
Download

ORIGINAL ARTICLE
Isoorientin alleviates ovalbumin-stimulated allergic rhinitis in mice by restoring Th1/Th2 balance
Jianyi Huanga, Ran Jib*, Xueshen Qianc, Yuli Shend
aDepartment of Traditional Chinese Medicine Classics, Taizhou Hospital of Traditional Chinese Medicine, Taizhou, China
bDepartment of E.N.T., Taizhou Integrated Traditional Chinese and Western Medicine Hospital, Taizhou, China
cClinical Laboratory, Taizhou Hospital of Traditional Chinese Medicine, Taizhou, China
dDepartment of E.N.T., Taizhou Hospital of Traditional Chinese Medicine, Taizhou, China
Abstract
Allergic rhinitis (AR) is a chronic, non-infectious inflammatory condition of the nasal mucosa mediated by IgE. There is a need for the development of novel medications to treat this ailment. Isoorientin is a naturally occurring flavonoid that possesses antioxidant, anti--inflammatory, and various other advantageous characteristics. However, its potential effects on AR remain unclear. This study evaluates the therapeutic effects of isoorientin on ovalbumin (OVA)-induced allergic rhinitis (AR) in mice and explores the underlying mechanism. Our study revealed that isoorientin administration effectively decreased the frequency of nose rubbing and sneezing in AR mice. The groups treated with isoorientin showed a significant decrease in serum levels of IgE and histamine, with reductions of 40% and 30%, respectively. Isoorientin ameliorated inflammation of the nasal mucosa and restored the Th1/Th2 balance. In addition, isoorientin inhibited the activation of the NF-κB pathway in nasal tissues. In summary, Isoorientin alleviates OVA-stimulated AR in mice by restoring Th1/Th2 balance and blocking the NF-κB pathway. Thus, isoorientin exhibits promise as a natural therapeutic agent for allergic rhinitis.
Key words: Allergic Rhinitis, Isoorientin, Nf-Κb Pathway, Natural Therapeutic Agent, Th1/Th2 Balance
*Corresponding author: Ran Ji, Department of E.N.T., Taizhou Integrated Traditional Chinese and Western Medicine Hospital, No.111, Jiangzhou South Road, Hailing District, Taizhou City, Jiangsu Province, China. Email address: [email protected]
Received 13 June 2024; Accepted 5 August 2024; Available online 1 September 2024
Copyright: Huang J, et al.
License: This open access article is licensed under Creative Commons Attribution 4.0 International (CC BY 4.0). http://creativecommons.org/licenses/by/4.0/
Introduction
Allergic rhinitis (AR) is a chronic, non-infectious inflammatory condition of the nasal mucosa mediated by IgE.1 This condition is triggered in genetically predisposed individuals upon exposure to allergens.2 It is important to note that systemic steroids are not recommended as maintenance therapy for AR due to potential side effects and the risk of symptom relapse upon discontinuation.3 AR affects an estimated 8.7–24.1% of the population in China.1 Current therapeutic approaches for allergic rhinitis (AR) mostly consist of corticosteroids and antihistamines.4 While corticosteroids are highly effective for most individuals, their prolonged usage at high dosages is restricted due to systemic side effects.4 Moreover, corticosteroids do not cure AR, as symptoms often relapse upon discontinuation.3 Antihistamines can alleviate sneezing, itching, and rhinorrhea but are less effective for nasal congestion.5 In addition, anticholinergic side effects such as fatigue, as well as depression, are associated with various antihistamine treatments.6 For these reasons, there is a practical need to develop new AR treatment strategies derived from safe natural products.
The imbalance between T-helper type 1 (Th1) and T-helper type 2 (Th2) cells is considered a major regulator of immunoglobulin E (IgE)-mediated allergic inflammation.4 Antigen-presenting cells present allergen to T lymphocytes when individuals are exposed to specific concentrations of allergens in the environment for an extended period.2 These T cells release cytokines that stimulate B cells to differentiate into cells, ultimately promoting the production of IgE while blocking the Th1 response.7 When IgE antibodies bind to receptors on mast cells, they cause the release of inflammatory cytokine, thereby stimulating a Th2 response.7 AR is characterized by an increase in Th2 cells and a decrease in Th1 cells.
Allergen immunotherapy (AIT) is an essential therapeutic option for allergic rhinitis (AR), especially for patients who do not have a sufficient response to traditional therapies or want to decrease their long-term medication usage. AIT is used in conjunction with pharmacotherapy to provide comprehensive treatment for AR. Allergen immunotherapy (AIT) is a treatment that involves gradually increasing the doses of the specific allergen to develop tolerance. It has been proven to alter the underlying process of allergic disease. This therapy offers extended relief and is the sole therapeutic method capable of modifying the inherent progression of AR.
Isoorientin is a naturally-occurring flavonoid found in several plants, including bamboo leaves, hawthorn, black buckwheat, and passionflower. It is classified as a luteolin 6-C-β-D-glucoside.8,9 It possesses multiple biological potencies, including anti-inflammatory and antibacterial effects.10–13 Studies have indicated that isoorientin might provide therapeutic benefits for allergic airway diseases.8 The underlying mechanisms are associated with a reduction in Th2 cytokines, activation of the Nrf2/HO-1 pathway, and attenuation of the NF-κB pathway.14 Nevertheless, the impact and underlying processes of isoorientin on AR are not yet well understood.
This study investigates the potential therapeutic effects of isoorientin on OVA-stimulated AR in C57BL/6 mice. The findings of this research could provide a basis for developing new treatment strategies for AR using naturally occurring compounds.
Materials and Methods
Materials
Animals
C57BL/6 mice (18–20 g, 6–8 weeks old) were purchased from Vital River. All animal experiments were conducted following the guidelines of the Ethics Committee of Taizhou Hospital of traditional Chinese Medicine. A total of 40 mice were used in this experiment. Mice were randomly divided into four groups with 10 mice in each group: Control, OVA, OVA+ISO (25 mg/kg), and OVA+ISO (50 mg/kg). At the end of the experiment, mice were euthanized by CO2 inhalation followed by cervical dislocation to ensure death.
Animal model and treatment
Mice were sensitized with an injection of 100 µL saline containing 50 µg OVA (Sigma-Aldrich, Cat# A5503) and 2 mg aluminum hydroxide (Thermo Fisher, Cat# 77161) on days 0, 7, and 14. From days 21 to 28, mice received daily administration of 20 µL saline containing 400 µg OVA. The groups were as follows: (i) control group (CON): mice in this group were injected with 100 µL saline without OVA and did not receive any isoorientin treatment. (ii) OVA group (OVA): mice in this group were injected with 100 µL saline containing 50 µg OVA and 2 mg aluminum hydroxide on days 0, 7, and 14. From days 21 to 28, they received daily administration of 20 µL saline containing 400 µg OVA but no isoorientin treatment. (iii) OVA+ISO 25 group (OVA+ISO 25): Mice in this group were injected with 100 µL saline containing 50 µg OVA and 2 mg aluminum hydroxide on days 0, 7, and 14. From days 21 to 28, they received a daily administration of 20 µL saline containing 400 µg OVA. In addition, they received isoorientin (Sigma-Aldrich, Cat# I9891) at a dose of 25 mg/kg orally from days 21 to 28. (iv) OVA+ISO 50 group (OVA + ISO 50): Mice in this group were injected with 100 µL saline containing 50 µg OVA and 2 mg aluminum hydroxide on days 0, 7, and 14. From days 21 to 28, they received daily administration of 20 µL saline containing 400 µg OVA. In addition, they received isoorientin (Sigma-Aldrich, Cat# I9891) at a dose of 50 mg/kg orally from days 21 to 28.
Methods
Evaluation of nasal symptoms
The number of sneezes and nasal rubbings was recorded over a period of 10 min following the last OVA challenge. Sneezes were counted by direct observation, and nasal rubbings were counted by observing how frequently the mice rubbed their noses against the cage walls.
Histopathological analysis
Nasal tissues were fixed in 10% formalin (Sigma-Aldrich, Cat# HT501128) for 24 h. Sections were deparaffinized in three changes of xylene (5 min each), and rehydrated through a series of graded alcohols (100%, 95%, 70%, and 50% for 2 min each), followed by rinsing in distilled water (2 min). Sections were stained with hematoxylin solution (Sigma-Aldrich, Cat# MHS16) for 5 min, rinsed in running tap water for 5 min, differentiated by dipping in 0.3% acid alcohol (1% HCl in 70% ethanol) for a few seconds, and rinsed again in running tap water for 5 min. Bluing was carried out by immersing the sections in 0.2% ammonia water or saturated lithium carbonate solution for 1 min, followed by rinsing in running tap water for 5 min. Sections were then stained with eosin Y solution (Sigma-Aldrich, Cat# HT110216) for 2 min, briefly rinsed in distilled water to remove excess stain, dehydrated through a series of graded alcohols (50%, 70%, 95%, and 100%) for 2 min each, and cleared in three changes of xylene (5 min each), and mounted with a coverslip using a resinous mounting medium.
Measurement of serum inflammatory markers
Serum was collected from mice by cardiac puncture under anesthesia. Blood was allowed to clot at room temperature for 30 min and then centrifuged at 2000×g for 10 min to separate the serum. The serum was carefully collected and stored at −80°C until analysis. Histamine levels were measured using a histamine ELISA kit (Thermo Fisher, Cat# EIASY141). OVA-specific IgE levels were measured using an OVA-specific IgE ELISA kit (Abcam, Cat# ab157718). All assays were performed according to the manufacturer’s instructions, and absorbance was read at 450 nm using a microplate reader (BioTek, Cat# ELx808).
Cytokine analysis
Nasal lavage fluid was collected by gently flushing the nasal cavity with 1 mL of saline using a pipette. The lavage fluid was then centrifuged at 1500×g for 10 min at 4°C to remove cellular debris, and the supernatant was collected and stored at −80°C until analysis. Cytokine levels (including IL-4, IL-5, IL-13, TNF-α, IL-6, and IFN-γ were measured using ELISA kits (Abcam, Cat# ab46034 for IL-4, ab100689 for IL-5, ab100716 for IL-13, ab100747 for TNF-α, ab46027 for IL-6, and ab100689 for IFN-γ). Each sample was analyzed in duplicate, and the assays were performed according to the manufacturer’s protocols. The absorbance was read at 450 nm using a microplate reader (BioTek, Cat# ELx808).
Immunoblot analysis
Nasal tissues were homogenized in RIPA buffer containing protease and phosphatase inhibitors (Thermo Fisher, Cat# 78440). Protein was separated by SDS-PAGE and transferred to PVDF membranes. Membranes were blocked with 5% non-fat milk and incubated overnight at 4°C with antibodies: p-p65 (Abcam, Cat# ab76302), p65 (Abcam, Cat# ab32536), p-IκBα (Abcam, Cat# ab133462), IκBα (Abcam, Cat# ab32518), and β-actin (Abcam, Cat# ab8226). After washing, membranes were incubated with HRP-conjugated secondary antibody (Abcam, Cat# ab6721) for 1 h. Bands were visualized using ECL detection reagent (Beyotime) and quantified by ImageJ.
Statistical analysis
Data were organized and analyzed using GraphPad Prism version 8.0 (GraphPad Software, San Diego, CA, USA). All results are presented as mean±standard deviation (SD). For comparisons between multiple groups, one-way ANOVA followed by Tukey’s post hoc test was used to determine statistical significance. In cases where non-parametric data were encountered during statistical analysis, non-parametric tests were employed. Specifically, the Kruskal–Wallis test followed by Dunn’s post hoc test was used for multiple group comparisons, and the Mann–Whitney U test was used for comparisons between two groups. All statistical tests were two-sided, and a p-value of less than 0.05 was considered statistically significant.
Results
Isoorientin alleviates nasal symptoms and reduces IgG releasing in OVA-stimulated AR in mice
To investigate the effects of isoorientin on nasal symptoms and inflammatory markers in AR, the OVA-stimulated mice model was constructed. In the OVA group, the number of sneezes was significantly higher. Treatment with isoorientin at doses of 25 and 50 mg/kg reduced the number of sneezes (Figure 1A). Similarly, the number of nasal rubbings was higher in the OVA group, while treatment with isoorientin at both doses reduced the nasal rubbing frequency (Figure 1B). Furthermore, the OVA group exhibited increased levels of histamine in the serum. Administration of Isoorientin at doses of 25 and 50 mg/kg resulted in a reduction in histamine levels (Figure 1C). Additionally, OVA-specific IgE levels were higher in the OVA group. Treatment with Isoorientin at both doses decreased the IgE levels (Figure 1D). Thus, Isoorientin mitigated nasal symptoms and decreased the release of IgG in mice with OVA-stimulated allergic rhinitis.
Figure 1 Isoorientin alleviates nasal symptoms and reduces IgE release in OVA-stimulated allergic rhinitis in mice. (A) Number of sneezes recorded in different groups: Control, OVA, OVA+ISO (25 mg/kg), and OVA+ISO (50 mg/kg). (B) Number of nasal rubbings recorded in different groups, showing a significant decrease in nasal rubbing frequency in the OVA+ISO (25 mg/kg) and OVA+ISO (50 mg/kg) groups compared to the OVA group. (C) Serum histamine levels (ng/mL) measured in different groups, illustrating reduced histamine levels in OVA+ISO (25 mg/kg) and OVA+ISO (50 mg/kg) groups compared to the OVA group. (D) OVA-specific IgE levels (ng/mL) in the serum of different groups, indicating a decrease in IgE levels in the OVA+ISO (25 mg/kg) and OVA+ISO (50 mg/kg) groups compared to the OVA group. Data are presented as mean ± standard deviation (SD). Statistical significance is indicated as follows: ***p < 0.001. OVA, ovalbumin; ISO, isoorientin; IgE, immunoglobulin E; SD, standard deviation.
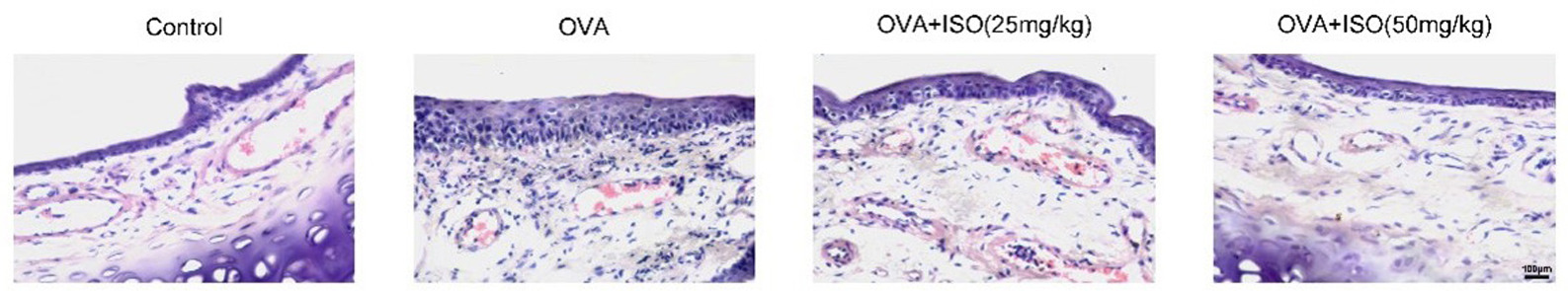
Isoorientin ameliorates histopathological changes in the nasal mucosa of OVA-stimulated AR mice
Next, we examined the effects of Isoorientin on histopathological changes in the nasal mucosa of OVA-stimulated mice using H&E staining. Histopathological analysis showed that in the control group, the nasal mucosa displayed a normal histological structure with mucosal thickness and immune cell infiltration (Figure 2). It indicates a thin epithelial layer and minimal immune cell infiltration in the nasal mucosa of the control group. In the OVA group, there was significant inflammatory cell infiltration and epithelial disruption (Figure 2). Treatment with isoorientin at doses of 25 and 50 mg/kg reduced inflammatory cell infiltration as well as improved epithelial structure compared to the OVA group (Figure 2). In summary, isoorientin ameliorated histopathological changes in the nasal mucosa of OVA-stimulated AR mice.
Figure 2 Isoorientin ameliorates histopathological changes in the nasal mucosa of OVA-stimulated allergic rhinitis mice. H&E staining showed histopathological changes in nasal mucosa sections from different experimental groups, including Control, OVA, OVA+ISO (25 mg/kg), and OVA+ISO (50 mg/kg). Scale bar, 100 μm. H&E, hematoxylin and eosin; OVA, ovalbumin; ISO, isoorientin.
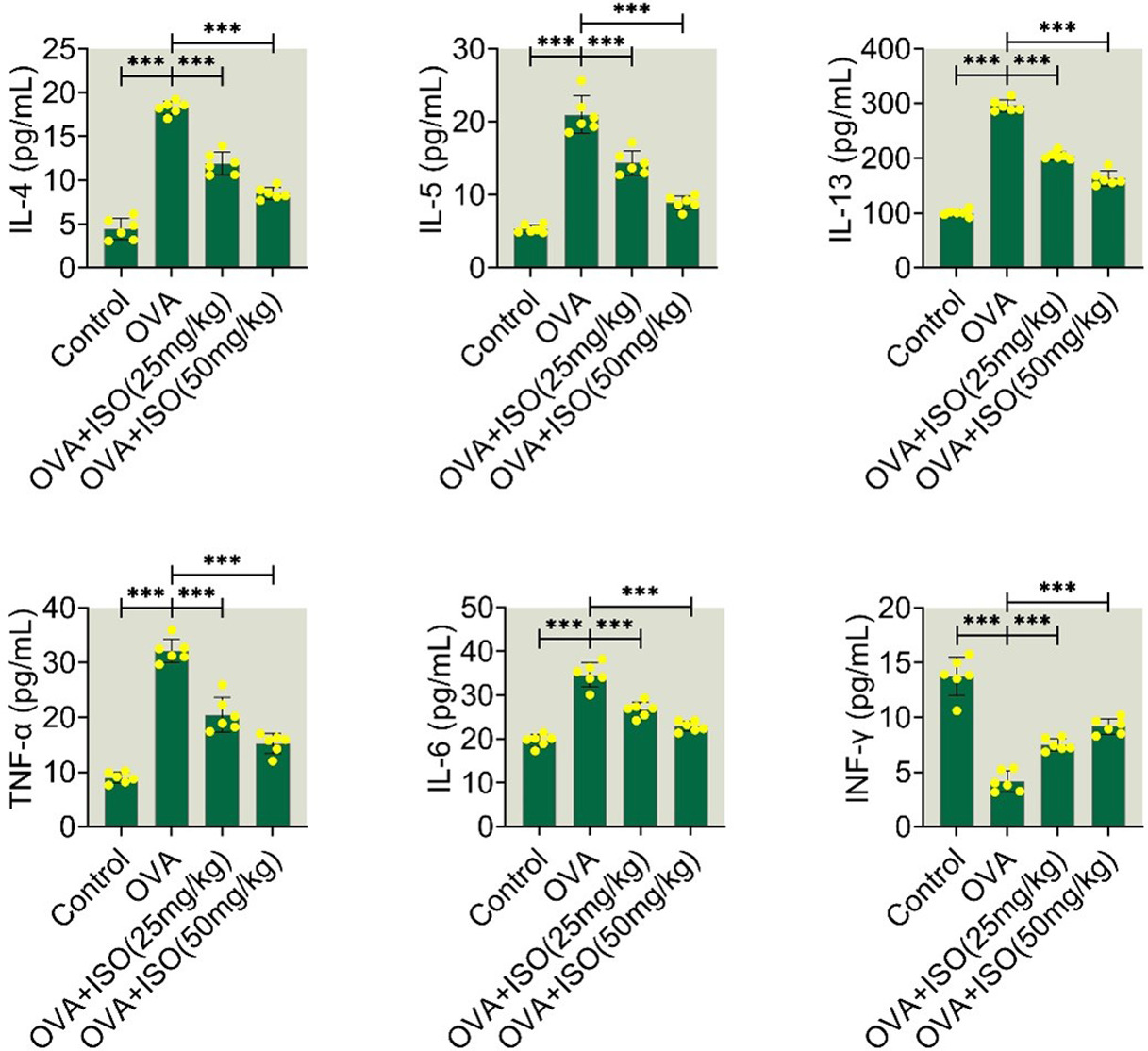
Isoorientin modulates cytokine levels in the nasal lavage fluid of OVA-stimulated allergic rhinitis mice
The effects of Isoorientin on the inflammatory response of OVA-stimulated mice were assessed through ELISA. Compared to the control group, the OVA group showed increased levels of IL-4, IL-5, IL-13, TNF-α, and IL-6, and decreased levels of IFN-γ, suggesting the promotion of inflammatory response (Figure 3). Treatment with isoorientin had an opposite result, suggesting suppression of the inflammatory response (Figure 3). Thus, isoorientin modulated cytokine levels in the nasal lavage fluid of OVA-stimulated AR mice.
Figure 3 Isoorientin modulates cytokine levels in the nasal lavage fluid of OVA-stimulated allergic rhinitis mice. The levels of IL-4, IL-5, IL-13, TNF-α, IL-6, and IFN-γ were measured through ELISA in the nasal lavage fluid of Control, OVA, OVA+ISO (25 mg/kg), and OVA+ISO (50 mg/kg) groups. Data are presented as mean±standard deviation (SD). Statistical significance is indicated as follows: ***p < 0.001. OVA, ovalbumin; ISO, isoorientin; IL, interleukin; TNF-α, tumor necrosis factor-alpha; IFN-γ, interferon-gamma; ELISA, enzyme-linked immunosorbent assay; SD, standard deviation.
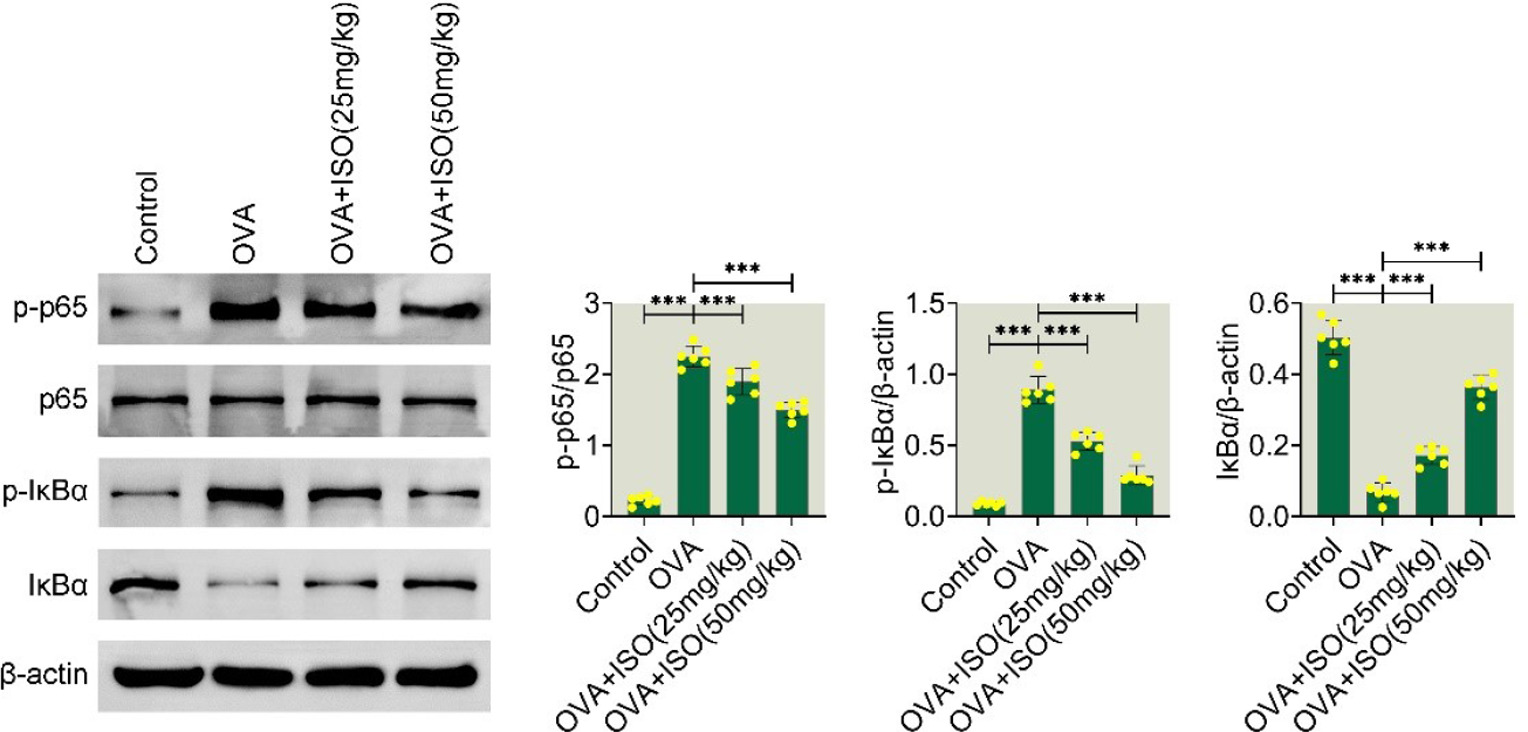
Isoorientin inhibits the NF-κB pathway in OVA-stimulated AR mice
We further investigated the possible mechanism underlying Isoorientin suppressing AR progression in mice. Immunoblot showed that the OVA group had increased levels of p-p65 and p-IκBα, along with decreased levels of IκBα (Figure 4). However, treatment with isoorientin reduced p-p65 and p-IκBα levels and increased IκBα levels, demonstrating an inhibition of the NF-κB pathway (Figure 4). Thus, isoorientin blocked the NF-κB pathway in OVA-stimulated AR mice.
Figure 4 Isoorientin inhibits the NF-κB signaling pathway in OVA-stimulated allergic rhinitis mice. The protein levels of phosphorylated p65 (p-p65), total p65, phosphorylated IκBα (p-IκBα), and total IκBα were analyzed in the Control, OVA, OVA + ISO (25 mg/kg), and OVA + ISO (50 mg/kg) groups. β-actin was used as a loading control. Quantification of the Immunoblot bands is presented in bar graphs for p-p65/p65, p-IκBα/β-actin, and IκBα/β-actin ratios. Data are presented as mean±standard deviation (SD). Statistical significance is indicated as follows: ***p < 0.001. NF-κB, nuclear factor kappa B; OVA, ovalbumin; ISO, isoorientin; IκBα, inhibitor of kappa B alpha; SD, standard deviation.
Discussion
Isoorientin, a naturally occurring flavonoid found in several plants such as bamboo leaves and hawthorn, possesses multiple biological activities.11 Previous studies have highlighted its therapeutic potential in various diseases, particularly allergic respiratory conditions such asthma.8,9,11 The mechanisms underlying these effects involve the reduction of Th2 cytokines, activation of the Nrf2/HO-1 pathway, and inhibition of the NF-κB signaling pathway.14 Our research extends these findings to AR, demonstrating that isoorientin can effectively mitigate allergic symptoms and inflammation in an AR model. The objective of our study is to rectify the Th1/Th2 equilibrium and establish a foundation for the development of novel treatments for allergic rhinitis.
Isoorientin significantly decreased inflammatory markers and alleviated nasal symptoms in mice with OVA-stimulated allergic rhinitis (AR), according to our findings. In addition, treatment with isoorientin resulted in a decrease in the occurrence of sneezing and nose rubbing, a reduction in the levels of IgE and histamine in the blood, improvement in the inflammation of the nasal mucosa, and restoration of the balance between Th1 and Th2 immune responses. Additionally, isoorientin inhibited the activation of the NF-κB pathway in nasal tissues. AR is triggered in genetically predisposed individuals upon exposure to environmental allergens.4,15 The prevalence of AR has been rising globally, affecting a significant portion of people worldwide and leading to substantial healthcare challenges.16 Existing therapies, such as corticosteroids and antihistamines, are constrained by drawbacks such as adverse effects and the recurrence of symptoms after cessation.17
The disparity between Th1 and Th2 cells plays a crucial role in IgE-mediated allergic inflammation.18 In AR, there is a notable increase in the population of Th2 cells and a corresponding reduction in the number of Th1 cells.7,19 This imbalance leads to an excessive production of IgE and subsequent allergic responses.7 Our study demonstrated that isoorientin could restore the Th1/Th2 balance in OVA-stimulated AR mice. This restoration was evidenced by increased levels of Th1 cytokines and decreased levels of Th2 cytokines, suggesting that targeting Th1/Th2 balance could be a viable therapeutic strategy for AR.
Inflammation plays a pivotal role in AR.20,21 Isoorientin’s anti-inflammatory properties are well-documented, and our study further elucidates its effects in AR.22 Our observation revealed that isoorientin effectively decreased nasal symptoms, such as nose rubbing and sneezing, in mice with allergic rhinitis. In addition, it reduced the concentration of IgE in the blood serum and the amount of histamine in the plasma. Histopathological analysis revealed that isoorientin ameliorated nasal mucosa inflammation, suggesting its potent anti-inflammatory effects. These results further highlight the potential of isoorientin as a natural anti-inflammatory agent for AR treatment.
The NF-κB pathway is vital in mediating immune responses.23 In AR, NF-κB activation contributes to inflammatory processes and Th1/Th2 imbalance.23 Our study showed that isoorientin inhibited the activation of the NF-κB pathway in nasal tissues of AR mice. This inhibition was indicated by decreased levels of p-p65 and p-IκBα, and increased levels of IκBα. By modulating the NF-κB pathway, isoorientin aided in restoring the Th1/Th2 balance and reduced inflammation, highlighting its therapeutic potential in managing AR.
Despite the promising results, our study has some limitations. The exact molecular mechanisms by which isoorientin exerts its effects in AR need further elucidation. In addition, our study was conducted in a murine model, and the findings need to be validated in human subjects. Future studies could focus on understanding the detailed mechanisms of isoorientin action and conducting clinical trials to confirm its efficacy and safety in humans. Investigating the long-term effects of isoorientin treatment on AR and exploring its potential synergistic effects with other AR treatments are also essential areas for future studies.
In conclusion, our study demonstrates that isoorientin significantly alleviates OVA-stimulated allergic rhinitis (AR) in mice by reducing nasal symptoms and inflammation, as well as by modulating Th1/Th2-related cytokines and inhibiting the NF-κB pathway. While these results suggest potential mechanisms, they do not conclusively establish that isoorientin ameliorates AR solely through the Th1/Th2 balance and NF-κB pathway. Further studies are needed to fully elucidate the exact molecular mechanisms and confirm these findings in human subjects.
Conflict of Interest
The authors declare no conflict of interest.
Ethics Approval
Ethical approval was obtained from the Ethics Committee of Taizhou Hospital of traditional Chinese Medicine.
Data Availability
The authors declare that all data supporting the findings of this study are available within the paper and any raw data can be obtained from the corresponding author upon request.
Author Contributions
JH and RJ designed the study and carried them out, JH, RJ, XQ, and YS supervised the data collection, JH, RJ, and XQ analyzed the data, JH, RJ, XQ, and YS interpreted the data, JH and RJ prepare the manuscript for publication and reviewed the draft of the manuscript. All authors have read and approved the manuscript.
REFERENCES
1. Zhang FZ, Tan M, Zeng J, Qi XW, Zhang YT, Che YT, et al. A supramolecular assembly of EGCG for long-term treatment of allergic rhinitis. ACS Biomater Sci Eng. 2024;10(4):2282–98. 10.1021/acsbiomaterials.4c00091
2. Ryu G, Bae JS, Kim JH, Kim EH, Chung YJ, Mo JH. Sneezing and rubbing counts in allergic rhinitis mouse models are a reliable indicator of type 2 immune response. Clin Exp Otorhinolaryngol. 2020;13(3):308–11. 10.21053/ceo.2019.02005
3. Yang Y, Wang L, Wang S, Wang Y, Du Y, Fan Y. Luteolin restored Treg/Th17 balance to ameliorate allergic rhinitis in a mouse model. Immunopharmacol Immunotoxicol. 2023;45(4):461–8. 10.1080/08923973.2023.2166527
4. Pfaar O, Gehrt F, Li H, Rudhart SA, Nastev A, Stuck BA, et al. Anti-IgE: A treatment option in allergic rhinitis? Allergol Select. 2021;5:119–27. 10.5414/ALX02205E
5. Hu L, He W, Li J, Miao Y, Liang H, Li Y. The role of adenoid immune phenotype in polysensitized children with allergic rhinitis and adenoid hypertrophy. Pediatr Allergy Immunol. 2024;35(6):e14166. 10.1111/pai.14166
6. Zhan J, Qi Y, Fu Y, Zheng J, Wu J, Wei X, et al. LncRNA ZFAS1 alleviated NLRP3 inflammasome-mediated pyroptosis through regulating miR-96-5p/Smad7 signaling in allergic rhinitis. Int Arch Allergy Immunol. 2024:1–14. 10.1159/000535646
7. Ke X, Chen Z, Wang X, Kang H, Hong S. Quercetin improves the imbalance of Th1/Th2 cells and Treg/Th17 cells to attenuate allergic rhinitis. Autoimmunity. 2023;56(1):2189133. 10.1080/08916934.2023.2189133
8. Liang S, Zhao Y, Chen G, Wang C. Isoorientin ameliorates OVA-induced asthma in a murine model of asthma. Exp Biol Med. 2022;247(16):1479–88. 10.1177/15353702221094505
9. Luo T, Jiang S, Zhou B, Song Q, Du J, Liu P, et al. Protective effect of isoorientin on oleic acid-induced oxidative damage and steatosis in rat liver cells. Front Pharmacol. 2022;13:818159. 10.3389/fphar.2022.818159
10. Mthembu SXH, Muller CJF, Dludla PV, Madoroba E, Kappo AP, Mazibuko-Mbeje SE. Rooibos flavonoids, aspalathin, isoorientin, and orientin ameliorate antimycin A-induced mitochondrial dysfunction by improving mitochondrial bioenergetics in cultured skeletal muscle cells. Molecules. 2021;26(20). 10.3390/molecules26206289
11. Sun X, Xue X, Wang X, Zhang C, Zheng D, Song W, et al. Natural variation of ZmCGT1 is responsible for isoorientin accumulation in maize silk. Plant J. 2022;109(1):64–76. 10.1111/tpj.15549
12. Huang S, Li Z, Jiang S, Xu M. Metabolomic study on the protective effect of isoorientin against myocardial infarction. Biochem Biophys Res Commun. 2022;598:81–8. 10.1016/j.bbrc.2022.02.008
13. Zhang T, Xiu YH, Xue H, Li YN, Cao JL, Hou WS, et al. A mechanism of isoorientin-induced apoptosis and migration inhibition in gastric cancer AGS cells. Pharmaceuticals. 2022;15(12). 10.3390/ph15121541
14. Zhang L, Zhu XZ, Badamjav R, Zhang JZ, Kou JP, Yu BY, et al. Isoorientin protects lipopolysaccharide-induced acute lung injury in mice via modulating Keap1/Nrf2-HO-1 and NLRP3 inflammasome pathways. Eur J Pharmacol. 2022;917:174748. 10.1016/j.ejphar.2022.174748
15. Huyin Yang DL, Yanping Wang. Physalin A exerts neuroprotective effects: inhibition of OGD/R-induced cellular pyroptosis and inflammatory responses in nerve cells. Signa Vitae. 2023;19(6):168–74.
16. Li HT, Chen ZG, Lin YS, Liu H, Ye J, Zou XL, et al. CpG-ODNs and budesonide act synergistically to improve allergic responses in combined allergic rhinitis and asthma syndrome induced by chronic exposure to ovalbumin by modulating the TSLP-DC-OX40L axis. Inflammation. 2018;41(4):1304–20. 10.1007/s10753-018-0779-6
17. Li E, Wang D, Xue Y, Yan J, Wang J. The protective role of cirsilineol against ovalbumin-induced allergic rhinitis in mice by suppression of inflammation and oxidative stress. J Environ Pathol Toxicol Oncol. 2021;40(3):63–73. 10.1615/JEnvironPatholToxicolOncol.2021038489
18. Tang KT, Lin CC, Lin SC, Wang JH, Tsai SW. Kurarinone attenuates collagen-induced arthritis in mice by inhibiting Th1/Th17 cell responses and oxidative stress. Int J Mol Sci. 2021;22(8). 10.3390/ijms22084002
19. Ren K, Pei J, Guo Y, Jiao Y, Xing H, Xie Y, et al. Regulated necrosis pathways: a potential target for ischemic stroke. Burns Trauma. 2023;11:tkad016. 10.1093/burnst/tkad016
20. Zheng Z, Wang P. Serum ATG5 concentration relates to Th2 cell, nasal symptoms and therapeutic outcomes in allergic rhinitis patients. Biomark Med. 2023;17(8):437–44. 10.2217/bmm-2023-0152
21. Hu X, Liu S, Jing Z, He Y, Qin G, Jiang L. Immunomodulation in allergic rhinitis: Insights from Th2 cells and NLRP3/IL-18 pathway. Cell Biochem Funct. 2024;42(3):e3997. 10.1002/cbf.3997
22. Cao Z, Liu W, Bi B, Wu H, Cheng G, Zhao Z. Isoorientin ameliorates osteoporosis and oxidative stress in postmenopausal rats. Pharm Biol. 2022;60(1):2219–28. 10.1080/13880209.2022.2142614
23. Liu J, Liu S, Pan W, Li Y. Wogonoside attenuates the articular cartilage injury and the infiltration of Th1/Th2-type cytokines in papain-induced osteoarthritis in rat model via inhibiting the NF-kappaB and ERK1/2 activation. Immunopharmacol Immunotoxicol. 2021;43(3):343–52. 10.1080/08923973.2021.1913503