
Download
REVIEW ARTICLE
Unveiling the hidden power of noncoding RNAs in pediatric respiratory diseases
Shishu Yu, Lili Chen, Mingyao Zhang, Yu Lu*
Department of Pediatrics, The Third Affiliated Hospital of Changchun University of Chinese Medicine, Changchun, Jilin, China
Abstract
Respiratory diseases in children are common health problems that significantly impact their quality of life and health status, and this has its own unique challenges compared to adults. A growing body of research has focused on epigenetic mechanisms that relate with the development of various diseases, such as pediatric respiratory diseases. Noncoding RNAs (ncRNAs), especially long noncoding RNAs, microRNA, and circular RNA, are reported to play a regulatory role in pediatric respiratory diseases whose mutations or aberrant expressions are strongly associated with the development of these diseases. In this review, we mainly discussed the functions of these three ncRNAs in pediatric respiratory diseases.
Key words: biomarker, noncoding RNAs, pediatric respiratory diseases, respiratory system
*Corresponding author: Yu Lu, Department of Pediatrics, The Third Affiliated Hospital of Changchun University of Chinese Medicine, Changchun, Jilin 130117, China. Email address: [email protected]
Received 15 May 2024; Accepted 20 September 2024; Available online 1 November 2024
Copyright: Yu S, et al.
License: This open access article is licensed under Creative Commons Attribution 4.0 International (CC BY 4.0). http://creativecommons.org/licenses/by/4.0/
Introduction
Many ribonucleic acids (RNAs) are present in cells that do not encode proteins but play important regulatory roles. In fact, these noncoding RNAs (ncRNAs) occupy a very large percentage of eukaryotic transcriptome and include a variety of classes.1 Increasing evidence has proved that ncRNAs are widely involved in growth, differentiation, development, immunity, and even have important regulatory roles in tumor formation.2–4 The rapid development of high-throughput sequencing technologies and computational platforms has accelerated the discovery of ncRNAs. A variety of ncRNAs have been discovered, and, based on their roles in biology, are categorized into housekeeping ncRNAs and regulatory ncRNAs.5 They have a role in gene regulation at different levels, including messenger RNA (mRNA) processing, transcription, translation, and chromatin modification Figure 1).6
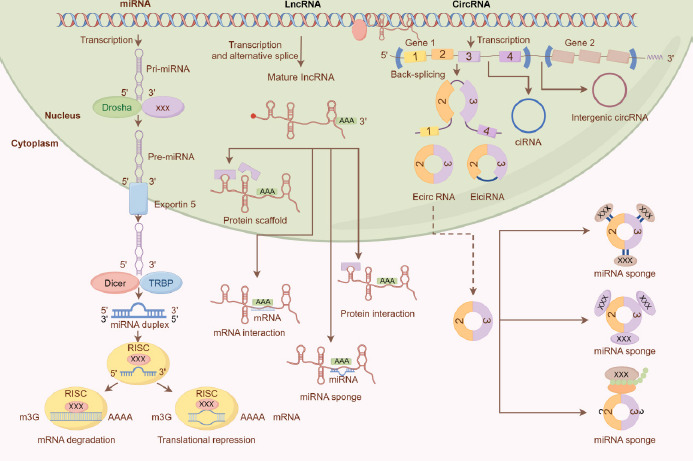
Figure 1 Functions of several ncRNAs. MiRNAs perform their biological functions by participating in the regulation of the translation of their downstream genes. LncRNAs are involved extensively in a variety of important regulatory processes, such as chromosome silencing, genomic imprinting, chromatin modification, transcriptional activation, transcriptional interference, and intranuclear transport. Biological functions of circRNAs include acting as miRNA sponges, regulating transcription, and facilitating protein–protein interactions. (This figure was created in Figdraw.)
Respiratory diseases, the most common diseases in children, are a major risk to their lives and health.7 More than 6.6 million deaths of children aged <5 years are reported annually, mainly caused by respiratory diseases.8 A global survey estimates that 14% of children suffer from asthma, and its prevalence is increasing.9 Approximately 13,000 and 14,000 live births are affected by cystic fibrosis (CF) every year globally, which is the second most common chronic respiratory disease in children.10
Many pediatric respiratory diseases remain incurable, contributing to the growing burden of noncommunicable respiratory diseases among adults.8 We have chosen three common pediatric respiratory diseases: childhood asthma, CF, and respiratory syncytial virus (RSV) infection. In this review, we summarize the existing research on ncRNAs as pathogenic factors and biomarkers of these diseases.
Noncoding RNAs and childhood asthma
Asthma is a chronic airway disease characterized by chronic airway inflammation.11 The clinical manifestations are recurrent wheezing, shortness of breath, chest tightness or cough, which often occurs or worsens at night or in early morning. Asthma being a common respiratory disease endangers the health of children. Its incidence is high, often manifested as a repeated chronic disease course, seriously affecting not only the learning, life, and activities but also the growth and development of children.12 In many case child asthma eventually develops into adult asthma because of delayed or improper treatment; lung function is impaired and some children even lose physical activity completely. Acute attacks of asthma can be fatal if not treated promptly and effectively. Although prevalence of asthma in children has increased over decades, no definitive explanation is known for this trend.13 Despite the fact that clinical manifestations of asthma alleviates in most children with effective asthma control by avoiding triggering factors, managing the disease rationally, and taking medications, no full cure is available to date. Furthermore, diagnosing asthma in children is difficult, since many childhood conditions exhibit symptoms similar to asthma, such as shortness of breath, wheezing, and coughing. Above all, there is a need to explore the pathogenesis and treatment of childhood asthma from a new perspective. In the face of these challenges, the discovery of long noncoding RNAs (lncRNAs), microRNAs (miRNAs), and circular RNAs (cirRNAs) has provided new opportunities to understand the pathogenesis and treatment of childhood asthma.
MicroRNAs and childhood asthma
MicroRNA is a small noncoding RNA that binds to the 3’ untranslated region (3’ UTR) of target genes to promote degradation or inhibition of the translation of mRNA.3 The processing and maturation of miRNAs is a multistep sophisticated reaction that undergoes spatial transition from the nucleus to the cytoplasm and is coordinated by various enzymes and auxiliary proteins, regulated at multiple levels.14 Since microarrays and new-generation sequencing techniques have been developed in the past few decades, it is possible to determine the expression and sequence of many miRNAs simultaneously. Kho et al. collected 153 children serum samples that were profiled for 754 miRNAs. MiR-146b, miR-206, and miR-720, involved in nuclear factor kappa B (NF-kβ) and glycogen synthase kinase 3–protein kinase B (GSK3/AKT) signaling pathways, were found to have excellent predictive power with childhood asthma exacerbation.15 Small-RNA (sRNA) sequencing was performed with whole blood samples of 374 asthmatic children (aged 6–14 years), and miR-451b, miR-7-5p, miR-532-3p, miR-296-5p, and miR-766-3p were related to childhood asthma.16 Analysis of plasma metabolite principal components from asthmatic children discovered miR-143-3p to be associated with metabolites and lung function trajectories in childhood asthma.17
MicroRNAs are implicated in asthma pathogenesis by the regulation of inflammatory reactions. According to miRNA profiling, levels of let-7e miRNA, miR-98, and miR-497 were more than two-fold higher in moderate-to-severe asthma patients than in controls. In addition, let-7a miRNA regulated interleukin 13 (IL-13), a cytokine critical for allergic lung disease expression, and let-7 miRNA inhibition suppressed allergic cytokine production and phenotype of asthma.18 Evidence was discovered that miR-21 levels were elevated in the serum of asthmatic patients, suggesting that it could be a biomarker of asthma.19 By utilizing transgenic mice that overexpressed IL-13, researchers identified that miR-21 expressed differently in animals with induced inflammation, compared to those averting induced allergic inflammation.
Airway remodeling is influenced by miR-133a, which negatively regulates the small G protein RhoA expression and contraction of bronchial smooth muscle cells.20 Furthermore, miR-26a promotes the hypertrophy of human airway smooth muscle cells.21 According to the next-generation sequencing, miR-10a was identified as the highest abundance miRNA accounting for over 20% of the sRNA reads from primary human airway smooth muscle cells, and regulated airway remodeling by reducing proliferation of cells.22 Let-7i-5p, whose level was related to particulate matter 2.5 (PM2.5) exposure in asthmatic children, was obviously overexpressed in asthmatic plasma, and mechanistically extracellular vesicles-packaged let-7i-5p could mediate remodeling during PM2.5-induced asthma attacks.23
Childhood asthma could be controlled and prevented with inhaled corticosteroids (ICS), and miR-155-5p and miR-532-5p could be the pharmacogenomic predictors of their response.24 Another study found that a poor response to ICS treatment was associated with immune dysregulation caused by miR-339-3p.25
Circular RNAs and childhood asthma
CircRNAs regulate transcription and splicing processes through specific interactions with DNA, RNA, and proteins via subcellular localization, modulate cytoplasmic mRNA stability and translation, interfere with signaling pathways, and serve as templates for translation in diverse biological and pathophysiological settings.26,27 The emerging applications of circRNAs in interfering with cellular processes, modulating immune responses, and directly translating proteins are opening up new ideas for biomedical research.28–30 Emerging applications are providing new ideas for pediatric asthma research.
Liang et al. found that circS100A11, which is predominantly expressed in monocytes, is significantly upregulated in children with asthma by microarray analysis of circRNAs sequencing profile. CircS100A11 can promote S100A11 gene translation by competitively binding to cell cycle associated protein 1 (CAPRIN1) coding gene to reduce the inhibition of host gene S100A11 translation by CAPRIN1. S100A11 then releases SP3 from nucleolin and promotes SP3 binding to the signal transducer and activator of transcription 6 (STAT6) promoter, thereby enhancing STAT6 expression and M2a macrophage activation. Meanwhile, specific knockdown of S100A11 in macrophages attenuates lung inflammation in the murine asthma model.31 Circ_0000029 has been proved as a potential target for childhood asthma treatment because it regulates KCNA1 gene expression by targeting miR-576-5p, which finally inhibits the abnormal migration and proliferation of airway smooth muscle cells.32 Currently, the study of circRNAs for asthma is focused mainly on phenotypes of adult patients. Therefore, more studies are needed to screen significantly altered circRNAs using childhood asthma samples or by using computational algorithms (such as deep learning models) to offer more potential candidate circRNAs, and explore the mechanism of action in conjunction with molecular biology experiments, in order to search for potential diagnostic and therapeutic targets of childhood asthma.33 Most importantly, more attention is required to the experimental limitations of the study for the participation of these circRNAs in human pathologies, including echnological and ethical and methodological challenges for obtaining biological samples in humans.34
Long noncoding RNAs and childhood asthma
Long noncoding RNAs are a class of RNA molecules that are more than 200 nucleotides (nt) in length and do not encode proteins. It has been found that lncRNAs have a conserved secondary structure, can interact with proteins, DNA, and RNA, and are involved in the regulation of genomic imprinting, chromatin modification, transcriptional activation or repression, transcriptional interference, and intranuclear transport, which are closely related to the occurrence and development of various diseases.35–37 The pathogenesis of asthma is influenced by gene signaling pathways, and lncRNA can regulate transcription and mRNA stability by various gene targets, contributing to airway remodeling and therapy-resistance.
A systematic transcriptome analysis of 10 asthmatic children prior to and after desensitization treatment showed that long intergenic non-protein coding RNA 2145 (LINC02145) and GUSB pseudogene 2 (GUSBP2) were identified as implicated in childhood asthma by involving in immune and inflammatory response.38 LncRNA CASC2 was shown to discriminate between healthy individuals and asthmatic children by quantitative real-time polymerase chain reaction (qRT-PCR) detection of mRNA levels in the serum of healthy and asthmatic children. In platelet-derived growth factor subunit B (PDGF-BB)-induced cell models, by sponging miR-31-5p, CASC2 inhibited cell proliferation, migration, and inflammation.39 Besides, lncRNA CDKN2B-AS1 was found to be significantly elevated in the plasma of children with asthma, and silencing CDKN2B-AS1 enhanced cell viability by inhibiting apoptosis and inflammation cytokine production in cell inflammation models. Consistently, in ovalbumin-induced mouse models, inflammation and hyper-responsiveness of the airways were alleviated by silencing CDKN2B-AS1.40 By detecting peripheral blood mononuclear cells and inflammatory cytokines in serum, lncRNA THRIL was proved to correlate with inflammatory cytokines and increase the risk and severity of childhood asthma.41
Dai et al. found that asthmatic children had high peripheral blood levels of lncRNA PTTG3P. In 16HBE cells, PTTG3P knockdown could inhibit epithelial–mesenchymal transition, proliferation, and migration, and through targeting miRNA-192-3p/CCNB1, PTTG3P could promote childhood asthma progression.42 A significant increase in lnc-BAZ2B gene expression was associated with childhood asthma, while a decrease in lncRNA-PTPRE-AS1 expression was associated with macrophage polarization, which contributes to type II inflammation.43 Recently, Zheng et al. found that exosomal lncRNA PM2.5-associated exosomal transcript (PAET) participated in childhood asthma by increasing DNA damages regulating m6A RNA-dependent oxidative phosphorylation (OXPHOS).44
In conclusion, the above studies suggest that lncRNAs can be involved in childhood asthma by modulating immune response, airway inflammation, and regulation of cytokine expression. Unfortunately, the mechanism by which lncRNAs are regulated in childhood asthma is unclear.
Most importantly, these findings indicate that ncRNAs play a role in childhood asthma pathogenesis not only by mediating inflammation and airway remodeling but also by immune response (Table 1).
Table 1 ncRNAs in childhood asthma pathogenesis.
| Class of ncRNAs | Subject | Functions |
|---|---|---|
| MicroRNAs | let-7i-5p miR-143-3p miR-145-5p miR-106a-5p/18a-5p/144-3p/ 375/21 miR-146a-5p/210-3p miR-192 let-7a/miR-155 miR-451a miR-21 miR-451a |
Modulating the mitogen-activated protein kinase (MAPK) signaling pathway.23 Drivers of poor lung function trajectories.17 Increasing airway smooth muscle cell proliferation.45 Promising biomarker for diagnosis.46,47 Correlating with T regulatory cells frequency.48 Suppressing T follicular helper cell differentiation.49 Potential biomarkers for diagnosis and severity.50 Contributing to T helper 2 (Th2) cell differentiation.51 Regulating bronchial epithelial cell proliferation.52 Inhibiting airway remodeling by targets of cadherin 11(CDH11).53 |
| CircRNAs | circS100A11 circ_0005519 circ_0000029 |
Enhancing M2a macrophage activation and lung inflammation.31 Increasing IL-13/IL-6.54 Repressing the abnormal airway smooth muscle cells (ASMCs) migration.32 |
| LncRNAs | CASC2 CDKN2B-AS1 PAET LINC02145/GUSBP2 THRIL PTTG3P LINC01559/SNHG8 BAZ2B TUG1 |
Inhibiting ASMCs proliferation, migration, and inflammation.39 Inhibiting ZFP36 promoter methylation and increasing nuclear receptor 4A1 (NR4A1).40 Enhancing DNA damage.44 Involving in immune response and inflammatory response.38 Correlating with inflammatory cytokines.41 Promoting progression of childhood asthma.42 Correlating with progression of childhood asthma.55 Promoting M2 macrophage activation and inflammation.56 Promoting proliferation and migration.57 |
Further studies are needed to screen significantly altered NcRNAs using childhood asthma samples (such as peripheral blood, sputum, bronchoalveolar lavage fluid, and exhaled breath condensate) using the latest sequencing technology, and explore the mechanism of action in conjunction with molecular biology experiments, in order to search for potential diagnostic and therapeutic targets of childhood asthma.
Noncoding RNAs and Cystic Fibrosis
Cystic fibrosis is an inherited disease that primarily affects the gastrointestinal and respiratory systems. It is usually characterized by chronic obstructive lung lesions, pancreatic exocrine dysfunction, and abnormally elevated sweat electrolytes.58 CF is primarily caused by mutations in the cystic fibrosis transmembrane receptor (CFTR) gene, and the lung disease is significantly affected by chronic inflammation and infection, both being secondary events contributing to CF.59
MicroRNAs and cystic fibrosis
The role of miRNAs in regulating the expression of the CFTR gene is thoroughly studied in numerous studies. Gillen et al. reported that miR-145 and miR-494 directly target discrete sites in the 3’UTR of the CFTR gene to regulate its expression.60 The bronchial brushings of CF patients showed higher levels of miR-145, miR-223, and miR-494 than non-CF brushings, illustrating the complexity of CFTR post-transcriptional regulation.61,62 In vitro, miR-101 and miR-494 regulated the CTFR gene expression, whereas miR-138 inhibited SIN3A, causing increase in the CFTR gene expression.63,64 Overexpression of MiR-138 enhanced CFTR molecule binding to the surface of cells, which is a potential therapeutic target of the CFTR gene.64 Oligonucleotide sequences specifically target and inhibit miR-101, miR-145, and miR-509-3p, successfully rescuing CFTR protein levels.65 In primary lung epithelial cells from CF patients, miR-16 restores functional expression of the cyclic AMP-activated apical chloride channel, F508del-CFTR.66 Since miRNAs act upstream of correctors and potentiators, miRNA-based therapies enhancing the CFTR gene expression may increase the effectiveness of the current CFTR modulators being tested in CF patients.
It is essential to carry out further research in order to develop biocompatible materials that are capable of delivering therapeutics efficiently at local sites because of barriers in the lungs with CF.
MicroRNAs may also control inflammatory responses in CF, particularly during pulmonary exacerbations, by affecting gene expression during inflammation. MiRNA-126 was the first described miRNA in CF; it decreased in airway epithelial cells regulating the TOM1 gene expression, involved in the ubiquitinated proteins trafficking and negative regulation of toll-like receptor 2 (TLR2), TLR4, and IL-1R1 pathways.67 In CF lung epithelial cells, increased miR-155 reduced SHIP1 expression, thereby triggering phosphatidylinositol 3-kinase–protein kinase B (PI3K/Akt) signaling and thereby promoting proinflammatory expression of IL-8.68 In another study, compared with healthy groups, miR-145 and miR-494 were obviously increased in nasal epithelial tissues of CF patients.69 There was a decrease in miR-17 levels in CF bronchial brushings and a decrease in miR-17 and miR-93 expressions in pseudomonas-induced bronchial epithelial cell lines, thus increasing the level of IL-8.70,71
Furthermore, macrophages from patients with CF are not capable of killing bacteria inside the cell. In CF cells, miR-181b is overexpressed, impairing some mechanisms of inflammation resolution dependent on N-formyl peptide receptor 2 (ALX/FPR2).72 In this regard, miR-181b targeting may contribute to the enhancement of anti-inflammatory and anti-microbial defenses in CF.
Long noncoding RNAs and cystic fibrosis
In a microarray analysis of lncRNAs, 1063 differentially expressed lncRNAs were detected in CF patients and non-CF patients, and several lncRNAs were divergently transcribed as gene promoters, suggesting a connection between lncRNAs and proteins.73 There are a number of reasons for this: (a) these heterogeneous findings can be attributed to a number of factors, such as age, gender, pulmonary function, microbial colonization, and use of medication; (b) because the sample size was small, it did not provide a high level of statistical power.
Saayman et al. found that lncRNA BGas modulated the local chromatin and DNA architecture of intron 11 of the CFTR gene in concert with HMGA1, HMGB1, and WIBG.74 Biotin-labeled oligonucleotides are used to identify the mechanism of how BGas modulates CFTR expression, as well as non-histone chromosomal proteins (HMG-14 and HMG-17), high mobility group protein B1 (HMGB1), and partner of Y14 and mago (WIBG), which are DNA-binding proteins that are capable of altering chromatin architecture to regulate transcription.75 In conclusion, BGas modulates CFTR expression by tethering chromatin architectural modifying proteins to intron 11 of the CFTR gene.
Noncoding RNAs and Respiratory Syncytial Virus Infection
Respiratory syncytial virus belongs to the family Paramyxoviridae, and is one of the major causes of lower respiratory tract infection in young children. A global estimate indicates 24.8 million cases of RSV and about 70–80K deaths reported annually, with 54% occurring in children aged ≤5 years.76 Clinical manifestations of RSV infection are related to age, underlying disease, environmental exposures, and history of previous respiratory infections. RSV leads to severe infections in high-risk children, with involvement of organs other than the respiratory system.77 NcRNAs are important for regulating gene expression and protein functions, and figuring out their role in RSV infection could reveal mechanism of the disease.
MicroRNAs and RSV infection
In a miRNA microarray of RSV-infected A549 cells, five miRNAs were induced and two miRNAs were repressed.78 The expression of miR-24, miR-29a, and miR-6087 in RSV-infected cell models was also demonstrated in other studies using qRT-PCR.76 Similarly, in human bronchial epithelial cell models, two miRNAs were upregulated and 24 downregulated when infected with RSV, among which miR-221 was obviously downregulated by RSV and inhibited nerve growth factor (NGF) and tropomyosin receptor kinase A (TrKA), finally increasing apoptosis and reducing viral replication and infectivity.79 Furthermore, according to a sequencing of exosomes from RSV-infected A549 cells, RSV-infected exosomes had an increased miRNA content.80
A study collected nasal mucosa cytology specimens of 19 healthy infants, 16 mild patients, 7 moderate patients, and 19 severe patients to perform deep sequencing. The result showed that compared with the healthy group, eight miRNAs were upregulated and three miRNAs were downregulated in the mild or severe group. One miRNA was upregulated in the severe group but downregulated in the mild group.81 Downregulation of miRNA-140-5p was observed in peripheral blood samples of children with bronchiolitis having RSV infection or healthy children. Further study showed that decreased miRNA-140-5p was related to RSV infection by targeting TLR4.82 A recent study identified miRNA targets in RSV-infected lung cells by using a biochemical method (CLEAR-CLIP) and found that miRNA-26 and miRNA-27 regulated cell cycle, metabolism, and antiviral immunity.83 Therefore, the altered miRNA profile is dramatically related to RSV infection, and RSV-regulated miRNAs are crucial for host responses to RSV infection.
Long noncoding RNAs and RSV infection
Long noncoding RNA was found to promote RSV replication by targeting miRNA miR-509-3p/Rab5c axis regulating vesicle transport.84 Tao et al. found that the lncRNA MEG3 expression was reduced when infected by RSV but the mRNA levels of TLR4, TNF-α, and IL-8 increased in patients’ samples and cell models, indicating MEG3 ameliorated RSV infection by suppressing TLR4 signaling.85
LncRNA n337374 is a promising target in RSV infection-induced asthma treatment because its overexpression could suppress dendritic cells maturation and relieve the symptoms of RSV-induced asthma by downregulating the CD86–ERK pathway.86
Circular RNAs and RSV infection
CircRNAs play a significant role in the interaction between cells and viruses. Yao et al. first characterized 53,719 cellular circRNAs and 2280 differentially expressed cellular circRNAs in RSC cell models, and profiled the general characteristics of both cellular and viral circRNAs.87 Their results revealed novel aspects of the host–RSV interaction, as well as circRNAs that could serve as novel therapeutic targets or biomarkers. It is currently believed that host circRNAs act as miRNA sponges, competing with other RNAs for miRNA binding sites, leading to indirect regulation of miRNA target genes and pathways.88 Future research into the functioning and mechanism of host circRNAs will enhance understanding of their roles in the onset and progression of viral diseases.89
Limitations and future outlook
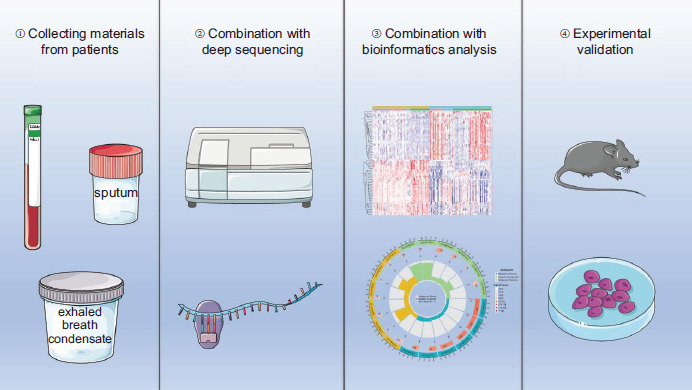
Limitations were observed to the use of ncRNA, such as how to locate nucleic acid drugs at specific targets or specific organs, and whether specific chemical modifications could be made to nucleic acid drugs because of natural mechanisms of degradation, as well as potential off-target effects. In addition, most of the evidences were based on animal and cell models, and these evidences must be carefully transposed to the case of human diseases. Thus, future research should pay more attention to human settings, and identify and validate more ncRNAs by collecting peripheral blood, sputum, and exhaled breath condensate from patients in combination with deep sequencing, bioinformatics analysis, and experimental validation (Figure 2).
Figure 2 The overall flowchart of ncRNA research.
Conclusions
The study of ncRNAs has a wide range of applications in the field of medicine, such as the treatment of relevant diseases or as markers of disease onset and progression by regulating the expression of specific ncRNAs.90–92 In pediatric respiratory diseases, miRNAs are studied most extensively, but lncRNA and circRNA also have potential for analysis. Furthermore, treatments based on synthetic ncRNAs or inhibitors could prove a breakthrough in treating pediatric respiratory diseases.
Funding
The authors declared that no funds, grants, or other support was received during the preparation of this manuscript.
Competing Interests
The authors had no relevant financial or nonfinancial interests to disclose.
Author Contributions
Shishu Yu and Lili Chen wrote the text of main manuscript and Mingyao Zhang collected the data. All authors reviewed the manuscript.
REFERENCES
1 Ding L, Wang R, Shen D, Cheng S, Wang H, Lu Z, et al. Role of noncoding RNA in drug resistance of prostate cancer. Cell Death Dis. 2021;12:590. 10.1038/s41419-021-03854-x
2 Cech TR, Steitz JA. The noncoding RNA revolution-trashing old rules to forge new ones. Cell. 2014;157:77–94. 10.1016/j.cell.2014.03.008
3 Zhang M, Han Y. MicroRNAs in chronic pediatric diseases (review). Exp Ther Med. 2024;27:100. 10.3892/etm.2024.12388
4 Safi A, Saberiyan M, Sanaei MJ, Adelian S, Davarani Asl F, Zeinaly M, et al. The role of noncoding RNAs in metabolic reprogramming of cancer cells. Cell Mol Biol Lett. 2023;28:37. 10.1186/s11658-023-00447-8
5 Zhang P, Wu W, Chen Q, Chen M. Noncoding RNAs and their integrated networks. J Integr Bioinform. 2019;16(3):20190027. 10.1515/jib-2019-0027
6 Dayal S, Chaubey D, Joshi DC, Ranmale S, Pillai B. Noncoding RNAs: Emerging regulators of behavioral complexity. Wiley Interdiscip Rev RNA. 2024;15:e1847. 10.1002/wrna.1847
7 Sonego M, Pellegrin MC, Becker G, Lazzerini M. Risk factors for mortality from acute lower respiratory infections (ALRI) in children under five years of age in low-and middle-income countries: A systematic review and meta-analysis of observational studies. PloS One. 2015;10:e0116380. 10.1371/journal.pone.0116380
8 Brennan LC, O’Sullivan A, MacLoughlin R. Cellular therapy for the treatment of paediatric respiratory disease. Int J Mol Sci. 2021;22(16):8906. 10.3390/ijms22168906
9 GBD Chronic Respiratory Disease Collaborators. Prevalence and attributable health burden of chronic respiratory diseases, 1990–2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet Resp Med. 2020;8:585–96. 10.1016/S2213-2600(20)30105-3
10 Scotet V, L’Hostis C, Férec C. The changing epidemiology of cystic fibrosis: Incidence, survival and impact of the CFTR gene discovery. Genes (Basel). 2020;11(6):589. 10.3390/genes11060589
11 Lambrecht BN, Hammad H. The immunology of asthma. Nat Immunol. 2015;16:45–56. 10.1038/ni.3049
12 Lin R, Guan R, Liu X, Zhao B, Guan J, Lu L. Significant rise of the prevalence and clinical features of childhood asthma in Qingdao China: Cluster sampling investigation of 10,082 children. BMC Public Health. 2014;14:1002. 10.1186/1471-2458-14-1002
13 Haktanir Abul M, Phipatanakul W. Severe asthma in children: Evaluation and management. Allergol Int. 2019;68:150–7. 10.1016/j.alit.2018.11.007
14 Lu TX, Rothenberg ME. MicroRNA. J Allergy Clin Immunol. 2018;141:1202–7. 10.1016/j.jaci.2017.08.034
15 Kho AT, McGeachie MJ, Moore KG, Sylvia JM, Weiss ST, Tantisira KG. Circulating microRNAs and prediction of asthma exacerbation in childhood asthma. Resp Res. 2018;19:128. 10.1186/s12931-018-0828-6
16 Tiwari A, Hobbs BD, Li J, Kho AT, Amr S, Celedón JC, et al. Blood miRNAs are linked to frequent asthma exacerbations in childhood asthma and adult COPD. Noncoding RNA. 2022;8(2):27. 10.3390/ncrna8020027
17 Mendez KM, Begum S, Tiwari A, Sharma R, Chen Q, Kelly RS, et al. Metabolite signatures associated with microRNA miR-143-3p serve as drivers of poor lung function trajectories in childhood asthma. EBioMedicine. 2024;102:105025. 10.1016/j.ebiom.2024.105025
18 Polikepahad S, Knight JM, Naghavi AO, Oplt T, Creighton CJ, Shaw C, et al. Proinflammatory role for LET-7 microRNAS in experimental asthma. J Biol Chem. 2010;285:30139–49. 10.1074/jbc.M110.14569810.1074/jbc.N110.145698
19 Sawant DV, Yao W, Wright Z, Sawyers C, Tepper RS, Gupta SK, et al. Serum MicroRNA-21 as a biomarker for allergic inflammatory disease in children. MicroRNA (Shariqah, UAE). 2015;4:36–40. 10.2174/2211536604666150220232507
20 Chiba Y. Noncoding RNAs and bronchial smooth muscle hyperresponsiveness in allergic bronchial asthma. Nihon Yakurigaku Zasshi Folia Pharmacol Japonica. 2020;155:364–8. 10.1254/fpj.20053
21 Mohamed JS, Lopez MA, Boriek AM. Mechanical stretch up-regulates microRNA-26a and induces human airway smooth muscle hypertrophy by suppressing glycogen synthase kinase-3β. J Biol Chem. 2010;285:29336–47. 10.1074/jbc.M110.101147
22 Hu R, Pan W, Fedulov AV, Jester W, Jones MR, Weiss ST, et al. MicroRNA-10a controls airway smooth muscle cell proliferation via direct targeting of the PI3 kinase pathway. FASEB J. 2014;28:2347–57. 10.1096/fj.13-247247
23 Zheng R, Du M, Tian M, Zhu Z, Wei C, Chu H, et al. Fine particulate matter induces childhood asthma attacks via extracellular vesicle-packaged Let-7i-5p-mediated modulation of the MAPK signaling pathway. Adv Sci (Weinheim, Baden-Wurttemberg, Germany). 2022;9:e2102460. 10.1002/advs.202102460
24 Li J, Panganiban R, Kho AT, McGeachie MJ, Farnam L, Chase RP, et al. Circulating microRNAs and treatment response in childhood asthma. Am J Resp Crit Care Med. 2020;202:65–72. 10.1164/rccm.201907-1454OC
25 Sharma R, Tiwari A, Kho AT, Celedón JC, Weiss ST, Tantisira KG, et al. Systems genomics reveals microRNA regulation of ICS response in childhood asthma. Cells. 2023;12(11):1505. 10.3390/cells12111505
26 Kristensen LS, Andersen MS, Stagsted LVW, Ebbesen KK, Hansen TB, Kjems J. The biogenesis, biology and characterization of circular RNAs. Nat Rev Genet. 2019;20:675–91. 10.1038/s41576-019-0158-7
27 Patop IL, Wüst S, Kadener S. Past, present, and future of circRNAs. EMBO J. 2019;38:e100836. 10.15252/embj.2018100836
28 Xue C, Li G, Zheng Q, Gu X, Bao Z, Lu J, et al. The functional roles of the circRNA/Wnt axis in cancer. Mol Cancer. 2022;21:108. 10.1186/s12943-022-01582-0
29 Chen W, Xu J, Wu Y, Liang B, Yan M, Sun C, et al. The potential role and mechanism of circRNA/miRNA axis in cholesterol synthesis. Int J Biol Sci. 2023;19:2879–96. 10.7150/ijbs.84994
30 Zhang W, He Y, Zhang Y. CircRNA in ocular neovascular diseases: Fundamental mechanism and clinical potential. Pharmacol Res. 2023;197:106946. 10.1016/j.phrs.2023.106946
31 Liang Q, Fu J, Wang X, Liu L, Xiao W, Gao Y, et al. circS100A11 enhances M2a macrophage activation and lung inflammation in children with asthma. Allergy. 2023;78:1459–72. 10.1111/all.15515
32 Wang R, Tan Y, Bao X, Xiong S, Liang R, Cai M, et al. Circ_0000029 interacts with the miR-576-5p/KCNA1 axis to hamper the development of pediatric asthma in an asthma-like in vitro assessment. Ann Clin Lab Sci. 2023;53:200–11.
33 Chen Y, Wang J, Wang C, Liu M, Zou Q. Deep learning models for disease-associated circRNA prediction: A review. Brief Bioinform. 2022;23(6):bbac364. 10.1093/bib/bbac364
34 Breuer J, Rossbach O. Production and purification of artificial circular RNA sponges for application in molecular biology and medicine. Methods Protoc. 2020;3(2):42. 10.3390/mps3020042
35 Wu T, Du Y. LncRNAs: From basic research to medical application. Int J Biol Sci. 2017;13:295–307. 10.7150/ijbs.16968
36 Ebrahimi N, Parkhideh S, Samizade S, Esfahani AN, Samsami S, Yazdani E, et al. Crosstalk between lncRNAs in the apoptotic pathway and therapeutic targets in cancer. Cytokine Growth Factor Rev. 2022;65:61–74. 10.1016/j.cytogfr.2022.04.003
37 Kopp F, Mendell JT. Functional classification and experimental dissection of long noncoding RNAs. Cell. 2018;172:393–407. 10.1016/j.cell.2018.01.011
38 Zheng P, Huang C, Leng D, Sun B, Zhang XD. Transcriptome analysis of peripheral whole blood identifies crucial lncRNAs implicated in childhood asthma. BMC Med Genom. 2020;13:136. 10.1186/s12920-020-00785-y
39 Yang Y, Sun Z, Ren T, Lei W. Differential expression of lncRNA CASC2 in the serum of childhood asthma and its role in airway smooth muscle cells proliferation and migration. J Asthma Allergy. 2022;15:197–207. 10.2147/JAA.S337236
40 Chen Z, Fan N, Shen G, Yang J. Silencing lncRNA CDKN2B-AS1 alleviates childhood asthma progression through inhibiting ZFP36 promoter methylation and promoting NR4A1 expression. Inflammation. 2023;46:700–17. 10.1007/s10753-022-01766-2
41 Wang X, Li W, Sun S, An H. Inter-correlation of lncRNA THRIL with microRNA-34a and microRNA-125b and their relationship with childhood asthma risk, severity, and inflammation. Allergol Immunopathol. 2023;51(1):187–94. 10.15586/aei.v51i1.736
42 Dai B, Sun F, Cai X, Li C, Liu F, Shang Y. Long noncoding RNA PTTG3P/miR-192-3p/CCNB1 axis is a potential biomarker of childhood asthma. Int Immunopharmacol. 2021;101:108229. 10.1016/j.intimp.2021.108229
43 Zeng H, Wang Y, Gu Y, Wang J, Zhang H, Gao H, et al. Polydatin attenuates reactive oxygen species-induced airway remodeling by promoting Nrf2-mediated antioxidant signaling in asthma mouse model. Life Sci. 2019;218:25–30. 10.1016/j.lfs.2018.08.013
44 Zheng R, Gao F, Xiao Y, Liang J, Mao Z, Gan C, et al. PM(2.5)-derived exosomal long noncoding RNA PAET participates in childhood asthma by enhancing DNA damage via m(6)A-dependent OXPHOS regulation. Environ Int. 2024;183:108386. 10.1016/j.envint.2023.108386
45 Tiwari A, Li J, Kho AT, Sun M, Lu Q, Weiss ST, et al. COPD-associated miR-145-5p is downregulated in early-decline FEV(1) trajectories in childhood asthma. J Allergy Clin Immunol. 2021;147:2181–90. 10.1016/j.jaci.2020.11.048
46 He L, Liu J, Wang X, Wang Y, Zhu J, Kang X. Identifying a novel serum microRNA biomarker panel for the diagnosis of childhood asthma. Exp Biol Med (Maywood, NJ). 2022;247:1732–40. 10.1177/15353702221114870
47 Elbehidy RM, Youssef DM, El-Shal AS, Shalaby SM, Sherbiny HS, Sherief LM, et al. MicroRNA-21 as a novel biomarker in diagnosis and response to therapy in asthmatic children. Mol Immunol. 2016;71:107–14. 10.1016/j.molimm.2015.12.015
48 Eldosoky MA, Hammad R, Rushdi A, Ibrahim HF, Tawfeik AM, Mora A, et al. MicroRNA-146a-5p and microRNA-210-3p correlate with T regulatory cells frequency and predict asthma severity in Egyptian pediatric population. J Asthma Allergy. 2023;16:107–21. 10.2147/JAA.S398494
49 Zhang D, Wu Y, Sun G. miR-192 suppresses T follicular helper cell differentiation by targeting CXCR5 in childhood asthma. Scand J Clin Lab Invest. 2018;78(3):236–42. 10.1080/00365513.2018.1440628
50 Karam RA, Abd Elrahman DM. Differential expression of miR-155 and Let-7a in the plasma of childhood asthma: Potential biomarkers for diagnosis and severity. Clin Biochem. 2019;68:30–6. 10.1016/j.clinbiochem.2019.04.007
51 Wang T, Zhou Q, Shang Y. Downregulation of miRNA-451a promotes the differentiation of CD4+ T cells towards Th2 cells by upregulating ETS1 in childhood asthma. J Innate Immun. 2021;13:38–48. 10.1159/000509714
52 Kang Y, Bai M, Deng L, Fan L, Wang X. MiRNA-21 regulates bronchial epithelial cell proliferation by activating Tgfβ1/Smad signaling pathway and its correlation with asthma severity in children. Iran J Public Health. 2021;50(10):1973–82. 10.18502/ijph.v50i10.7497
53 Wang T, Zhou Q, Shang Y. MiRNA-451a inhibits airway remodeling by targeting Cadherin 11 in an allergic asthma model of neonatal mice. Int Immunopharmacol. 2020;83:106440. 10.1016/j.intimp.2020.106440
54 Huang Z, Cao Y, Zhou M, Qi X, Fu B, Mou Y, et al. Hsa_circ_0005519 increases IL-13/IL-6 by regulating HSA-LET-7a-5p in CD4(+) T cells to affect asthma. Clin Exp Allergy. 2019;49:1116–27. 10.1111/cea.13445
55 Huang Z, Fu B, Qi X, Xu Y, Mou Y, Zhou M, et al. Diagnostic and therapeutic value of Hsa_circ_0002594 for T Helper 2-mediated allergic asthma. Int Arch Allergy Immunol. 2021;182:388–98. 10.1159/000511612
56 Xia L, Wang X, Liu L, Fu J, Xiao W, Liang Q, et al. lnc-BAZ2B promotes M2 macrophage activation and inflammation in children with asthma through stabilizing BAZ2B pre-mRNA. J Allergy Clin Immunol. 2021;147:921–32.e9. 10.1016/j.jaci.2020.06.034
57 Wang X, Chen J. Long noncoding RNA TUG1 promotes proliferation and migration in PDGF-BB-stimulated HASMCs by regulating miR-216a-3p/SMURF2 axis. BMC Mol Cell Biol. 2021;22:56. 10.1186/s12860-021-00396-0
58 Glasgow AMA, De Santi C, Greene CM. Noncoding RNA in cystic fibrosis. Biochem Soc Trans. 2018;46:619–30. 10.1042/BST20170469
59 Britto MT, Kotagal UR, Hornung RW, Atherton HD, Tsevat J, Wilmott RW. Impact of recent pulmonary exacerbations on quality of life in patients with cystic fibrosis. Chest. 2002;121:64–72. 10.1378/chest.121.1.64
60 Gillen AE, Gosalia N, Leir SH, Harris A. MicroRNA regulation of expression of the cystic fibrosis transmembrane conductance regulator gene. Biochem J. 2011;438:25–32. 10.1042/BJ20110672
61 Oglesby IK, Chotirmall SH, McElvaney NG, Greene CM. Regulation of cystic fibrosis transmembrane conductance regulator by microRNA-145,-223, and-494 is altered in ΔF508 cystic fibrosis airway epithelium. J Immunol (Baltimore, MD). 2013;190:3354–62. 10.4049/jimmunol.1202960
62 Ramachandran S, Karp PH, Osterhaus SR, Jiang P, Wohlford-Lenane C, Lennox KA, et al. Post-transcriptional regulation of cystic fibrosis transmembrane conductance regulator expression and function by microRNAs. Am J Resp Cell Mol Biol. 2013;49:544–51. 10.1165/rcmb.2012-0430OC
63 Megiorni F, Cialfi S, Dominici C, Quattrucci S, Pizzuti A. Synergistic post-transcriptional regulation of the cystic fibrosis transmembrane conductance regulator (CFTR) by miR-101 and miR-494 specific binding. PloS One. 2011;6:e26601. 10.1371/journal.pone.0026601
64 Ramachandran S, Karp PH, Jiang P, Ostedgaard LS, Walz AE, Fisher JT, et al. A microRNA network regulates expression and biosynthesis of wild-type and DeltaF508 mutant cystic fibrosis transmembrane conductance regulator. Proc Nat Acad Sci Am. 2012;109:13362–7. 10.1073/pnas.1210906109
65 Glasgow AMA, De Santi C, Greene CM. Noncoding RNA in cystic fibrosis. Biochem Soc Trans. 2018;46:619–30. 10.1042/BST20170469
66 Kumar P, Bhattacharyya S, Peters KW, Glover ML, Sen A, Cox RT, et al. miR-16 rescues F508del-CFTR function in native cystic fibrosis epithelial cells. Gene Ther. 2015;22:908–16. 10.1038/gt.2015.56
67 Oglesby IK, Bray IM, Chotirmall SH, Stallings RL, O’Neill SJ, McElvaney NG, et al. miR-126 is downregulated in cystic fibrosis airway epithelial cells and regulates TOM1 expression. J Immunol (Baltimore, MD). 2010;184:1702–9. 10.4049/jimmunol.0902669
68 Bhattacharyya S, Balakathiresan NS, Dalgard C, Gutti U, Armistead D, Jozwik C, et al. Elevated miR-155 promotes inflammation in cystic fibrosis by driving hyperexpression of interleukin-8. J Biol Chem. 2011;286:11604–15. 10.1074/jbc.M110.198390
69 Megiorni F, Cialfi S, Cimino G, De Biase RV, Dominici C, Quattrucci S, et al. Elevated levels of miR-145 correlate with SMAD3 down-regulation in cystic fibrosis patients. J Cyst Fibros. 2013;12(6):797–802. 10.1016/j.jcf.2013.03.007; 10.1016/S1569-1993(13)60124-1 Epub 2013 Apr 28
70 Fabbri E, Borgatti M, Montagner G, Bianchi N, Finotti A, Lampronti I, et al. Expression of microRNA-93 and interleukin-8 during pseudomonas aeruginosa-mediated induction of proinflammatory responses. Am J Resp Cell Mol Biol. 2014;50:1144–55. 10.1165/rcmb.2013-0160OC
71 Oglesby IK, Vencken SF, Agrawal R, Gaughan K, Molloy K, Higgins G, et al. miR-17 overexpression in cystic fibrosis airway epithelial cells decreases interleukin-8 production. Eur Resp J. 2015;46:1350–60. 10.1183/09031936.00163414
72 Pierdomenico AM, Patruno S, Codagnone M, Simiele F, Mari VC, Plebani R, et al. microRNA-181b is increased in cystic fibrosis cells and impairs lipoxin A(4) receptor-dependent mechanisms of inflammation resolution and antimicrobial defense. Sci Rep. 2017;7:13519. 10.1038/s41598-017-14055-y
73 McKiernan PJ, Molloy K, Cryan SA, McElvaney NG, Greene CM. Long noncoding RNA are aberrantly expressed in vivo in the cystic fibrosis bronchial epithelium. Int J Biochem Cell Biol. 2014;52:184–91. 10.1016/j.biocel.2014.02.022
74 Saayman SM, Ackley A, Burdach J, Clemson M, Gruenert DC, Tachikawa K, et al. Long noncoding RNA BGas regulates the cystic fibrosis transmembrane conductance regulator. Mol Ther. 2016;24:1351–7. 10.1038/mt.2016.112
75 Klune JR, Dhupar R, Cardinal J, Billiar TR, Tsung A. HMGB1: Endogenous danger signaling. Mol Med (Cambridge, MA). 2008;14:476–84. 10.2119/2008-00034.Klune
76 Wu W, Choi EJ, Lee I, Lee YS, Bao X. Noncoding RNAs and their role in respiratory syncytial virus (RSV) and human metapneumovirus (hMPV) infections. Viruses. 2020;12(3):345. 10.3390/v12030345
77 Esposito S, Abu Raya B, Baraldi E, Flanagan K, Martinon Torres F, Tsolia M, et al. RSV prevention in all infants: Which is the most preferable strategy? Front Immunol. 2022;13:880368. 10.3389/fimmu.2022.880368
78 Bakre A, Mitchell P, Coleman JK, Jones LP, Saavedra G, Teng M, et al. Respiratory syncytial virus modifies microRNAs regulating host genes that affect virus replication. J Gen Virol. 2012;93:2346–56. 10.1099/vir.0.044255-0
79 Othumpangat S, Walton C, Piedimonte G. MicroRNA-221 modulates RSV replication in human bronchial epithelium by targeting NGF expression. PloS One. 2012;7:e30030. 10.1371/journal.pone.0030030
80 Chahar HS, Corsello T, Kudlicki AS, Komaravelli N, Casola A. Respiratory syncytial virus infection changes cargo composition of exosome released from airway epithelial cells. Sci Rep. 2018;8:387. 10.1038/s41598-017-18672-5
81 Inchley CS, Sonerud T, Fjærli HO, Nakstad B. Nasal mucosal microRNA expression in children with respiratory syncytial virus infection. BMC Infect Dis. 2015;15:150. 10.1186/s12879-015-0878-z
82 Zhang Y, Shao L. Decreased microRNA-140-5p contributes to respiratory syncytial virus disease through targeting Toll-like receptor 4. Exp Ther Med. 2018;16:993–9. 10.3892/etm.2018.6272
83 Ressel S, Kumar S, Bermúdez-Barrientos JR, Gordon K, Lane J, Wu J, et al. RNA–RNA interactions between respiratory syncytial virus and miR-26 and miR-27 are associated with regulation of cell cycle and antiviral immunity. Nucleic Acids Res. 2024;52(9):4872–88. 10.1101/2023.06.05.543706
84 Li J, Li M, Wang X, Sun M, Ma C, Liang W, et al. Long noncoding RNA NRAV promotes respiratory syncytial virus replication by targeting the MicroRNA miR-509-3p/Rab5c Axis to regulate vesicle transportation. J Virol. 2020;94(10):e00113–20. 10.1128/JVI.00113-20.
85 Tao XW, Zeng LK, Wang HZ, Liu HC. LncRNA MEG3 ameliorates respiratory syncytial virus infection by suppressing TLR4 signaling. Mol Med Rep. 2018;17:4138–44. 10.3892/mmr.2017.8303
86 Sun S, Yao M, Yuan L, Qiao J. Long-chain noncoding RNA n337374 relieves symptoms of respiratory syncytial virus-induced asthma by inhibiting dendritic cell maturation via the CD86 and the ERK pathway. Allergol Immunopathol. 2021;49:100–7. 10.15586/aei.v49i3.85
87 Yao W, Pan J, Liu Z, Dong Z, Liang M, Xia S, et al. The cellular and viral circRNAome induced by respiratory syncytial virus infection. mBio. 2021;12:e0307521. 10.1128/mBio.03075-21
88 Wang S, Li X, Liu G, Qiu Z, Wang J, Yang D, et al. Advances in the understanding of circRNAs that influence viral replication in host cells. Med Microbiol Immunol. 2024;213:1. 10.1007/s00430-023-00784-7
89 Dremel SE, Tagawa T, Koparde VN, Hernandez-Perez C, Arbuckle JH, Kristie TM, et al. Interferon induced circRNAs escape herpesvirus host shutoff and suppress lytic infection. EMBO Rep. 2024;25:1541–69. 10.1038/s44319-023-00051-z
90 Song M, Wang H, Liu C, Jin S, Liu B, Sun W. Noncoding RNAs as regulators of the Hippo pathway in cardiac development and cardiovascular disease. Front Pharmacol. 2024;15:1348280. 10.3389/fphar.2024.1348280
91 Zhang Y, Zhan L, Jiang X, Tang X. Comprehensive review for noncoding RNAs: From mechanisms to therapeutic applications. Biochem Pharmacol. 2024;224:116218. 10.1016/j.bcp.2024.11621810.1016/j.jpba.2024.116218
92 Caporali A, Anwar M, Devaux Y, Katare R, Martelli F, Srivastava PK, et al. Noncoding RNAs as therapeutic targets and biomarkers in ischaemic heart disease. Nat Rev Cardiol. 2024;21(8):556–73. 10.1038/s41569-024-01001-5