
Download
ORIGINAL ARTICLE
Allergo-immunopathology mechanism of thymol-inhibiting airway remodeling in asthmatic mice by regulating TGF-β/Smad3 pathway
Liyuan Zhanga, Wenna Zhanga, Yanan Wanga, Pei Caia, Chaoran Lia, Yan Shia, Seyyed Shamsadin Atharib, Ailing Lia*
aDepartment of Respiratory and Critical Care Medicine, Xi’an International Medical Center Hospital, Xi’an, 710100, China
bDepartment of Immunology, School of Medicine, Zanjan University of Medical Sciences, Zanjan, Iran
Abstract
Allergic asthma is an important public health problem and is a complicated respiratory sickness that is characterized by bronchial inflammation, bronchoconstriction, and breathlessness. Asthma is orchestrated by type 2 immune response and remodeling is one of the important outputted problem in chronic asthma. Thymol is a naturally occurring monocyclic phenolic, it has a series of biological properties, and its immunomodulatory and anti-remodeling effects on allergic asthma were evaluated. The OVA-LPS-induced asthmatic mice were treated with thymol. Methacholine challenge test, eosinophil count, and levels of IL-4, IL-5, IL-13, and IL-33 in bronchoalveolar lavage fluid, total and OVA-specific IgE levels in serum, remodeling factors, gene expression of TGF-β, Smad2, Smad3, and lung histopathology were done. Treatment with thymol could control AHR, eosinophil percentage levels of Th2 cytokines and Igs, remodeling factors, expression of TGF-β, Smad2 and Smad3 genes, inflammation, goblet cell hyperplasia, and mucus production in asthmatic mice. Thymol can control asthma pathogens and related remodeling and fibrosis bio-factors and can be a potential treatment of asthma.
Key words: Signaling Pathways, Asthma, T helper, Drug, Remodeling
*Corresponding author: Ailing Li, Department of Respiratory and Critical Care Medicine, Xi’an International Medical Center Hospital, Xi’an, 710100, China. Email addresses: [email protected]; [email protected]
Received 7 May 2024; Accepted 24 July 2024; Available online 1 September 2024
Copyright: Zhang L, et al.
License: This open access article is licensed under Creative Commons Attribution 4.0 International (CC BY 4.0). http://creativecommons.org/licenses/by/4.0/
Key massage
Allergic asthma is an important public health problem and remodeling is one of the important outputted problem in chronic asthma.
Thymol is a naturally occurring monocyclic phenolic, and treatment of asthma with thymol could control remodeling, inflammation, and immune reactions.
Introduction
Asthma is an important and complicated respiratory sickness that has higher prevalence in the world and leads to more socioeconomic costs for its prevention, diagnosis, and treatment. Asthma is characterized by airway hyperresponsiveness (AHR), bronchial inflammation, bronchoconstriction, and recurrent attacks of breathlessness. Allergic asthma is an important public health problem and a major cause of respiratory morbidity and mortality. Asthmatic patients cannot be completely cured, and all produced treatments generally aim to control the pathological conditions.1–3
Asthma is orchestrated by type 2 immune response, and several pathways are identified in the allergic asthma pathophysiology. The imbalance between Th1 and Th2, inflammatory and anti-inflammatory factors, and also dysregulation of the lung immune system through the signaling pathways lead to pulmonary injury.4,5
There are many components that can activate toll-like receptors (TLRs) signaling and may have an effect on asthma pathogenesis. Lipopolysaccharide (LPS) is part of bacteria’s structure and ligand of TLR4. Activation of TLR4 is initiated through LPS/TLR4/NF-kB pathway. On the other hand, LPS induces acute lung inflammation that leads to inflammatory cell accumulation in the lung and airway. Ovalbumin (OVA) induces allergic asthma l, but OVA/LPS induces asthma and acute inflammation in airways.6,7
Thymol as a monoterpenic phenol is found in oil of thyme. Thymol (2-isopropyl-5-methylphenol) is a naturally occurring monocyclic phenolic compound derived from lamiaceae (thymus vulgaris), and it has many biological properties, including antispasmodic, antioxidant, anti-platelet aggregation and anti-inflammatory effects. Thymol reduces the DNA oxidative damage by H2O2. Thymol’s antioxidant potential has been demonstrated in vitro and in vivo in the form of nephrotoxicity and cardiotoxicity.8,9
However, several studies in the asthma model focused more on the anti-inflammatory effect of thymol, while there is little information about the effect of this substance on remodeling. In this study, immunomodulatory and anti-remodeling effects of thymol on allergic asthma pathophysiology in a mouse model were evaluated. Also, pulmonary pathological features and allegro-inflammatory airway response were studied.
Materials and methods
Experimental animal model
A total of 40 male BALB/c mice, aged 6–8 weeks, were acclimatized to standard laboratory conditions (free access to water and food, 55±15% humidity, 22±2⁰C temperature with 12 hour dark/light cycle). All experimental protocols were followed in accordance with the stipulations of the Institutional Animal Ethical Committee. The mice were allocated in four groups (N=10), namely, the negative control group [Control (normal saline administrated)]; asthma group (Asthma [OVA, aluminum hydroxide and (alum) and LPS administrated]; thymol group [Thymol, induced asthma (OVA, alum, LPS), and treated with intra-peritoneal injection of thymol]; drug control group [Drug, induced asthma (OVA, alum, LPS) and treated with intra-peritoneal injection of aminophylline saline solution].
OVA-LPS sensitization and challenge
During the study, all mice received food and water ad libitum. To produce the OVA-LPS asthma model, on days 1, 7, and 14, the mice were sensitized by intra-peritoneal injection of OVA emulsified in alum. On days 15, 17, and 19, the mice were challenged intra-nasally with OVA and LPS in saline and on days 21, 23, and 25, the mice were challenged with OVA aerosol in saline by a nebulizer.6 The negative control group only received normal saline following the same protocol. Treatments were done on days 20 and 22. and on day 27, sampling was done.
AHR Assessment
In each group, five mice were used for AHR, and five mice were used for histopathology and biological factors analyses. The AHR in mice were assessed in response to methacholine (MCH) challenge via intubation on day 27. AHR was measured by determining the enhanced pause (penh value) according to the previously described method.6
Cell count in BALF
After the last challenge, the mice were euthanized, tracheotomized bronchoalveolar lavage fluid (BALF) was collected, and differential cell count was done in cytospin slide (with Gimsa stain) to determine the eosinophil percentage. Firstly, BALF was cryo-centrifuged, the supernatant was separated, and the cell sediment was used for the preparation of slides.6,10,11
Cytokines
The supernatant of BALF was stored and used for the measurement of cytokines’ levels. The bioplex-multiplex method was used to determine levels of interleukin (IL)-4, IL-5, IL-13, and IL-33 in BALF’s supernatant.
Immunoglobulins
Blood samples were taken. After separation of serum, total and OVA-specific immunoglobulin (Ig)E levels were measured by specific ELISA kits following the manufacturer’s protocol.
Remodeling factors
In the lung tissue, content of hydroxyproline (HP) as an important index of collagen deposition was measured by a colorimetric modified method.10 Also, transforming growth factor-beta (TGF-β) as another remodeling factor was measured in homogenate lung tissue supernatants.10
Gene expression
To determine gene expression of three related molecules of remodeling signaling pathways, TGF-β, Smad2, and Smad3, quantitative real-time PCR was applied. From BALF cells, total RNA was extracted and reversed transcribed to cDNA and also, with specific primers, PCR was applied. GAPDH was used as an internal gene.
Histopathology
After interception and fixation of lung tissues, slides of the histological sections were prepared and stained with hematoxylin and eosin (H&E), periodic acid Schiff (PAS), Alcian blue, and AB-PAS stains. The sections were evaluated under the light microscopy for eosinophilic infiltration in and around bronchi and vessels, hyperplasia of the goblet cell, and mucus hypersecretion.6,10,11
Statistical Analysis
Result’s data were expressed as mean±SD, and correlation was analyzed using Pearson’s method. To analyze the differences between groups, the paired t-test was applied. P-value less than 0.05 was supposed to be significant. SPSS software was used for analysis and performed using GraphPad Prism to draw graphs.
Result
AHR Assessment
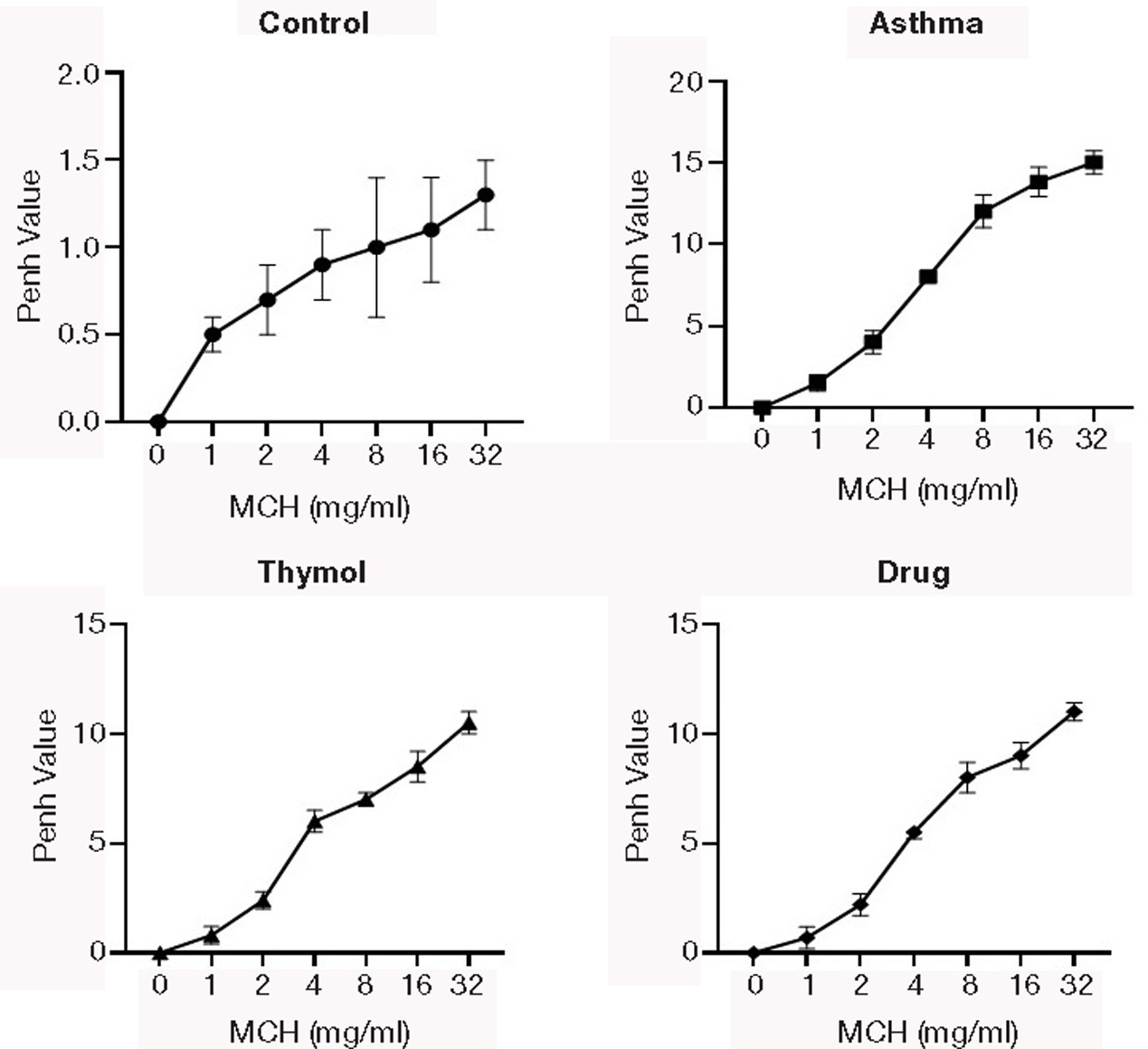
The penh value was increased in the asthma group compared to the control group in all concentrations of MCH (Figure 1). The penh value was decreased in drug and thymol groups compared to the asthma group in all concentrations of MCH. The penh value decreasing in the thymol group was similar to that in the drug group.
Figure 1. AHR. Methacholine (MCH) challenge test was done in all groups, and airway hyperresponsiveness was assessed in response to MHC as penh value.
Cell count in BALF
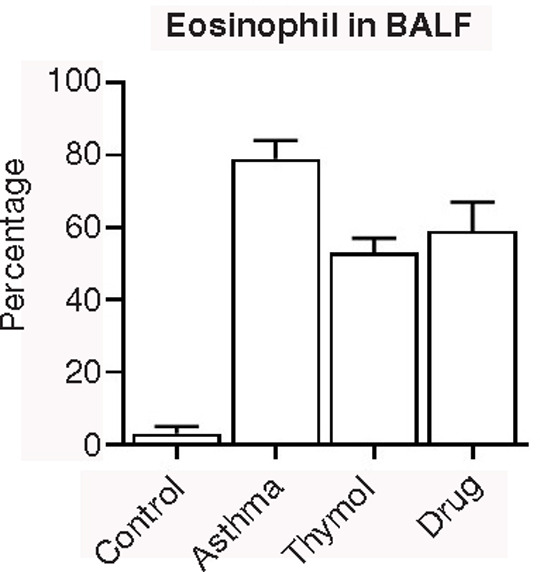
Percentage of eosinophil was increased in the asthma group (79±5%) compared to the control group (3±2%), and treatments in thymol (53±4%) and drug (59±8%) groups could decrease eosinophil’s percentage (Figure 2).
Figure 2. BALF Cells. The count of eosinophil in bronchoalveolar lavage fluid was done in all groups, and it was observed that treatments had a significant effect on eosinophil count.
Cytokines
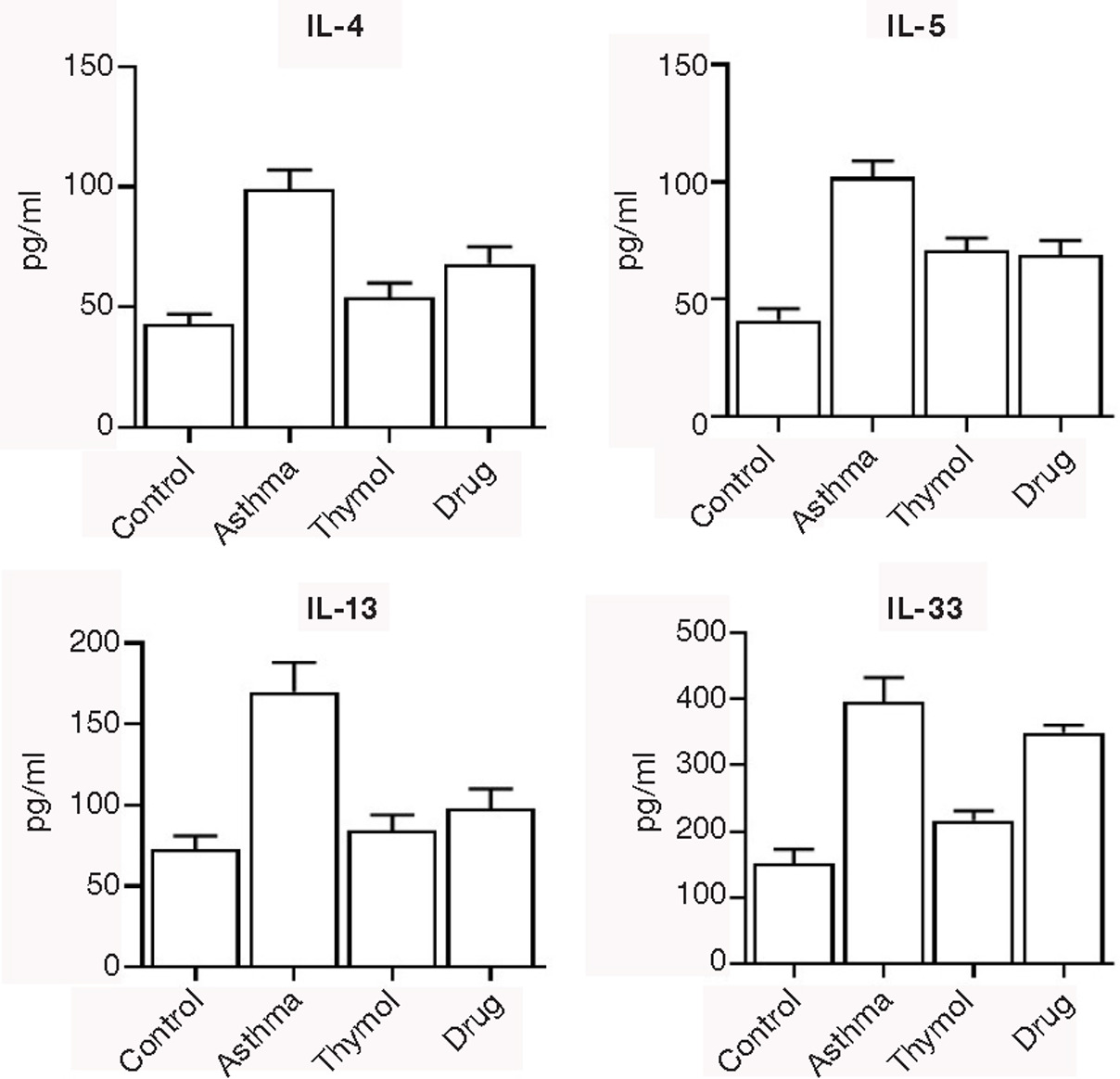
The main type 2 cytokines IL-4, 5, 13, and 33 were significantly (p<0.05) increased in the asthma group (99±8, 102±7, 170±18 and 395±37 pg/mL, respectively) compared to the control group (43±4, 41±5, 73±8 and 152±21 pg/mL, respectively). Increased levels of the main type 2 cytokines (IL-4, 5, 13, and 33) were significantly (p<0.05) controlled in thymol and drug groups (Figure 3).
Figure 3. Cytokines. The main Th2 cytokines levels, IL-4, -5, -13, and -33, were measured in the bronchoalveolar lavage fluid of all mice.
Immunoglobulins
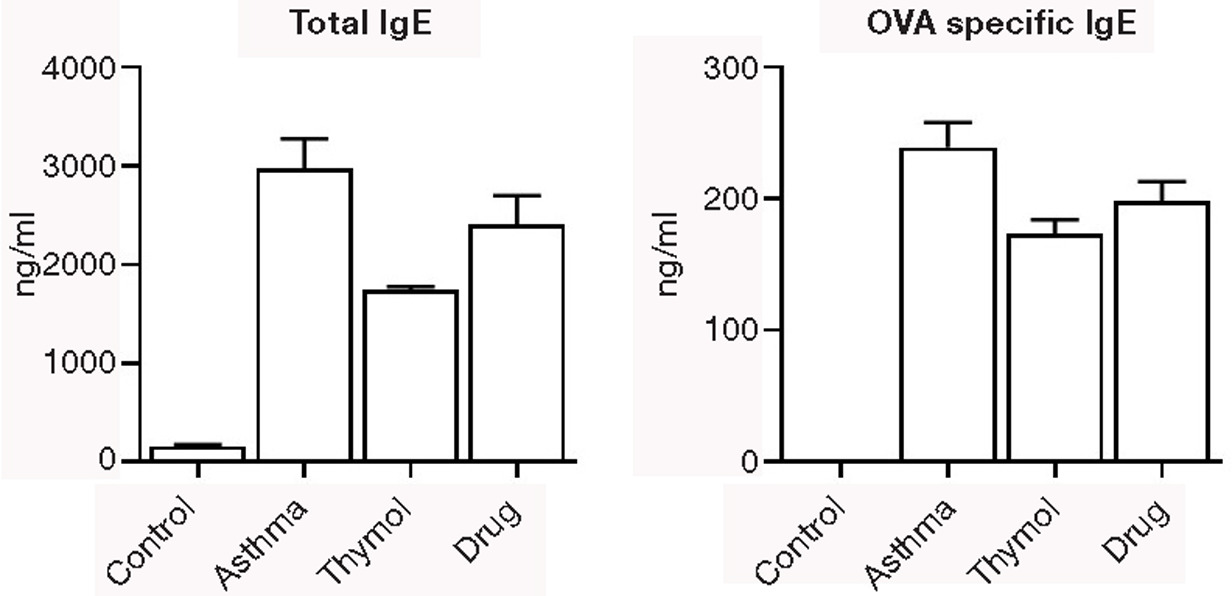
The main two allergic immunoglobulins (total and OVA-specific IgE) were significantly (p<0.05) increased in the asthma group (2974±301 and 239±19 ng/mL, respectively) compared to the control group (154±21 and 0±0 ng/mL, respectively). Increased levels of total and OVA-specific IgE were significantly (p<0.05) controlled in thymol and drug groups (Figure 4).
Figure 4. Immunoglobulin. The level of total and ovalbumin-specific IgE was measured in serum of all mice.
Remodeling factors
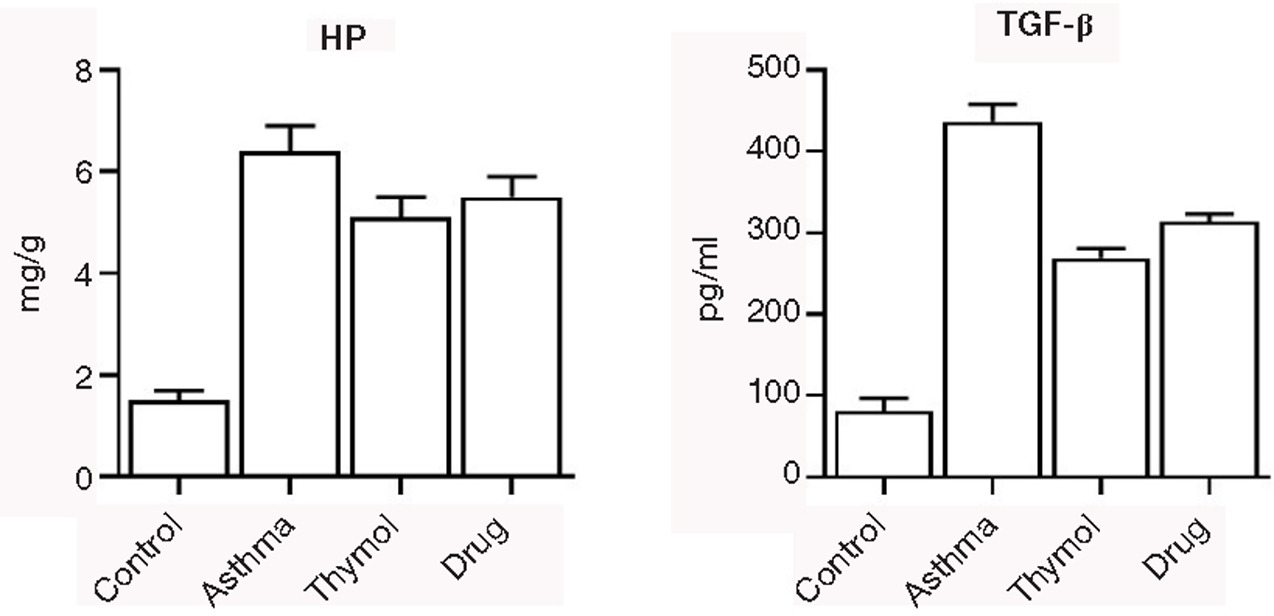
HP and TGF-β were elevated in the asthma group (6.4±0.5 mg/g and 437±21 pg/mL, respectively) significantly (p<0.05) compared to the control group (1.5±0.2 mg/g and 81±16 pg/mL, respectively). Treatments in two thymol and drug groups could decrease levels of HP and TGF-β significantly (p<0.05) compared to the asthma group (Figure 5).
Figure 5. Remodeling Biomarkers. Levels of hydroxyproline and TGF-β were measured in all groups, and it was observed that treatment with thymol could decrease levels of these biomarkers.
Gene expression
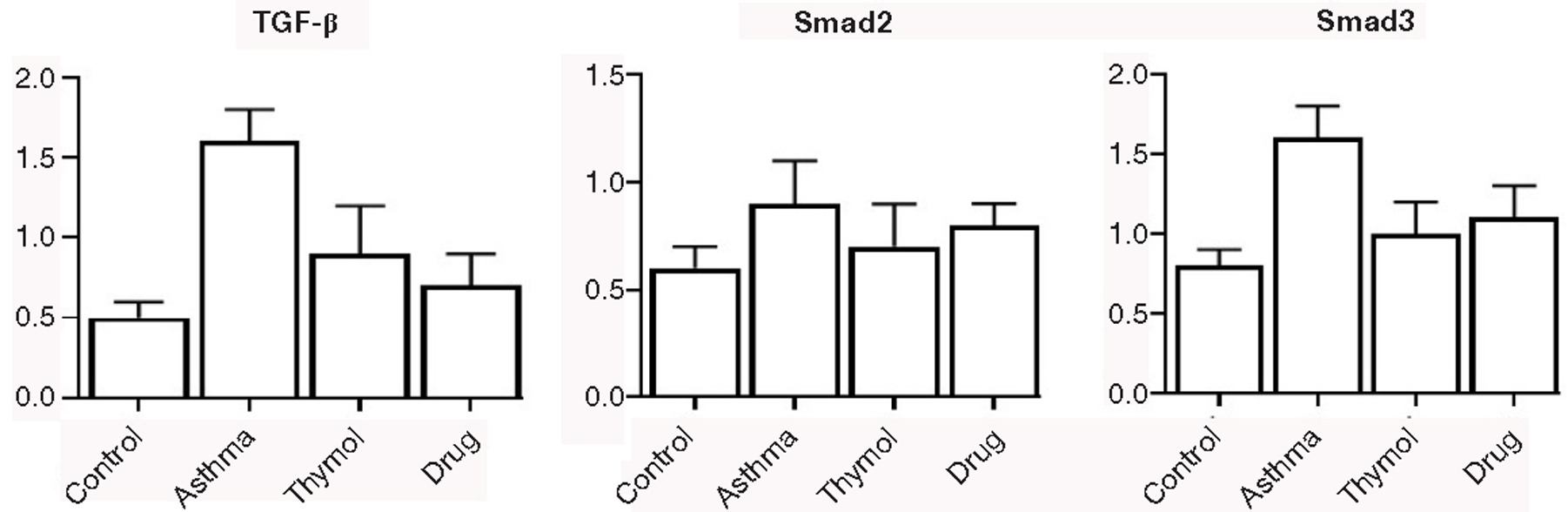
To evaluate the potential mechanisms of thymol treatment in reversing the asthma symptoms and inflammation in allergen-sensitized and challenged mice, the expressions of the genes, which were involved in the remodeling process (TGF-β, Smad2, and Smad3), were examined. Gene expressions of TGF-β, Smad2, and Smad3 were increased in the asthma group (1.6±0.2, 0.9±0.2, and 1.6±0.2). The thymol group could control gene expressions of TGF-β, Smad2, and Smad3 compared to the asthma group. The controlling was significant (p<0.05) in TGF-β and Smad3, but decreasing gene expressions of Smad2 in the thymol group was not significant (p>0.05) compared to the asthma group (Figure 6).
Figure 6. Gene expression. Expression of three genes that were involved in remodeling signaling pathways was studied in all mice.
Histopathology
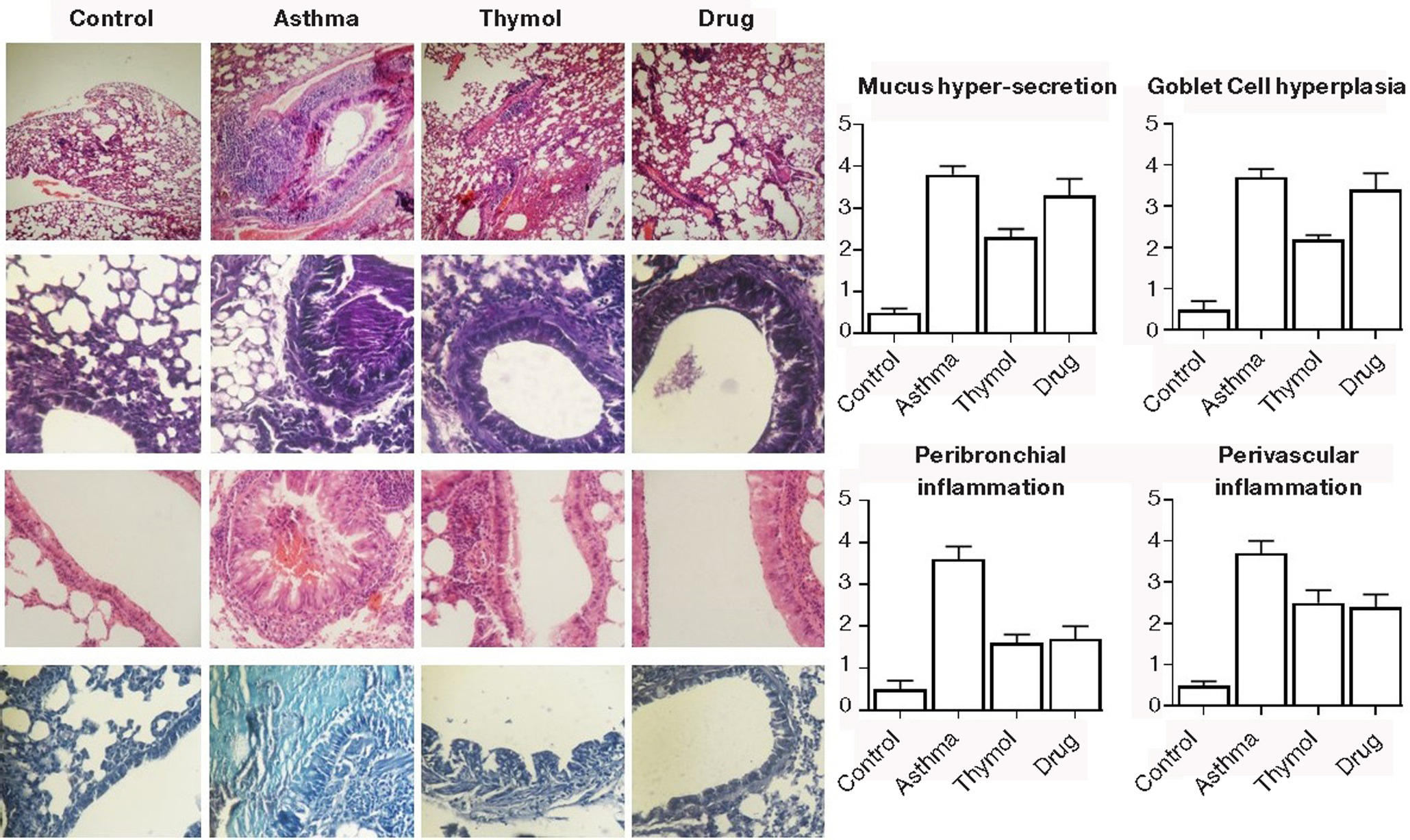
In a histopathological study of the lung tissue sections, pulmonary infiltration by inflammatory cells (especially eosinophils) into the peribronchial and perivascular regions, scores of goblet cell hyperplasia, and mucus production were increased in the asthma group (3.7±0.3, 3.6±0.3, 3.7±0.2, and 3.8±0.2) significantly (p<0.05) compared to the control group (0.5±0.1, 0.5±0.2, 0.5±0.2, and 0.5±0.1). Inflammation in the peribronchial and perivascular regions were significantly (p<0.05) controlled in thymol and drug groups (Figure 7). The goblet cell hyperplasia and mucus production were significantly (p<0.05) controlled only in the thymol group, and the controlling was not significant (p>0.05) in the drug group.
Figure 7. Pathology. Eosinophilic inflammation in perivascular and peribronchial metaplasia of goblet cells and mucus hyperproduction were studied in pathological sections.
Discussion
Allergy affects more than 30% of the global population. The prevalence of allergic diseases has been increasing in developed countries during the past two decades. Bronchial inflammation is the principal problem in asthma. Free oxygen radicals can play an important role in airway inflammation and allergic asthma pathogenesis, and antioxidative defense modulation is the new therapy for asthma.6,11 Bronchial epithelial cells not only act as a physical barrier but also play an important function in the process of immunity and inflammatory responses.12 In this study, AHR as the main concept of allergic reaction in airways was notable in the asthma group that was challenged with MCH. Using thymol for treatment could decrease AHR and response to the MCH challenge test. Also, percentage of eosinophil as the main cell in allergic asthma pathogenesis was decreased by thymol treatment in asthmatic mice.
Airway remodeling in asthma is the result of persistent inflammation that is a major driver of airway remodeling. Infiltrating inflammatory cells and damaged bronchial structure cells release large number of cytokines, growth factors, enzymes, and metabolites that result in remodeling of the airway. In addition, IL-13 induces profibrotic TGF-β releasing by epithelial cells. However, whether IL-17 is directly involved in airway remodeling and loss of Th17 cells leads to reduced collagen fibers staining, alpha-smooth muscle actin (α-SMA), and airway remodeling in chronic asthma.13,14 Pulmonary fibrosis has several pathological and genetic mechanisms. In lung fibrosis, thymol can modulate oxidative stress, inflammation, and PI3K/phospho-Akt signaling and exert anti-oxidant and anti-inflammatory effects, while decreasing TGF-β and PI3K/Akt signaling.15 HP and TGF-β as main remodeling trigger factors were elevated in the asthma group, and treatments with thymol could decrease the levels of HP and TGF-β significantly. Therefore, thymol can inhibit remodeling and in continues, lung fibrosis in asthmatic mice. Also, other main type 2 allergic cytokines (which have important role in asthma pathogenesis and airway obstruction) and their upper-hand cytokine, IL-33 were controlled in asthma group by treatment with thymol.
TGF-β is a crucial factor in the development, injury, and repair of lung tissue. Fibroblasts and myofibroblasts can secrete TGF-β1 that plays a critical role in the process of pathological pulmonary fibrosis. The TGF-β1/Smad3/p-Smad3 pathway is one of the mechanisms of fibrosis in the lung tissues.16,17 Various factors such as SERPINE1, SMAD6, SMAD7, TGFB1, and LTBP3 were reported as Smad2/3 target genes.18,19 The epithelium is the central component of bronchial inflammation and remodeling. The cellular biology of an epithelial transcriptional reprogramming event known as epithelial-mesenchymal transition (EMT) is one of the main processes in the lung that can be interfaced by growth factors and inflammation. TGF-β is the most potent inducer of EMT. It leads to disruption of the mucosal barrier function by reduced epithelial cadherin and disruption of epithelial adherens junctions. In addition, EMT enables to force transformed epithelial cells to express α-SMA fibers and intermediate filament vimentin, secretion of collagen and fibronectin, and expression of matrix metalloproteinases (MMPs) to promote airway remodeling. Although EMT (type II) is a protective mechanism to repair tissues, excessive and prolonged EMT processes can lead to lung fibrosis, and EMT plays an important role in the pathogenesis of asthma subtypes.20–23 TGF-β can bind to its receptor type II (TGFβRII) that recruits and phosphorylates TGFβRI. The activated TGFβRI/II complex recruits cytoplasmic transcription factors. Smads 2/3 are phosphorylated by TGFβRI, and phosphorylated Smad2/3 binds to Smad4 and then, the complex translocates to the nucleus. The complex binds to Smad-binding elements in jun-B and c-Jun, and modulates transcription with other coactivators including the cAMP response element–binding protein (CBP)/p300 histone acetyltransferases. TGF-β1 binding to its receptors stimulates signaling pathways including PI3K/Akt, p38, Wnt/β-catenin, Ras small GTPases, ERK, and JNK. All three MAPKs’ activation leads to cell proliferation, and fibronectin production is dependent on ERK1/2 and JNK.21–25 To evaluate the potential mechanisms of thymol treatment in reversing of inflammation in allergen-sensitized and challenged mice, it was observed that the expressions of the genes were involved in remodeling process; TGF-β, Smad2, and Smad3 were increased in the asthma group, and thymol could control gene expressions of TGF-β, Smad2, and Smad3. This control was significant with regard to TGF-β and Smad3 gene expressions.
Thymol has various biological properties, which can attenuate inflammatory cells infiltration.26 It was mentioned that thymol inhibited cell proliferation and has anti-tumoral activity through inhibition activity of Wnt/β-catenin pathway, as well as regulation of the β-catenin downstream targets, namely, c-myc, cyclin D1, and survivin. The β-catenin inhibition leads to the Wnt/β-catenin signaling downregulation, and thymol could downregulate the expression of Snail, vimentin, and N-cadherin. Thymol also inhibited invasion of colon cancer cells through the PI3K/AKT and ERK pathways activation.27 In a study, the results of examination showed that thymol reduced LPS-induced histopathology changes in the lung. Also, the expression of TNF-α, IL-1β, and IL-6 were suppressed, which indicated the protective role of thymol on LPS-induced inflammatory and oxidative response. Also, thymol inhibited LPS-induced NF-κB activation that plays an important role in the inflammatory cytokine production. Nrf2, a critical transcription factor, could have protection against pulmonary fibrosis, and thymol increased the expression of Nrf2.28 In a study, targeting of the reactive oxygen species with thymol could reduce inflammatory cell infiltration and the symptoms of inflammation in experimental asthma. Also, thymol recovered the antioxidant enzymes’ activity and glutathione content. The oxidative marker 8-OHdG increased in asthma, and thymol can reduce this marker. Thymol reduces oxidative damage to DNA by H2O2 at a nontoxic concentration.8 Oxidative stress changes protein, and releases peroxidases by activated eosinophils in the bronchi. Carbonyl protein creates asthma-related phenotype via enhancing mucin secretion, cytokines’ expression, and epithelial cells’ apoptosis. Therefore, carbonyl protein can be a suitable target for the treatment of asthma. In a study, thymol reduced carbonyl protein content in the asthma model. Lipid peroxidation is one of the oxidative properties in asthma that have important role in the leukocyte’s chemical attraction to the inflammation site. Studies showed that thymol reduces the malondialdehyde formation. Thymol can increase zinc that increases antioxidant and anti-inflammatory properties and improve asthma. Also, thyme’s antioxidant property increases the selenium level and has a protective effect against oxidative stress and asthma-induced inflammation.29–31 In a histopathological study, pulmonary infiltration by inflammatory cells (especially eosinophils) into peribronchial and perivascular regions was noted. Also, hyperplasia of goblet cell and mucus production were increased in asthmatic mice, and these factors were controlled in the thymol group. On the other hand, thymol could control increased levels of total and OVA-specific IgE, which are the two main allergic immunoglobulins. Therefore, thymol can control asthma pathogens and related factors of asthma pathophysiology. Also, remodeling and fibrosis-related biofactors can have potential as treatment for allergic asthma.
This study had some limitations, such as we could not evaluate levels of other related cytokines and also other related inflammatory and allergic biofactors. Moreover, we could not study the effect of thymol on the chronic form of allergic asthma.
Ethics Approval and Consent to Participate
This research was approved by Xi’an International Medical Center Hospital animal ethical committee, Approved No. GJYX-KY-2023-014.
Consent for Publication
Not Applicable.
Availability of Data and Materials
Data are available on request from the corresponding author.
Conflict of Interest
There is no conflict of interest.
Funding
The study was supported by Xi’an International Medical Center Hospital with grant No: 2022QN09.
Authors’ contributions
LZ, WZ, YW, PC, CL, YS, SSA, and AL participated in the planning, study, testing, analysis of data, and writing of the manuscript.
REFERENCES
1. Wenzel, SE. Asthma: Defining of the persistent adult phenotypes. Lancet. 2006;368:804–813. 10.1016/S0140-6736(06)69290-8
2. Montuschi, P. Pharmacotherapy of patients with mild persistent asthma: Strategies and unresolved issues. Front Pharmacol. 2011; 2:35. 10.3389/fphar.2011.00035
3. Zhang, Y, Li, S, Huang, S, Cao, L, Liu, T, Zhao, J, et al. IL33/ST2 contributes to airway remodeling via p-JNK MAPK/STAT3 signaling pathway in OVA-induced allergic airway inflammation in mice. Exp Lung Res. 2019; 45:65–75. 10.1080/01902148.2019.1611972
4. Cantani, A. and Micera, M. A study on 300 asthmatic children, 300 controls and their parents confirms the genetic transmission of allergy and asthma. Eur Rev Med Pharmacol. SCI. 2011; 15:1051–1056.
5. Vale, K. Targeting the JAK-STAT pathway in the treatment of ‘Th2-high’ severe asthma. Future Med Chem. 2016; 8:405–419. 10.4155/fmc.16.4
6. Zhiwei Wu, Entezar Mehrabi Nasab, Poonam Arora, Seyyed Shamsadin Athari. Study effect of probiotics and prebiotics on treatment of OVA-LPS-induced of allergic asthma inflammation and pneumonia by regulating the TLR4/NF-kB signaling pathway. J Transl Med. 2022; 20:130. 10.1186/s12967-022-03337-3
7. Narendra VT, Anamika S, Robin J, Monika K, Vishal A. Vitex negundo linn extract alleviates inflammatory aggravation and lung injury by modulating AMPK/PI3K/Akt/p38-NF-κB and TGF-β/Smad/Bcl2/caspase/LC3 cascade and macrophages activation in murine model of OVA-LPS induced allergic asthma. J Ethnopharmacol. 2021; 271:113894. 10.1016/j.jep.2021.113894
8. Akbar Mohammadi, Soleiman Mahjoub, Kamran Ghafarzadegan, Hamid Reza Nouri. Immunomodulatory effects of thymol through modulation of redox status and trace element content in experimental model of asthma. Biomed Pharmacother. 2018; 105:856–861. 10.1016/j.biopha.2018.05.154
9. El-Sayed el SM, Mansour AM, Abdul-Hameed MS. Thymol and carvacrol prevent doxorubicin-induced cardiotoxicity by abrogation of oxidative stress, inflammation, and apoptosis in rats. J Biochem Mol Toxicol. 2016; 30(1):37–44. 10.1002/jbt.21740
10. Bahador Hajimohammadi, Seyyede Masoume Athari, Mohammad Abdollahi, Ghasem Vahedi, Seyyed Shamsadin Athari. Oral administration of acrylamide worsens the inflammatory responses in the airways of asthmatic mice through agitation of oxidative stress in the lungs. Front Immunol. 2020; 11:1940. 10.3389/fimmu.2020.01940
11. Masoume Athari S, Mehrabi Nasab E, Shamsadin Athari S. Study effect of ocimum basilicum seeds on mucus production and cytokine gene expression in allergic asthma mice model. Revue française d’allergologie. 2018; 58(7):489–493. 10.1016/j.reval.2018.08.003
12. Wenxin Wang, Zhaochuan Yang, Meixiang Li, Zhenhong Wang, Yanchun Shan, Zhenghai Qu. Six1 promotes epithelial-mesenchymal transition in bronchial epithelial cells via the TGFβ1/Smad signalling pathway. Int Arch Allergy Immunol. 2021; 182:479–488. 10.1159/000512873
13. Martin T Johnson, Cory Benson J, Trayambak Pathak, Ping Xin, Abagail S McKernan, Scott M. Emrich, et al. The airway smooth muscle sodium/calcium exchanger NCLX is critical for airway remodeling and hyperresponsiveness in asthma. J Biol Chem. 2022; 298(8):102259. 10.1016/j.jbc.2022.102259
14. Jun-xia Jiang, Yan Guan, Hui-juan Shen, Yong-liang Jia, Jian Shen, Lin-hui Zhang, et al. Inhibition of soluble epoxide hydrolase attenuates airway remodeling in a chronic asthma model. Eur J Pharmacol. 2020; 868:172874. 10.1016/j.ejphar.2019.172874
15. Rasha M Hussein, El-Shaimaa A Arafa, Sayed Abdel Raheem, Wafaa R Mohamed. Thymol protects against bleomycin-induced pulmonary fibrosis via abrogation of oxidative stress, inflammation, and modulation of miR-29a/TGF-β and PI3K/Akt signaling in mice. Life Sci. 2023; 314: 121256. 10.1016/j.lfs.2022.121256
16. Kexin Ma, Cailong Li, Jingbin Xu, Fei Ren, Xin Xu, Chaosheng Liu, et al. LncRNA Gm16410 regulates PM2.5-induced lung endothelial-mesenchymal transition via the TGF-β1/Smad3/p-Smad3 pathway. Ecotoxicol Environ Saf. 2020; 205:111327. 10.1016/j.ecoenv.2020.111327
17. Ying Zhou, Qirui Duan, Dong Yang. In vitro human cell-based models to study airway remodeling in asthma. Biomed Pharmacother. 2023; 159:114218. 10.1016/j.biopha.2023.114218
18. Feifei Chen, Kenji Ogawa, Xubao Liu, Teresa M Stringfield, Yan Chen. Repression of Smad2 and Smad3 transactivating activity by association with a novel splice variant of CCAAT-binding factor C subunit. Biochem J. 2002; 364:571–577. 10.1042/bj20011703
19. Ji Young Kang, Chin Kook Rhee, Ju Sang Kim, Chan Kwon Park, Seung Joon Kim, Sang Haak Lee, et al. Effect of tiotropium bromide on airway remodeling in a chronic asthma model. Ann Allergy Asthma Immunol. 2012; 109:29–35. 10.1016/j.anai.2012.05.005
20. Talha Ijaz, Konrad Pazdrak, Mridul Kalita, Rolf Konig, Sanjeev Choudhary, Bing Tian, et al. Systems biology approaches to understanding epithelial mesenchymal transition (EMT) in mucosal remodeling and signaling in asthma. World Allergy Organ J. 2014; 7:13. 10.1186/1939-4551-7-13
21. Daniel J. Bernard. Both SMAD2 and SMAD3 mediate activin-stimulated expression of the follicle-stimulating hormone—Subunit in mouse gonadotrope cells. Mol Endocrinol. 2004; 18(3):606–623. 10.1210/me.2003-0264
22. Bin Sun, Huiqiao Wu, Jiajia Lu, Rongcheng Zhang, Xiaolong Shen, Yifei Gu, et al. Irisin reduces bone fracture by facilitating osteogenesis and antagonizing TGF-β/Smad signaling in a growing mouse model of osteogenesis imperfecta. J Orthop Translat. 2023; 38:175–189. 10.1016/j.jot.2022.10.012
23. Rie Watanabe, Yuichiro Yamada, Yu Ihara, Yoshimichi Someya, Akira Kubota, Shinji Kagimoto, et al. The MH1 domains of Smad2 and Smad3 are involved in the regulation of the ALK7 signals. Biochem Biophys Res Commun. 1999; 254:707–712. 10.1006/bbrc.1998.0118
24. Taro Umezu, Satoshi Nakamura, Yuiko Sato, Tami Kobayashi, Eri Ito, Takaya Abe, et al. Smad2 and Smad3 expressed in skeletal muscle promote immobilization-induced bone atrophy in mice. Biochem Biophys Res Commun. 2021; 582:111e117. 10.1016/j.bbrc.2021.10.043
25. Moir LM, Burgess JK, Black JL. Transforming growth factor beta 1 increases fibronectin deposition through integrin receptor alpha 5 beta 1 on human airway smooth muscle. J Allergy Clin Immunol. 2008; 121:1034–1039. 10.1016/j.jaci.2007.12.1159
26. Ershun Zhou, Yunhe Fu, Zhengkai Wei, Yuqiang Yu, Xichen Zhang, Zhengtao Yang. Thymol attenuates allergic airway inflammation in ovalbumin (OVA)-induced mouse asthma. Fitoterapia. 2014; 96:131–137. 10.1016/j.fitote.2014.04.016
27. Qiongyao Zeng, Yuncheng Che, Yu Zhang, Mei Chen, Qiang Guo, Wenjing Zhang. Thymol isolated from thymus vulgaris L. inhibits colorectal cancer cell growth and metastasis by suppressing the Wnt/β-Catenin pathway. Drug Des Devel Ther. 2020:14 2535–2547. 10.2147/DDDT.S254218
28. Lan Yao, Guo Hou, Lu Wang, Xiao-shu Zuo, Zhou Liu. Protective effects of thymol on LPS-induced acute lung injury in mice. Microb Pathog. 2018: 116:8–12. 10.1016/j.micpath.2017.12.065
29. Minmin Huang, Entezar Mehrabi Nasab, Seyyed Shamsadin Athari. Immunoregulatory effect of mesenchymal stem cell via mitochondria signaling pathways in allergic asthma. Saudi J Biol Sci. 2021; 28:6957–6962. 10.1016/j.sjbs.2021.07.071
30. Wang, Z, DiDonato, JA, Buffa, J, Comhair, SA, Aronica, MA, Dweik, RA, et al. Eosinophil peroxidase catalyzed protein carbamylation participates in asthma, J Biol Chem. 2016; 291(42):22118–22135. 10.1074/jbc.M116.750034
31. El-Sayed, EM, Abd-Allah, AR, Mansour, AM, El-Arabey, AA. Thymol and carvacrol prevent cisplatin-induced nephrotoxicity by abrogation of oxidative stress, inflammation, and apoptosis in rats. J Biochem Mol Toxicol. 2015; 29(4):165–172. 10.1002/jbt.21681