
Download
ORIGINAL ARTICLE
Rhizoma coptidis can inhibit the excessive proliferation, inflammation, and transformation of lung fibroblasts into myofibroblasts
Jie Yanga, Yuting Huangb*, Zhimin Cuia, Chang Liua, Guofang Xiea
aDepartment of Geriatrics, ChaoHu Hospital of Anhui Medical University, Hefei City, Anhui Province, China
bDepartment of Radiotherapy, ChaoHu Hospital of Anhui Medical University, Hefei City, Anhui Province, China
Abstract
Background: Pulmonary fibrosis (PF) is a chronic, progressive, and irreversible heterogeneous disease of lung interstitial tissue. To combat progression of PF, new drugs are required to be developed. Rhizoma coptidis (COP), one of the main alkaloids of Coptis chinensis, is a traditional herbal medicine used to treat various inflammatory diseases.
Objective: To investigate the possible effects of Coptisine (Cop) on the growth, inflammation, as well as FMT of TNF-β1-induced HFL1 cells and uncover the mechanism.
Material and Methods: Human fetal lung fibroblast 1 (HFL1) was induced using 6ng/mL TGF-β1 as a model of pulmonary fibrosis. CCK-8, Brdu, and transwell assays indicated the effects on cell growth as well as motility. qPCR and the corresponding kits indicted the effects on cell inflammation. Immunoblot showed the effects on FMT and further confirmed the mechanism.
Results: Coptisine inhibits excessive growth as well as motility of TNF-β1-induced HFL1 cells. It further inhibits inflammation and ROS levels in TNF-β1-induced HFL1 cells. Coptisine inhibits the FMT process of TNF-β1-induced HFL1 cells. Mechanically, coptisine promotes the Nrf2/HO-1 pathway.
Conclusion: Coptisine can inhibit the excessive growth, inflammation as well as FMT of lung fibroblasts into myofibroblasts. It could serve as a promising drug of PF.
Key words: Rhizoma coptidis (COP), FMT, inflammation, Nrf2/HO-1 pathway, pulmonary fibrosis (PF)
*Corresponding author: Yuting Huang, Department of Radiotherapy, ChaoHu Hospital of Anhui Medical University, No. 64 Chaohu North Road, Chaohu City, Anhui Province, 238000, China. Email address: [email protected]
Received 18 April 2024; Accepted 6 May 2024; Available online 1 July 2024
Copyright: Yang J, et al.
License: This open access article is licensed under Creative Commons Attribution 4.0 International (CC BY 4.0). http://creativecommons.org/licenses/by/4.0/
Introduction
Pulmonary fibrosis (PF) is a chronic, progressive, and irreversible heterogeneous disease of lung interstitial tissue, characterized by persistent alveolar epithelial damage. In recent years, many studies have found that the occurrence of PF is related to the activation of fibroblast-to-myofibroblast transformation (FMT), abnormal tissue remodeling, immune response, and excessive deposition of extracellular matrix (ECM).1 In addition, the FMT process is an important pathway leading to ECM deposition, and inhibition of myofibroblast differentiation is shown to be an effective way to prevent PF.2 Multiple studies have found that levels of reactive oxygen species (ROS) are upregulated during transforming growth factor-beta (TGF-β1)-induced FMT, which is triggered by the secretion of inflammatory cytokines in PF. Studies on the pathogenesis of PF and treatment strategies have lagged behind medical advances and clinical requirement.3 The treatment of this disease remains a clinical challenge, and effective therapeutic drugs are still limited. Traditional Chinese medicine monomer compounds have various biological activities, with high safety. They play a wide range of roles in the treatment of diseases and are candidates for the prevention and treatment of PF. To combat progression of PF, new drugs are still required to be developed.
Rhizoma coptidis (COP), one of the main alkaloids of Coptis chinensis, is a traditional herbal medicine used to treat various inflammatory diseases.4 Rhizoma coptidis has anti-inflammatory, anticancer, and antibacterial effects and acts by regulating different signaling pathways.5 Rhizoma coptidis has been shown to have therapeutic effects on lung diseases. For example, Rhizoma coptidis alleviates septic lung damage by inhibiting lipopolysaccharide (LPS)-induced inflammation and apoptosis of lung epithelial cells.6 Rhizoma coptidis inhibits oxidative stress-induced cytotoxicity in V79-4 Chinese hamster lung fibroblasts by inducing nuclear factor erythroid 2 (Nrf2)-mediated heme oxygenase-1 (HO-1) expression.7 Rhizoma coptidis induces Gap 2 (G2) and mitosis (M) (G2/M) phase cell cycle arrest and ROS-dependent mitochondria-mediated apoptosis in A549 cells of non-small cell lung cancer.8 However, the role and mechanism of Rhizoma coptidis in PF remained unclear.
This study aimedto elucidate the effect of Rhizoma coptidis on PF and reveal its mechanism. Our results confirmed that Rhizoma coptidis inhibits hyperproliferation, inflammation, and myofibroblast transformation of lung fibroblasts through Nrf2–HO-1 signaling. Therefore, Rhizoma coptidis has the potential to serve as a promising drug to treat PF.
Materials and methods
Cell culture and treatment
Human HFL1 cell line, a fibroblast cell line isolated from the lung of a White, normal embryo, was obtained from iCell Bioscience Inc (China) and maintained in Dulbecco’s modified eagle medium (DMEM) with 10% fetal bovine serum (FBS) at 37°C with 5% CO2. Cells were stimulated with TGF-β1 (Sigma, St. Louis, MO, US) at a concentration of 6 ng/mL for 24 h. Cells were stimulated with Rhizoma coptidis (Sigma) at different concentrations of 0, 12.5, 25, and 50 μM for 24 h.
Cell viability
Cell viability was discovered by adding cell counting kit-8 (CCK-8) solution. After treatment with TGF-β1 or Rhizoma coptidis, CCK-8 agent was added to cells, following washing with PBS. Optical density (OD) in each well was quantified with a microplate reader at 450 nm.
5-Bromo-2’-deoxyuridine (BrdU) assay
Human HFL1 cells were incubated with BrdU agent (Abcam, Cambridge, UK) for 2 h. The agent was removed, and the cells were photographed by a fluorescence microscope (Zeiss, Jena, German).
Enzyme-linked immunosorbent serologic assay (ELISA)
After indicated stimulations, cell supernatants were subjected to ELISA to determine the level of tumor necrosis factor-α (TNF-α), interleukin 1β (IL-1β), and IL-6 following the manufacturer’s guidelines.
Transwell assay
Cell culture inserts were coated with 100-μL Matrigel (invasion, 1:3 diluted with serum-free media), and 105 cells were placed in the upper chamber. Invaded cells underside were fixed with 4% paraformaldehyde (PFA), stained with the 0.2% crystal violet, and imaged.
ROS detection
Cells were incubated with ROS kit (Sigma). The proportion of ROS positive nuclei in eight random fields (total nuclei: 500) was analyzed for each group.
Immunoblot assay
Proteins were extracted with radioimmunoprecipitation assay (RIPA) buffer (Beyotime, Shanghai, China). Then, the proteins were separated by 9% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE), and transferred onto polyvinylidene fluoride (PVDF) membranes, followed by blocking with 5% bovine serum albumin (BSA). Membranes were conjugated with primary antibodies against E-cadherin (1:1000, ab231303; Abcam), collagen I (1:1000, ab138492; Abcam), FN1 (1:500, ab2413; Abcam), alpha smooth muscle-actin (α-SMA) (1:500, ab7817; Abcam), Nrf2 (1:1000, ab62352; Abcam), HO-1 (1:500, ab305290; Abcam), and β-actin (1:3000, ab8226; Abcam) for 2 h. Subsequently, the membranes were incubated with secondary antibodies for 1 h. The blots were analyzed with enhanced chemiluminescence (ECL) kit.
Statistical analysis
Student’s t-test was used to determine statistical significance. P < 0.05 was considered as statistically significant. Error bars represent standard deviation.
Results
Rhizoma coptidis inhibits excessive growth as well as motility of TGF-β1-induced HFL1 cells
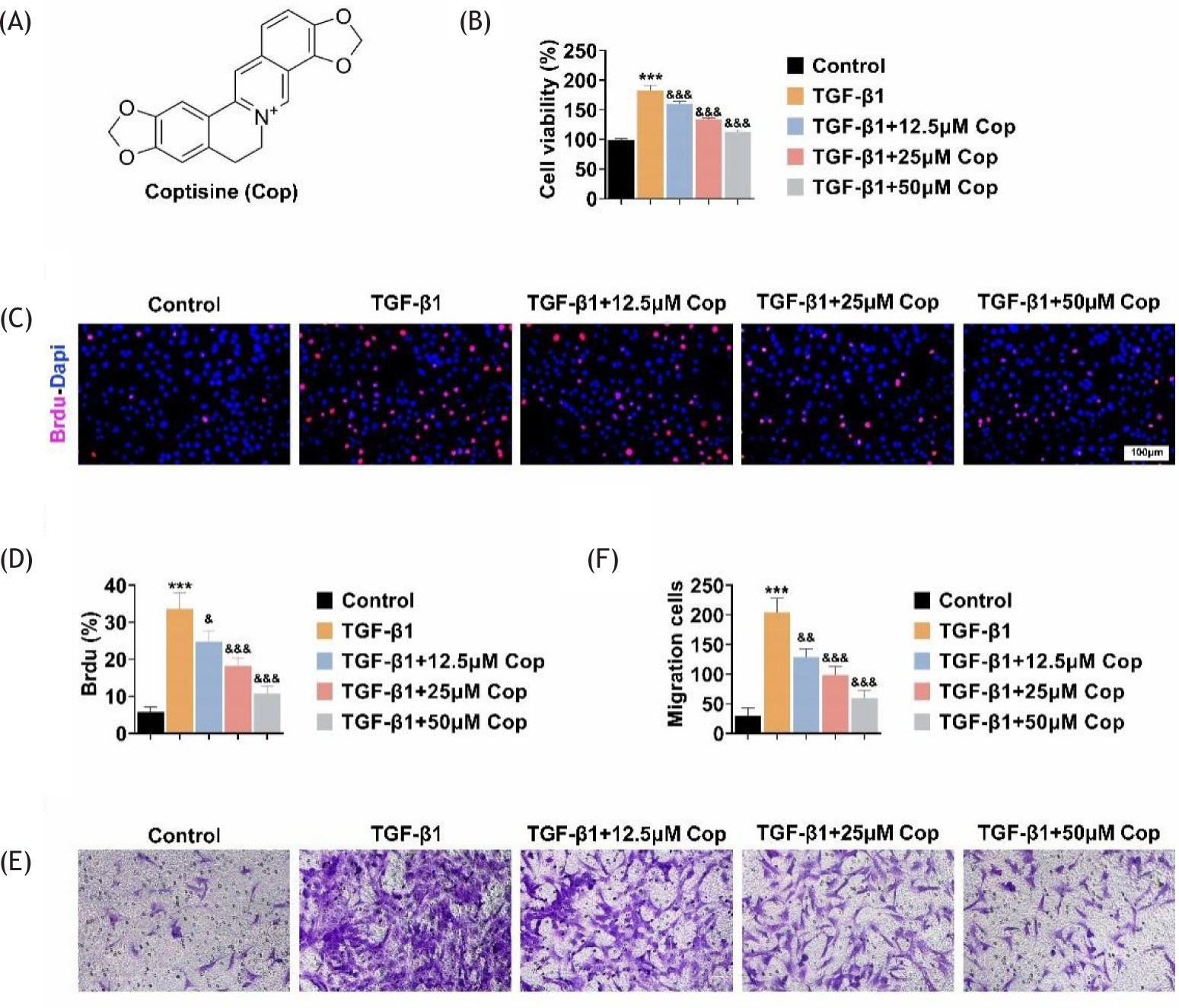
In order to detect the effects of Rhizoma coptidis on the progression of PF in vitro, we first established a cell model by treating HFL1 cells with TGF-β1. The molecular formula of Rhizoma coptidis is shown in Figure 1A. Through CCK-8 assay, the results showed that TGF-β1 dramatically increased the growth of HFL1 cells, with the increased OD450 value (Figure 1B). Rhizoma coptidis treatment decreased the OD450 value of TGF-β1-treated HFL1 cells, suggesting the suppression of cell growth (Figure 1B). Similarly, BrdU assay also confirmed that Rhizoma coptidis treatment reversed the promotion of cell growth caused by TGF-β1 treatment in HFL1 cells, with the decreased percentage of BrdU-positive cells (Figures 1C,D).
Figure 1 Rhizoma coptidis inhibits excessive growth as well as motility of TGF-β1-induced HFL1 cells. (A) Molecular formula of Rhizoma coptidis (COP. (B) CCK-8 assay indicates the growth of HFL1 cells upon the treatment of TGF-β1 with COP at concentrations of 0, 12.5, 25, and 50 μM for 24 h. The OD450 value was measured. (C) BrdU assay indicates the growth of HFL1 cells upon the treatment of TGF-β1 with COP at concentrations of 0, 12.5, 25, and 50 μM for 24 h. Red panel indicates BrdU. Scale bar: 100 μm. (D) Quantification of panel C. The percentage of BrdU positive cells was calculated. (E) Transwell assay indicates the effects of COP on the migration of HFL1 cells upon the treatment of TGF-β1 at concentrations of 0, 12.5, 25, and 50 μM for 24 h. (F) Quantification of panel E. The number of migration cells was quantified. ***P < 0.001, TGF-β1 vs control, &P < 0.05, &&P < 0.01, &&&P < 0.001, TGF-β1+COP vs TGF-β1.
Further, the transwell assay was performed to confirm the effects of Rhizoma coptidis on the migration of HFL1 cells. The results indicated that TGF-β1 treatment contributed to the migration of HFL1 cells, whereas Rhizoma coptidis treatment suppressed the migration of TGF-β1-treated HFL1 cells (Figures 1D,E). Collectively, Rhizoma coptidis inhibits excessive growth and motility of TGF-β1-induced HFL1 cells.
Rhizoma coptidis inhibits inflammation and ROS levels in TGF-β1-induced HFL1 cells
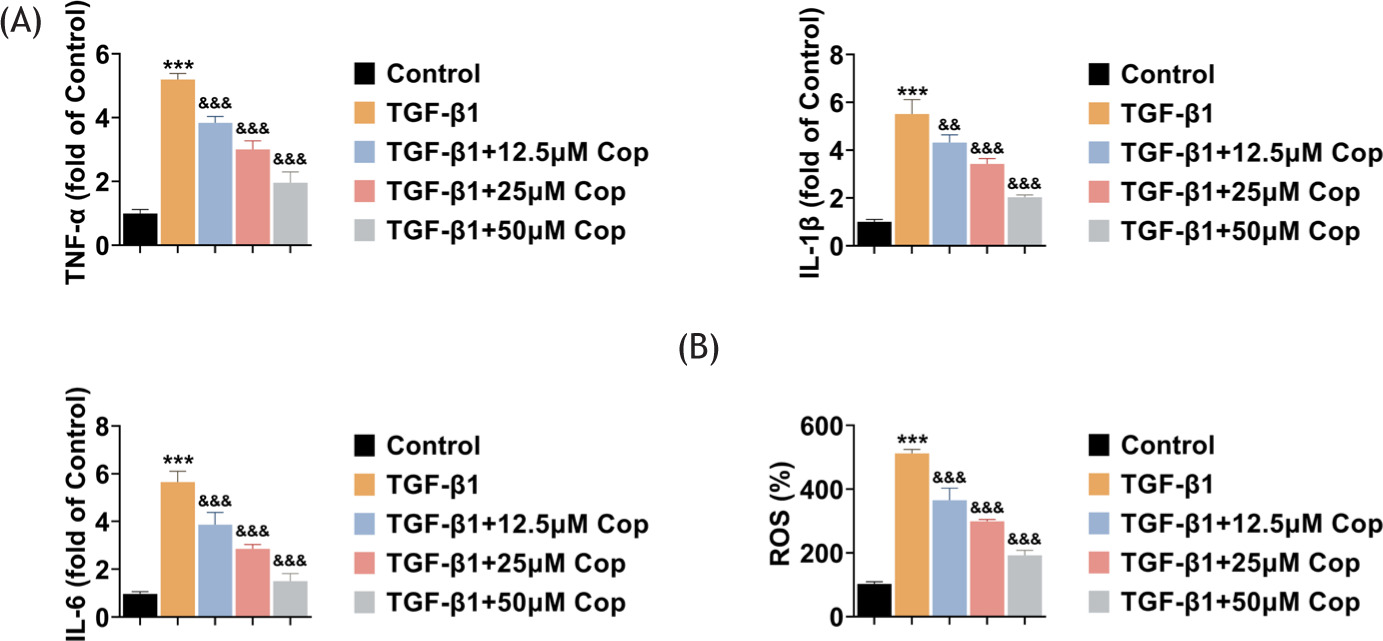
Subsequently, the effects of Rhizoma coptidis on the inflammation and ROS production were observed in HFL1 cells. ELISA results showed that the secretion of several inflammatory factors, such as TNF-α, IL-1β, and IL-6, were found in HFL1 cells and TGF-β1 treatment increased the secretion of these factors, suggesting the promotion of inflammation (Figure 2A). However, the treatment of Rhizoma coptidis suppressed the secretion of these factors in TGF-β1-induced HFL1 cells (Figure 2A). Further, through the detection of ROS levels, the results confirmed that TGF-β1 contributed to the production of ROS in HFL1 cells, whereas Rhizoma coptidis treatment suppressed ROS levels in TGF-β1-induced HFL1 cells (Figure 2B). Therefore, Rhizoma coptidis inhibits inflammation and ROS levels in TGF-β1-induced HFL1 cells.
Figure 2 Rhizoma coptidis (COP) inhibits inflammation and ROS levels in TGF-β1-induced HFL1 cells. (A) ELISA indicates the secretion of TNF-α, IL-1β, and IL-6 of HFL1 cells upon the treatment of TGF-β1 with COP at concentrations of 0, 12.5, 25, and 50 μM for 24 h. (B) ROS levels in HFL1 cells upon treatment of TGF-β1 with COP at concentrations of 0, 12.5, 25, and 50 μM for 24 h were quantified. ***P < 0.001, TGF-β1 vs control, &&P < 0.01, &&&P < 0.001, TGF-β1+COP vs TGF-β1.
Rhizoma coptidis inhibits the FMT process of TGF-β1-induced HFL1 cells
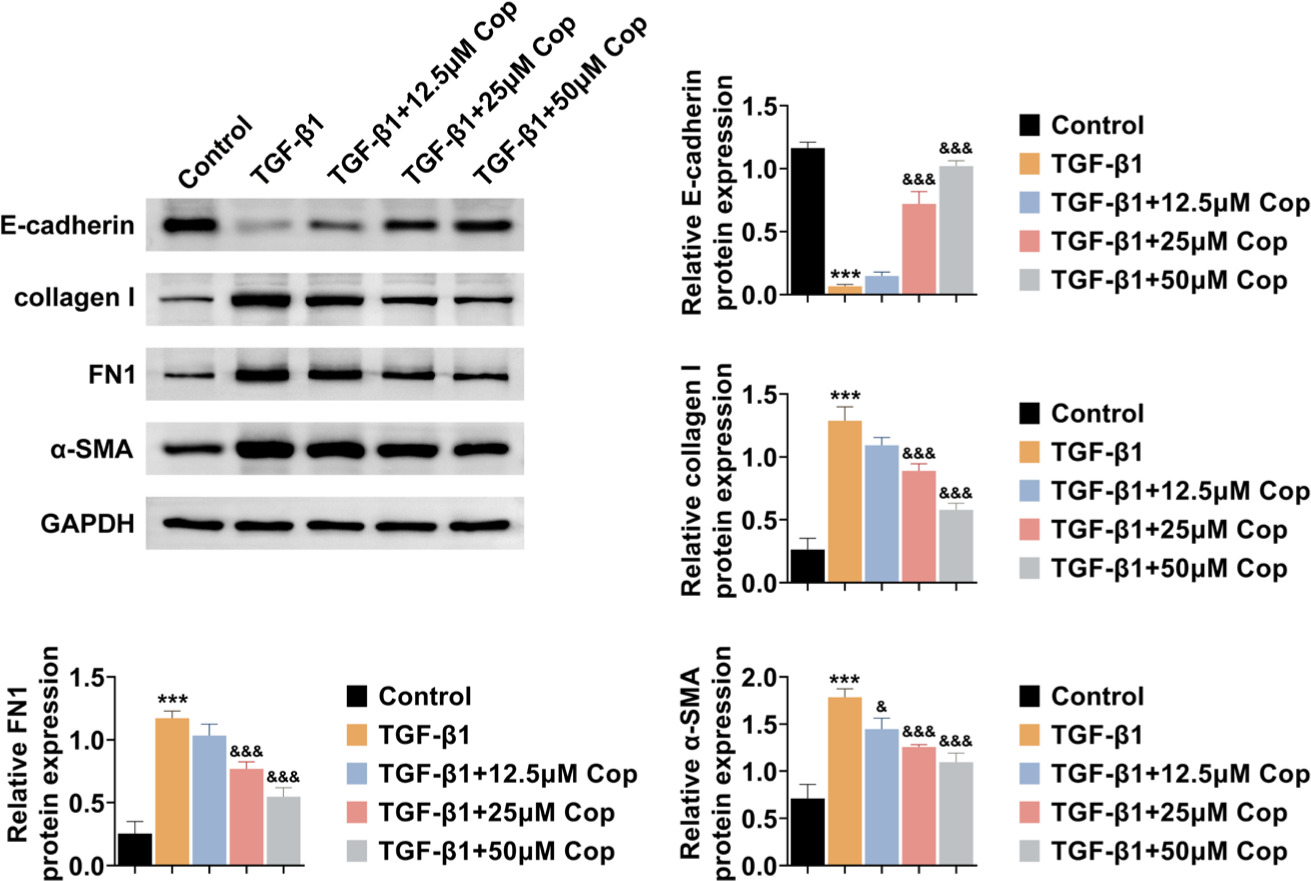
Then the effects of Rhizoma coptidis on the process of FMT were observed in HFL1 cells. Immunoblot assay showed the expression of several markers of FMT, such as E-cadherin, collagen I, FN1, and α-SMA, in HFL1 cells. The results showed that TGF-β1 suppressed the expression of E-cadherin but increased the expression of collagen I, FN1, and α-SMA in HFL1 cells (Figure 3). However, Rhizoma coptidis further increased the expression of E-cadherin and decreased the expression of collagen I, FN1, and α-SMA in TGF-β1-induced HFL1 cells, suggesting the suppression of FMT (Figure 3). Finally, Rhizoma coptidis inhibits the FMT process of TGF-β1-induced HFL1 cells.
Figure 3 Rhizoma coptidis (COP) inhibits the FMT process of TGF-β1-induced HFL1 cells. Immunoblot assay shows the expression of E-cadherin, collagen I, FN1, and α-SMA of HFL1 cells upon the treatment of TGF-β1 with COP at the concentration of 0, 12.5, 25, and 50 μM for 24 h. The relative expressions of the indicated proteins were quantified. ***P < 0.001, TGF-β1 vs control, &P < 0.05, &&&P < 0.001, TGF-β1+COP vs TGF-β1.
Rhizoma coptidis promotes Nrf2/HO-1 pathway in TGF-β1-induced HFL1 cells
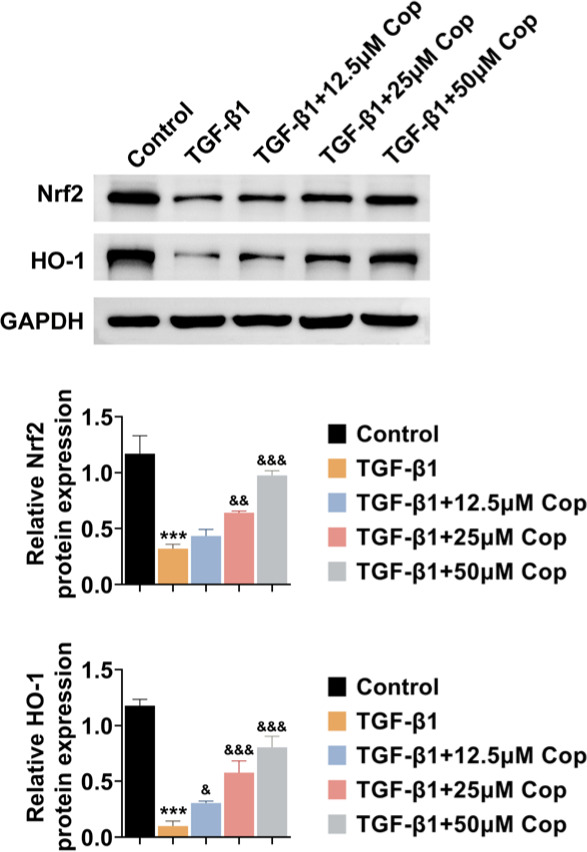
Finally, the possible mechanism by which Rhizoma coptidis affects the inflammation and FMT of HFL1 cells was further investigated. The effects of Rhizoma coptidis on the Nrf2/HO-1 pathway were observed through immunoblot assay. Interestingly, TGF-β1 treatment decreased the expression of Nrf2 and HO-1 in HFL1 cells, suggesting the suppression of the Nrf2/HO-1 pathway (Figure 4). Interestingly, Rhizoma coptidis treatment increased the expression of Nrf2 and HO-1 in TGF-β1-induced HFL1 cells, suggesting promotion of the Nrf2/HO-1 pathway (Figure 4). Therefore, Rhizoma coptidis promotes the Nrf2/HO-1 pathway in TGF-β1-induced HFL1 cells.
Figure 4 Rhizoma coptidis (COP) promotes the Nrf2/HO-1 pathway in TGF-β1-induced HFL1 cells. Immunoblot assay shows the expression of Nrf2 and HO-1 of HFL1 cells upon the treatment of TGF-β1 with COP at concentrations of 0, 12.5, 25, and 50 μM for 24 h. The relative expressions of the indicated proteins were quantified. ***P < 0.001, TGF-β1 vs control, &P < 0.05, &&P < 0.01, &&&P < 0.001, TGF-β1+COP vs TGF-β1.
Discussion
Pulmonary fibrosis is a chronic and progressive lung disease characterized by excessive scarring and hardening of lung tissues, leading to a loss of lung elasticity, thus severely affecting respiratory function and gas exchange.9 It could be caused by different factors, such as long-term exposure to harmful substances, side effects of certain medications, chronic inflammatory diseases, genetic factors, or, in many cases, by unknown factors. The development of PF involves chronic inflammatory responses, activation of fibroblasts and their transformation into myofibroblasts, and accumulation of collagen and other extracellular matrix proteins, ultimately leading to remodeling and functional impairment of lung tissues.10 Treatment primarily focuses on slowing of disease progression, lessening of clinical manifestations, and enhancing quality of life, such as using anti-fibrotic and anti-inflammatory medications, oxygen therapy, and considering lung transplantation in severe cases. Therapy for PF mainly includes anti-fibrotic drugs (such as nitnanib and perfenidone) to slow the decline of lung function, corticosteroids, immunosuppressants (such as cyclosporine, azathioprine, and metopodone) to control inflammation, and antacids to treat gastroesophageal reflux.10,11 Herein, the results confirmed that Rhizoma coptidis inhibits hyperproliferation, inflammation as well as myofibroblast transformation of lung fibroblasts. Therefore, it has the potential to serve as a therapy to combat PF.
Transition from fibroblasts to myofibroblasts is a key mechanism in the pathogenesis of PF. In this process, normal fibroblasts transform into myofibroblasts under the stimulation of growth factors (such as TGF-β) and inflammatory cytokines. Unlike fibroblasts, which primarily synthesize collagen and other extracellular matrix components, myofibroblasts also have contractile properties that lead to tissue hardening and contraction.12 These myofibroblasts play a crucial role in PF by producing an abundance of collagen, leading to scarring and hardening of lung tissues, thereby exacerbating tissue stiffness and functional impairment.13 Understanding the mechanism of this transition is vital for developing new therapeutic strategies for PF, such as inhibiting this transformation or reversing the transition that has already occurred. Interestingly, we constructed a cell model of PF by using TGF-β1 and confirmed that Rhizoma coptidis inhibits the FMT process of TGF-β1-induced HFL1 cells, thereby suppressing PF.
Rhizoma coptidis, an alkaloid found in Coptis chinensis, the Chinese goldthread, is known for its multiple pharmacological activities. It exhibits significant antibacterial properties by disrupting bacterial cell walls and membranes, thus inhibiting bacterial growth.14 In addition, Rhizoma coptidis has anti-inflammatory effects, potentially achieved by inhibiting the production of inflammatory mediators and regulating the activity of immune cells. Concerning antitumor activity, it shows antiproliferative effects on certain types of cancer cells, possibly by inducing apoptosis and interfering with tumor angiogenesis.15 Rhizoma coptidis also has antioxidant properties, capable of scavenging free radicals and protecting cells from oxidative stress damage. Our study confirmed its activities on mediating ROS, inflammation, and FMT of HFL1 cells. Despite these potential pharmacological effects, further studies are required to clarify its mechanism of action and safety and efficacy in humans.
The Nrf2/HO-1 pathway plays a crucial role in PF. This pathway is a key regulator of cellular antioxidant responses, essential for maintaining cellular redox balance and protecting against oxidative stress.16,17 Nrf2, a transcription factor, is activated in response to oxidative stress, transferring to the nucleus to promote the expression of antioxidant and cell-protective genes, such as HO-1. HO-1, a downstream target of Nrf2, degrades heme to release iron, carbon monoxide, and biliverdin, which have anti-inflammatory and cytoprotective effects, aiding in mitigation of tissue damage. In the context of PF, oxidative stress is a key factor driving disease progression, and the activation of Nrf2 and expression of HO-1 can help alleviate this stress, potentially slowing the progression of PF.18 Therefore, activation of the Nrf2/HO-1 pathway is considered as a potential strategy for treating PF, enhancing lung tissues’ antioxidant capacity to reduce inflammation and fibrosis, thus offering a new avenue to treatment. Our study confirmed that Rhizoma coptidis suppressed the progression of PF through mediating this pathway. However, the precise mechanism needs further study.
There are certain limitations to this study. First, animal experiments need to be conducted in the future to further confirm the results in vitro. Next, an animal model must be constructed to better validate the therapeutic effects of Rhizoma coptidis. In addition, this study lacks in-depth mechanism research. We discovered that although Rhizoma coptidis targets the Nrf2/HO-1 pathway, its precise mechanism is still unclear. In future studies, biochemical experiments must be conducted to verify downstream regulatory factors.
Conclusion
Rhizoma coptidis inhibits inflammation as well as myofibroblast transformation of lung fibroblasts through the Nrf2/HO-1 pathway. It could therefore serve as a potential therapy for PF.
Funding
This work was supported by Natural Science Research Project of Anhui Educational Committee (Grant No. 2022AH050791), Research Fund Project of Anhui Medical University (Grant No. 2022xkj206), and Health Research Program of Anhui (Grant No. AHWJ2023BAa20196).
Competing interests
The authors stated that there was no conflict of interest to declare.
Data availability
The authors declare that all data supporting the findings of this study are available within the paper and any raw data can be obtained from the corresponding author upon request.
Author Contributions
Jie Yang and Yuting Huang designed and conducted the study. Jie Yang, Yuting Huang, Zhimin Cui, Chang Liu, and Guofang Xie supervised data collection, analysis, and interpretation. Jie Yang and Yuting Huang prepared the manuscript for publication and reviewed its draft. All authors read and approved the final manuscript for publication.
REFERENCES
1. Thannickal VJ, Toews GB, White ES, Lynch JP, 3rd, Martinez FJ. Mechanisms of pulmonary fibrosis. Annu Rev Med. 2004;55:395–417. 10.1146/annurev.med.55.091902.103810
2. Koudstaal T, Funke-Chambour M, Kreuter M, Molyneaux PL, Wijsenbeek MS. Pulmonary fibrosis: From pathogenesis to clinical decision-making. Trends Mol Med. 2023;29(12):1076–87. 10.1016/j.molmed.2023.08.010
3. Yi XM, Li M, Chen YD, Shu HB, Li S. Reciprocal regulation of IL-33 receptor-mediated inflammatory response and pulmonary fibrosis by TRAF6 and USP38. Proc Natl Acad Sci USA. 2022;119(10):e2116279119. 10.1073/pnas.2116279119
4. Wu J, Luo Y, Jiang Q, Li S, Huang W, Xiang L, et al. Rhizoma coptidis from Coptis chinensis blocks NLRP3 inflammasome activation by inhibiting caspase-1. Pharmacol Res. 2019;147:104348. 10.1016/j.phrs.2019.104348
5. Wu J, Luo Y, Deng D, Su S, Li S, Xiang L, et al. Rhizoma coptidis from Coptis chinensis exerts diverse beneficial properties: A concise review. J Cell Mol Med. 2019;23(12):7946–60. 10.1111/jcmm.14725
6. Huang J, Ren K, Huang L. Rhizoma coptidis attenuates sepsis lung injury by suppressing LPS-induced lung epithelial cell inflammation and apoptosis. Allergol Immunopathol (Madr). 2023;51(6):30–6. 10.15586/v51i6.972
7. Jo HG, Park C, Lee H, Kim GY, Keum YS, Hyun JW, et al. Inhibition of oxidative stress induced-cytotoxicity by Rhizoma coptidis in V79-4 Chinese hamster lung fibroblasts through the induction of Nrf-2-mediated HO-1 expression. Genes Genomics. 2021;43(1):17–31. 10.1007/s13258-020-01018-3
8. Rao PC, Begum S, Sahai M, Sriram DS. Rhizoma coptidis--induced cell cycle arrest at G2/M phase and reactive oxygen species-dependent mitochondria-mediated apoptosis in non-small-cell lung cancer A549 cells. Tumour Biol. 2017;39(3):1010428317694565. 10.1177/1010428317694565
9. Savin IA, Zenkova MA, Sen’kova AV. Pulmonary fibrosis as a result of acute lung inflammation: Molecular mechanisms, relevant in vivo models, prognostic and therapeutic approaches. Int J Mol Sci. 2022;23(23):14959. 10.3390/ijms232314959
10. Kinoshita T, Goto T. Molecular mechanisms of pulmonary fibrogenesis and its progression to lung cancer: A review. Int J Mol Sci. 2019;20(6):1461. 10.3390/ijms20061461
11. Xiaoshen Zhang LC. MiR-216b modulates cisplatin resistance and stem cell-like features in breast cancer cells by targeting E2F4. Trop J Pharm Res. 2023;22(1). 10.4314/tjpr.v22i1.1
12. Saha P, Talwar P. Idiopathic pulmonary fibrosis (IPF): Disease pathophysiology, targets, and potential therapeutic interventions. Mol Cell Biochem. 2023. 10.1007/s11010-23-04845-6
13. Wynn TA. Integrating mechanisms of pulmonary fibrosis. J Exp Med. 2011;208(7):1339–50. 10.1084/jem.20110551
14. Zhang R, Tian S, Zhang T, Zhang W, Lu Q, Hu Q, et al. Antibacterial activity mechanism of Rhizoma coptidis against Pasteurella multocida. Front Cell Infect Microbiol. 2023;13:1207855. 10.3389/fcimb.2023.1207855
15. Zhou L, Yang F, Li G, Huang J, Liu Y, Zhang Q, et al. Rhizoma coptidis induces apoptosis in human hepatoma cells through activating 67-kDa laminin receptor/cGMP signaling. Front Pharmacol. 2018;9:517. 10.3389/fphar.2018.00517
16. Zhang Q, Liu J, Duan H, Li R, Peng W, Wu C. Activation of Nrf2/HO-1 signaling: An important molecular mechanism of herbal medicine in the treatment of atherosclerosis via the protection of vascular endothelial cells from oxidative stress. J Adv Res. 2021;34:43–63. 10.1016/j.jare.2021.06.023
17. Ren K, Pei J, Guo Y, Jiao Y, Xing H, Xie Y, et al. Regulated necrosis pathways: A potential target for ischemic stroke. Burns Trauma. 2023;11:tkad016. 10.1093/burnst/tkad016
18. Wang Y, Wei J, Deng H, Zheng L, Yang H, Lv X. The role of Nrf2 in pulmonary fibrosis: Molecular mechanisms and treatment approaches. Antioxidants (Basel). 2022;11(9):1685. 10.3390/antiox11091685