, as a consequence of the persistent inflammatory response throughout the body.
GBP5 is a part of the interferon-induced GTPase family, involved in immune and inflammatory responses. GBP5 has been linked to a range of diseases, including lupus nephritis and rosacea, where it worsens inflammatory conditions. Additionally, GBP5 enhances the activation of inflammasomes and the secretion of pro-inflammatory cytokines, thereby playing a role in the development of disease.19 In lupus nephritis, GBP5 expression is upregulated in the renal cortex, and its inhibition has been shown to ameliorate renal damage. Similarly, in rosacea, GBP5 drives macrophage polarization towards a pro-inflammatory M1 phenotype, thus aggravating skin inflammation. Prior to this study, the involvement of GBP5 in psoriasis and PRRD was not investigated. We found that GBP5 is markedly upregulated in psoriatic tissues, and its silencing it leads to the mitigation of psoriasis manifestations and renal impairment in an IMQ-induced mouse modelinduced by IMQ. This finding highlights GBP5’s significant impact on cellular processes and inflammation, making it a promising target for therapeutic intervention.
In the context of inflammatory diseases, intricate pathways frequently come into play, with GBP5 emerging as a key player. GBP5 is known to intensify inflammation by promoting shift toward the pro-inflammatory M1 phenotype through NF-κB axis.15,20 Our research confirmed that GBP5 knockdown leads to reduced inflammation in psoriasis and PRRD, demonstrating that GBP5 is a key mediator in these conditions. This finding aligns with previous studies indicating that GBP5 inhibition can ameliorate inflammatory responses and tissue damage in various disease models. For instance, GBP5 is vital in osteoarthritis by promoting chondrocyte pyroptosis through IRF1/GBP5 axis, further underscoring its involvement in inflammatory processes.
The NF-κB/STAT3 pathway plays a crucial role in regulating inflammation and is linked to the development of psoriasis and PRRD. Activation of NF-κB results in the synthesis of inflammatory cytokines whereas the signaling of STAT3 is linked to the survival and growth of cells. Our study showed that GBP5 knockdown inhibits the activation of this pathway, resulting in decreased levels of p-p65, p-IκBα, and p-STAT3. This inhibition correlates with reduced inflammation and improved renal function, highlighting the pathway’s critical role in disease progression as well as the therapeutic potential of targeting GBP5. Earlier research showed that inhibiting the NF-κB/STAT3 axis reduced inflammation and disease severity in psoriasis, which aligned with our results.
Despite the promising results, our study has limitations. The sample size was relatively limited, calling for additional research involving larger cohorts. Moreover, although we illustrated the effects of reducing GBP5 in a mouse model and cell lines, clinical investigations are essential to assess its therapeutic applicability in human subjects. Future research should also assess the long-term effects and safety of GBP5-targeted therapies. Furthermore, delving into the interplay between GBP5 and other pathways could offer a more holistic insight into its involvement in psoriasis and PRRD.
Conclusion
Our study identified GBP5 as a novel therapeutic target for psoriasis and PRRD, providing insights into its role in regulating the NF-κB/STAT3 signaling pathway. These findings could have significant implications for improving patient outcomes and advancing our understanding of inflammatory diseases.
Availability of Data and Materials
All data generated and analyzed in this study are included in this published article. The datasets used and/or analyzed in the present study are available from the corresponding author on reasonable request.
Competing Interests
The authors stated that there was no conflict of interest to disclose.
Statement of Informed Consent
Written informed consent was obtained from a legally authorized representative(s) for anonymized patient information published in this article.
Author Contributions
Fang Ren and Jin Li designed the study, completed the experiment, and supervised data collection. Xingyan Ni analyzed and interpreted the data. Hongshan Yuan and Wenliang Yan prepared the manuscript for publication and reviewed its draft. All authors read and approved the manuscript.
REFERENCES
1. Griffiths CEM, Armstrong AW, Gudjonsson JE, Barker J. Psoriasis. Lancet. 2021;397(10281):1301–15. 10.1016/S0140-6736(20)32549-6
2. Rendon A, Schakel K. Psoriasis pathogenesis and treatment. Int J Mol Sci. 2019;20(6):1475. 10.3390/ijms20061475
3. Armstrong AW, Read C. Pathophysiology, clinical presentation, and treatment of psoriasis: A review. JAMA. 2020;323(19):1945–60. 10.1001/jama.2020.4006
4. Langley RG, Krueger GG, Griffiths CE. Psoriasis: Epidemiology, clinical features, and quality of life. Ann Rheum Dis. 2005;64(Suppl 2):ii18–23; discussion ii4–5. 10.1136/ard.2004.033217
5. Krupka-Olek M, Bozek A, Drewienkowska M, Dryjka K, Medrala A. Immunological differences between atopic dermatitis, psoriasis, and their combination in adult patients. Allergol Immunopathol (Madr). 2022;50(4):143–6. 10.15586/aei.v50i4.557
6. Ahamad N, Kar A, Mehta S, Dewani M, Ravichandran V, Bhardwaj P, et al. Immunomodulatory nanosystems for treating inflammatory diseases. Biomaterials. 2021;274:120875. 10.1016/j.biomaterials.2021.120875
7. Ka-Eun Kim N-RS, Soon-Hi Park, Sang-Yun Nam, Yong-Soon Yoon, Soon-Kwon Park, Jeong-Sook Park, Il-Young Cho. Modulation of the human immune status by spinal thermal massage: A non-randomized controlled study. Signa Vitae. 2021;18(2):137–46. 10.22514/sv.2021.144
8. Tashiro T, Sawada Y. Psoriasis and systemic inflammatory disorders. Int J Mol Sci. 2022;23(8):4457. 10.3390/ijms23084457
9. Yu H, Lin L, Zhang Z, Zhang H, Hu H. Targeting NF-kappaB pathway for the therapy of diseases: Mechanism and clinical study. Signal Transduct Target Ther. 2020;5(1):209. 10.1038/s41392-020-00312-6
10. Salas A, Hernandez-Rocha C, Duijvestein M, Faubion W, McGovern D, Vermeire S, et al. JAK-STAT pathway targeting for the treatment of inflammatory bowel disease. Nat Rev Gastroenterol Hepatol. 2020;17(6):323–37. 10.1038/s41575-020-0273-0
11. Zhang D, Jing B, Chen ZN, Li X, Shi HM, Zheng YC, et al. Ferulic acid alleviates sciatica by inhibiting neuroinflammation and promoting nerve repair via the TLR4/NF-kappaB pathway. CNS Neurosci Ther. 2023;29(4):1000–11. 10.1111/cns.14060
12. Xu Z, Li J, Zhou K, Wang K, Hu H, Hu Y, et al. Exocarpium citri grandis ameliorates LPS-induced acute lung injury by suppressing inflammation, NLRP3 inflammasome, and ferroptosis. J Ethnopharmacol. 2024;329:118162. 10.1016/j.jep.2024.118162
13. Li JM, Deng HS, Yao YD, Wang WT, Hu JQ, Dong Y, et al. Sinomenine ameliorates collagen-induced arthritis in mice by targeting GBP5 and regulating the P2X7 receptor to suppress NLRP3-related signaling pathways. Acta Pharmacol Sin. 2023;44(12):2504–24. 10.1038/s41401-023-01124-4
14. Liu N, Gao Y, Liu Y, Liu D. GBP5 inhibition ameliorates the progression of lupus nephritis by suppressing NLRP3 inflammasome activation. Immunol Invest. 2023;52(1):52–66. 10.1080/08820139.2022.2122834
15. Zhou L, Zhao H, Zhao H, Meng X, Zhao Z, Xie H, et al. GBP5 exacerbates rosacea-like skin inflammation by skewing macrophage polarization towards M1 phenotype through the NF-kappaB signalling pathway. J Eur Acad Dermatol Venereol. 2023;37(4):796–809. 10.1111/jdv.18725
16. Cao FY, Wang CH, Li X, Ma MZ, Tao GC, Yang C, et al. Guanylate binding protein 5 accelerates gastric cancer progression via the JAK1-STAT1/GBP5/CXCL8 positive feedback loop. Am J Cancer Res. 2023;13(4):1310–28.
17. Xu J, Chen A, Peng Y, Wen Z, Huang K, Wang P. A case report of adalimumab successfully treated a severe plaque psoriasis patient with psoriatic end-stage renal disease. Clin Cosmet Investig Dermatol. 2023;16:2841–6. 10.2147/CCID.S420900
18. Thatikonda S, Pooladanda V, Sigalapalli DK, Godugu C. Piperlongumine regulates epigenetic modulation and alleviates psoriasis-like skin inflammation via inhibition of hyperproliferation and inflammation. Cell Death Dis. 2020;11(1):21. 10.1038/s41419-019-2212-y
19. Ding K, Li X, Ren X, Ding N, Tao L, Dong X, et al. GBP5 promotes liver injury and inflammation by inducing hepatocyte apoptosis. FASEB J. 2022;36(1):e22119. 10.1096/fj.202101448R
20. Zhong K, Liu K, Song Y, Chen S, Hu X, Xue R, et al. A synthetic steroid 5 alpha-androst-3beta, 5, 6beta-triol alleviates radiation-induced brain injury in mice via inhibiting GBP5/NF-kappaB/NLRP3 signal axis. Mol Neurobiol. 2024;61(7):4074–89. 10.1007/s12035-023-03831-9
Supplementary
Figure S1 Illustration of the time and dose pattern of injection administration to animals.